
Exoproteome Analysis of Antagonistic Interactions between the Probiotic Bacteria Limosilactobacillus reuteri LR1 and Lacticaseibacillus rhamnosus F and Multidrug Resistant Strain of Klebsiella pneumonia

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Sequencing and Annotation of L. reuteri LR1 and L. rhamnosus F Genomes
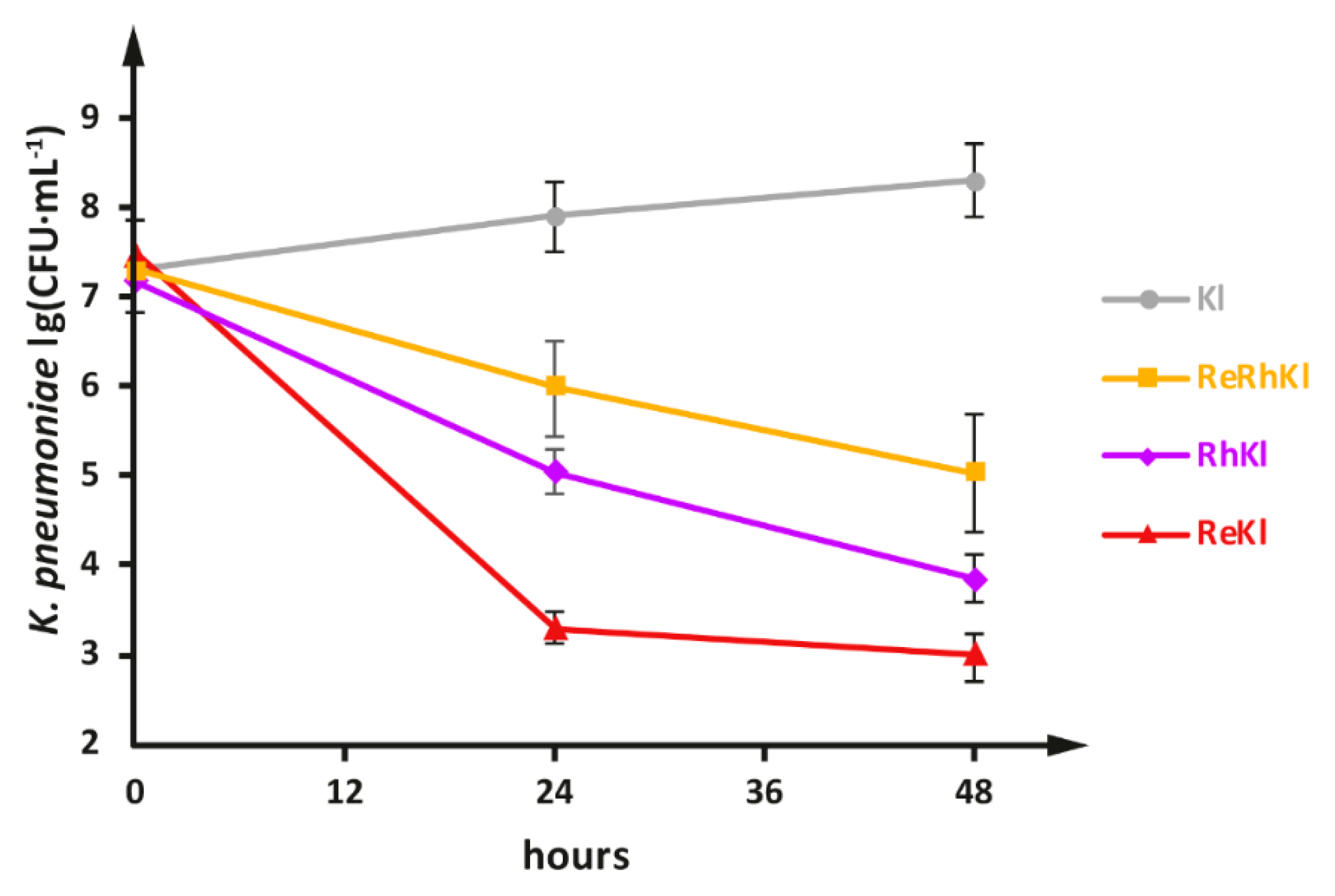
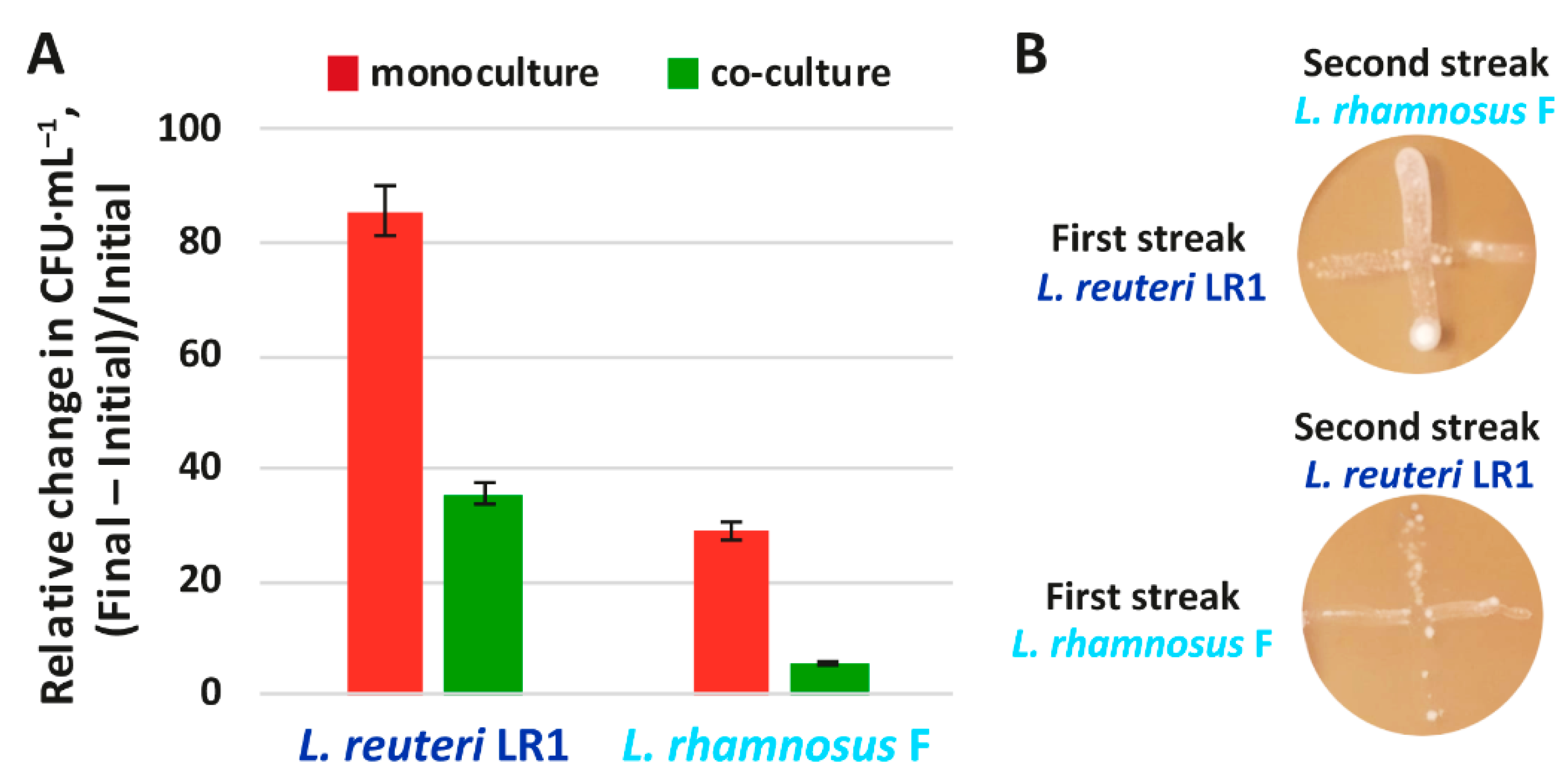
2.2. Co-Cultivation of L. reuteriLR1 and L. rhamnosus F with Multidrug Resistant K. pneumonia
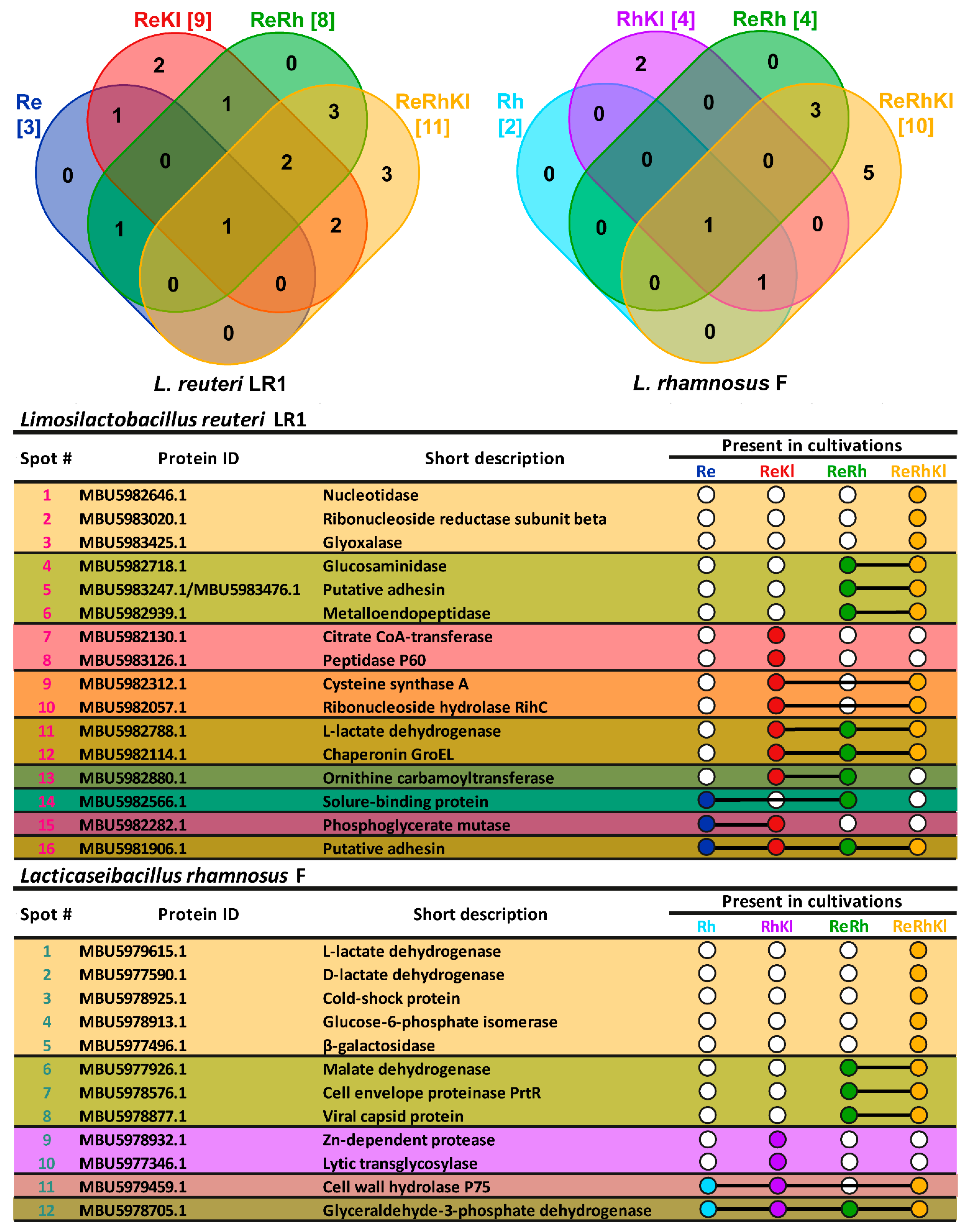
2.3. Compositional Analysis of Exoproteomes
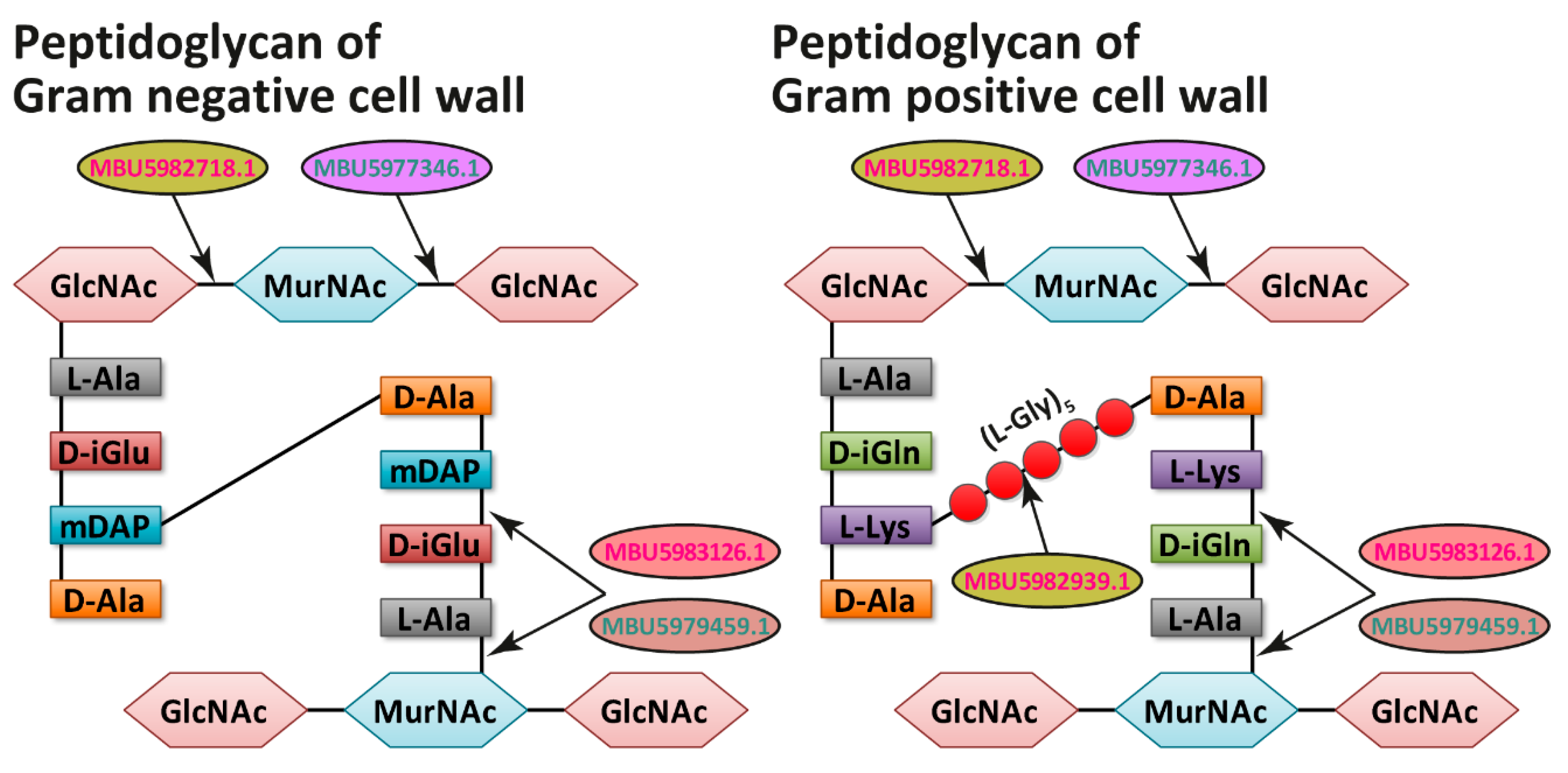
2.4. Functional Analysis of Exoproteomes
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Pre-Cultivations
4.2. DNA Isolation, Sequencing and Annotation
4.3. Co-Cultivations of LAB with K. pneumoniae
4.4. Co-Cultivations of LAB with Each Other
4.5. Perpendicular Streak Test
4.6. Exoproteomics Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tanwar, J.; Das, S.; Fatima, Z.; Hameed, S. Multidrug Resistance: An Emerging Crisis. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neubeiser, A.; Bonsignore, M.; Tafelski, S.; Alefelder, C.; Schwegmann, K.; Rüden, H.; Geffers, C.; Nachtigall, I. Mortality attributable to hospital acquired infections with multidrug-resistant bacteria in a large group of German hospitals. J. Infect. Public Health 2020, 13, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Burnham, J.P.; Olsen, M.A.; Kollef, M.H. Re-estimating annual deaths due to multidrug-resistant organism infections. Infect. Control Hosp. Epidemiol. 2019, 40, 112–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, R.E.; Hatfield, K.M.; Wolford, H.; Samore, M.H.; Scott, R.D.; Reddy, S.C.; Olubajo, B.; Paul, P.; Jernigan, J.A.; Baggs, J. National Estimates of Healthcare Costs Associated With Multidrug-Resistant Bacterial Infections Among Hospitalized Patients in the United States. Clin. Infect. Dis. 2021, 72, S17–S26. [Google Scholar] [CrossRef]
- Silva, D.R.; de Sardi, J.C.O.; de Pitangui, N.S.; Roque, S.M.; da Silva, A.C.B.; Rosalen, P.L. Probiotics as an alternative antimicrobial therapy: Current reality and future directions. J. Funct. Foods 2020, 73, 104080. [Google Scholar] [CrossRef]
- Yang, H.; Sun, Y.; Cai, R.; Chen, Y.; Gu, B. The impact of dietary fiber and probiotics in infectious diseases. Microb. Pathog. 2020, 140, 103931. [Google Scholar] [CrossRef]
- Vuotto, C.; Longo, F.; Donelli, G. Probiotics to counteract biofilm-associated infections: Promising and conflicting data. Int. J. Oral Sci. 2014, 6, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakhmanova, A.; Khan, Z.A.; Shah, K. A mini review fermentation and preservation: Role of Lactic Acid Bacteria. MOJ Food Process. Technol. 2018, 6, 414–417. [Google Scholar] [CrossRef]
- Dimidi, E.; Cox, S.; Rossi, M.; Whelan, K. Fermented Foods: Definitions and Characteristics, Impact on the Gut Microbiota and Effects on Gastrointestinal Health and Disease. Nutrients 2019, 11, 1806. [Google Scholar] [CrossRef] [Green Version]
- Rozhkova, I.V.; Moiseenko, K.V.; Glazunova, O.A.; Begunova, A.V.; Fedorova, T.V. 9 Russia and Commonwealth of Independent States (CIS) domestic fermented milk products. In Food Science and Technology; De Gruyter: Berlin, Germany, 2020; pp. 215–234. [Google Scholar]
- De Vuyst, L.; Vandamme, E.J. Antimicrobial Potential of Lactic Acid Bacteria. In Bacteriocins of Lactic Acid Bacteria; Springer: Boston, MA, USA, 1994; pp. 91–142. [Google Scholar]
- Barzegari, A.; Kheyrolahzadeh, K.; Hosseiniyan Khatibi, S.M.; Sharifi, S.; Memar, M.Y.; Zununi Vahed, S. The Battle of Probiotics and Their Derivatives Against Biofilms. Infect. Drug Resist. 2020, 13, 659–672. [Google Scholar] [CrossRef] [Green Version]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and Inputs From Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters During Food-Animal Production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, J.A.; Paula, A.T.; Casarotti, S.N.; Penna, A.L.B. Lactic Acid Bacteria Antimicrobial Compounds: Characteristics and Applications. Food Eng. Rev. 2012, 4, 124–140. [Google Scholar] [CrossRef]
- Bengoechea, J.A.; Sa Pessoa, J. Klebsiella pneumoniae infection biology: Living to counteract host defences. FEMS Microbiol. Rev. 2019, 43, 123–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyres, K.L.; Holt, K.E. Klebsiella pneumoniae as a key trafficker of drug resistance genes from environmental to clinically important bacteria. Curr. Opin. Microbiol. 2018, 45, 131–139. [Google Scholar] [CrossRef]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef]
- González, J.F.; Hahn, M.M.; Gunn, J.S. Chronic biofilm-based infections: Skewing of the immune response. Pathog. Dis. 2018, 76, fty023. [Google Scholar] [CrossRef]
- El-Mokhtar, M.A.; Hassanein, K.M.; Ahmed, A.S.; Gad, G.F.; Amin, M.M.; Hassanein, O.F. Antagonistic Activities of Cell-Free Supernatants of Lactobacilli Against Extended-Spectrum β-Lactamase Producing Klebsiella pneumoniae and Pseudomonas aeruginosa. Infect. Drug Resist. 2020, 13, 543–552. [Google Scholar] [CrossRef] [Green Version]
- Mogna, L.; Deidda, F.; Nicola, S.; Amoruso, A.; Del Piano, M.; Mogna, G. In Vitro Inhibition of Klebsiella pneumoniae by Lactobacillus delbrueckii Subsp. delbrueckii LDD01 (DSM 22106). J. Clin. Gastroenterol. 2016, 50, S136–S139. [Google Scholar] [CrossRef]
- Raras, T.; Firman, A.; Kinanti, I.; Noorhamdani, N. Anti-Biofilm Activity of Lactic Acid Bacteria Isolated from Kefir Against Multidrug-Resistant Klebsiella pneumoniae. J. Pure Appl. Microbiol. 2019, 13, 983–992. [Google Scholar] [CrossRef] [Green Version]
- Lagrafeuille, R.; Miquel, S.; Balestrino, D.; Vareille-Delarbre, M.; Chain, F.; Langella, P.; Forestier, C. Opposing effect of Lactobacillus on in vitro Klebsiella pneumoniae in biofilm and in an in vivo intestinal colonisation model. Benef. Microbes 2018, 9, 87–100. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. eggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Bendtsen, J.D.; Kiemer, L.; Fausbøll, A.; Brunak, S. Non-classical protein secretion in bacteria. BMC Microbiol. 2005, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- McFarland, L.V. Efficacy of Single-Strain Probiotics Versus Multi-Strain Mixtures: Systematic Review of Strain and Disease Specificity. Dig. Dis. Sci. 2021, 66, 694–704. [Google Scholar] [CrossRef]
- Kitts, P.A.; Church, D.M.; Thibaud-Nissen, F.; Choi, J.; Hem, V.; Sapojnikov, V.; Smith, R.G.; Tatusova, T.; Xiang, C.; Zherikov, A.; et al. Assembly: A resource for assembled genomes at NCBI. Nucleic Acids Res. 2016, 44, D73–D80. [Google Scholar] [CrossRef]
- Mitrea, L.; Vodnar, D.C. Klebsiella pneumoniae—A Useful Pathogenic Strain for Biotechnological Purposes: Diols Biosynthesis under Controlled and Uncontrolled pH Levels. Pathogens 2019, 8, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Tang, H.; Lin, Z.; Xu, P. Mechanisms of acid tolerance in bacteria and prospects in biotechnology and bioremediation. Biotechnol. Adv. 2015, 33, 1484–1492. [Google Scholar] [CrossRef]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Said, L.B.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a new generation of antimicrobials: Toxicity aspects and regulations. FEMS Microbiol. Rev. 2021, 45, fuaa039. [Google Scholar] [CrossRef] [PubMed]
- Prudêncio, C.V.; dos Santos, M.T.; Vanetti, M.C.D. Strategies for the use of bacteriocins in Gram-negative bacteria: Relevance in food microbiology. J. Food Sci. Technol. 2015, 52, 5408–5417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Åvall-Jääskeläinen, S.; Palva, A. Lactobacillus surface layers and their applications. FEMS Microbiol. Rev. 2005, 29, 511–529. [Google Scholar] [CrossRef] [Green Version]
- Teame, T.; Wang, A.; Xie, M.; Zhang, Z.; Yang, Y.; Ding, Q.; Gao, C.; Olsen, R.E.; Ran, C.; Zhou, Z. Paraprobiotics and Postbiotics of Probiotic Lactobacilli, Their Positive Effects on the Host and Action Mechanisms: A Review. Front. Nutr. 2020, 7, 570344. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Urdaci, M.C.; Margolles, A. Extracellular proteins secreted by probiotic bacteria as mediators of effects that promote mucosa–bacteria interactions. Microbiology 2010, 156, 3232–3242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Q.; Zhang, D. Principle and potential applications of the non-classical protein secretory pathway in bacteria. Appl. Microbiol. Biotechnol. 2020, 104, 953–965. [Google Scholar] [CrossRef] [PubMed]
- Vermassen, A.; Leroy, S.; Talon, R.; Provot, C.; Popowska, M.; Desvaux, M. Cell Wall Hydrolases in Bacteria: Insight on the Diversity of Cell Wall Amidases, Glycosidases and Peptidases Toward Peptidoglycan. Front. Microbiol. 2019, 10, 331. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.-S.; Lim, H.-S.; Oh, J.-S.; Lim, Y.; Wuertz-Kozak, K.; Harro, J.M.; Shirtliff, M.E.; Achermann, Y. Antimicrobial activity of Lactobacillus salivarius and Lactobacillus fermentum against Staphylococcus aureus. Pathog. Dis. 2017, 75, ftx009. [Google Scholar] [CrossRef]
- Xu, Q.; Sudek, S.; McMullan, D.; Miller, M.D.; Geierstanger, B.; Jones, D.H.; Krishna, S.S.; Spraggon, G.; Bursalay, B.; Abdubek, P.; et al. Structural Basis of Murein Peptide Specificity of a γ-D-Glutamyl-L-Diamino Acid Endopeptidase. Structure 2009, 17, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.J.; Jun, J.S.; Moon, J.A.; Hong, K.W. Surface display of p75, a Lactobacillus rhamnosus GG derived protein, on Bacillus subtilis spores and its antibacterial activity against Listeria monocytogenes. AMB Express 2020, 10, 139. [Google Scholar] [CrossRef]
- Singh, N.; Bhalla, N. Moonlighting Proteins. Annu. Rev. Genet. 2020, 54, 265–285. [Google Scholar] [CrossRef]
- Copley, S.D. Moonlighting is mainstream: Paradigm adjustment required. BioEssays 2012, 34, 578–588. [Google Scholar] [CrossRef]
- Jeffery, C.J. Moonlighting proteins. Trends Biochem. Sci. 1999, 24, 8–11. [Google Scholar] [CrossRef]
- Jeffery, C.J. Protein moonlighting: What is it, and why is it important? Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20160523. [Google Scholar] [CrossRef] [PubMed]
- Kainulainen, V.; Korhonen, T. Dancing to Another Tune—Adhesive Moonlighting Proteins in Bacteria. Biology 2014, 3, 178–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffery, C. Intracellular proteins moonlighting as bacterial adhesion factors. AIMS Microbiol. 2018, 4, 362–376. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, J.C. Intracellular/surface moonlighting proteins that aid in the attachment of gut microbiota to the host. AIMS Microbiol. 2019, 5, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.; Gupta, A.; Gupta, V. Insights into multifaceted activities of CysK for therapeutic interventions. 3 Biotech 2019, 9, 44. [Google Scholar] [CrossRef]
- Pal, V.K.; Bandyopadhyay, P.; Singh, A. Hydrogen sulfide in physiology and pathogenesis of bacteria and viruses. IUBMB Life 2018, 70, 393–410. [Google Scholar] [CrossRef]
- Bernier, S.P.; Létoffé, S.; Delepierre, M.; Ghigo, J.-M. Biogenic ammonia modifies antibiotic resistance at a distance in physically separated bacteria. Mol. Microbiol. 2011, 81, 705–716. [Google Scholar] [CrossRef]
- Juarez, G.E.; Mateyca, C.; Galvan, E.M. Proteus mirabilis outcompetes Klebsiella pneumoniae in artificial urine medium through secretion of ammonia and other volatile compounds. Heliyon 2020, 6, e03361. [Google Scholar] [CrossRef]
- Wang, X.; Zheng, Z.; Dou, P.; Qin, J.; Wang, X.; Ma, C.; Tang, H.; Xu, P. Cloning, expression, purification, and activity assay of proteins related to D-lactic acid formation in Lactobacillus rhamnosus. Appl. Microbiol. Biotechnol. 2010, 87, 2117–2123. [Google Scholar] [CrossRef]
- Viana, R.; Yebra, M.J.; Galán, J.L.; Monedero, V.; Pérez-Martínez, G. Pleiotropic effects of lactate dehydrogenase inactivation in Lactobacillus casei. Res. Microbiol. 2005, 156, 641–649. [Google Scholar] [CrossRef]
- Valerio, F. Production of phenyllactic acid by lactic acid bacteria: An approach to the selection of strains contributing to food quality and preservation. FEMS Microbiol. Lett. 2004, 233, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Hummel, W.; Schütte, H.; Kula, M.R. d-2-hydroxyisocaproate dehydrogenase from Lactobacillus casei-A new enzyme suitable for stereospecific reduction of 2-ketocarboxylic acids. Appl. Microbiol. Biotechnol. 1985, 21, 7–15. [Google Scholar] [CrossRef]
- Liu, F.; Tang, C.; Wang, D.; Sun, Z.; Du, L.; Wang, D. The synergistic effects of phenyllactic acid and slightly acid electrolyzed water to effectively inactivate Klebsiella oxytoca planktonic and biofilm cells. Food Control 2021, 125, 107804. [Google Scholar] [CrossRef]
- Cumby, N.; Davidson, A.R.; Maxwell, K.L. The moron comes of age. Bacteriophage 2012, 2, e23146. [Google Scholar] [CrossRef] [PubMed]
- Owen, S.V.; Canals, R.; Wenner, N.; Hammarlöf, D.L.; Kröger, C.; Hinton, J.C.D. A window into lysogeny: Revealing temperate phage biology with transcriptomics. Microb. Genom. 2020, 6, e000330. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Keightley, A.; Biswas, I. Characterization of a stress tolerance-defective mutant of Lactobacillus rhamnosus LRB. Mol. Oral Microbiol. 2019, 34, 153–167. [Google Scholar] [CrossRef]
- Jarocki, P.; Komoń-Janczara, E.; Podleśny, M.; Kholiavskyi, O.; Pytka, M.; Kordowska-Wiater, M. Genomic and Proteomic Characterization of Bacteriophage BH1 Spontaneously Released from Probiotic Lactobacillus rhamnosus Pen. Viruses 2019, 11, 1163. [Google Scholar] [CrossRef] [Green Version]
- Fedorova, T.V.; Vasina, D.V.; Begunova, A.V.; Rozhkova, I.V.; Raskoshnaya, T.A.; Gabrielyan, N.I. Antagonistic Activity of Lactic Acid Bacteria Lactobacillus spp. against Clinical Isolates of Klebsiella pneumoniae. Appl. Biochem. Microbiol. 2018, 54, 277–287. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Moiseenko, K.V.; Glazunova, O.A.; Savinova, O.S.; Vasina, D.V.; Zherebker, A.Y.; Kulikova, N.A.; Nikolaev, E.N.; Fedorova, T.V. Relation between lignin molecular profile and fungal exo-proteome during kraft lignin modification by Trametes hirsuta LE-BIN 072. Bioresour. Technol. 2021, 335, 125229. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Limosilactobacillus reuteri LR1 (GB Accession: GCA_018966925.1) | Lacticaseibacillus rhamnosus F (GB Accession: GCA_018966895.1) | ||||||
|---|---|---|---|---|---|---|---|
| Sequencing | Sequencing | ||||||
| Sequencing technology | Ion Torrent | Number of reads | 3,358,179 | Sequencing technology | Ion Torrent | Number of reads | 3,817,756 |
| Mean read size, bp | 209 | Mean read size, bp | 211 | ||||
| Assembly | Structural annotation | Assembly | Structural annotation | ||||
| Assembly size, bp | 2,053,706 | Genes (total): | 2179 | Assembly size, Mb | 2,893,669 | Genes (total): | 2736 |
| Overall coverage | 100× | -Protein coding | 1947 | Overall coverage | 100× | -Protein coding | 2607 |
| Number of contigs | 319 | -RNA coding | 83 | Number of contigs | 57 | -RNA coding | 71 |
| Longest contig, bp | 64,232 | -Pseudo genes | 149 | Longest contig, bp | 310,149 | -Pseudo genes | 85 |
| N50 contig size, bp | 20,552 | CRISPR arrays | 0 | N50 contig size, bp | 144,365 | CRISPR arrays | 1 |
| Mean contig size, bp | 6437 | Mean contig size, bp | 44,023 | ||||
| Limosilactobacillus reuteri LR1 | ||||
|---|---|---|---|---|
| Protein ID | Functional Description/Possible Function | Functional Category | COG | Secretion |
| MBU5982788.1 | L-lactate dehydrogenase (EC 1.1.1.27) that catalyzes the oxidation of (S)-lactate to the pyruvate. | Metabolism | C | None |
| MBU5983425.1 | Possibly glyoxalase III (EC 4.4.1.5) that catalyzes conversion of methylglyoxal to lactate. | Metabolism | E | None |
| MBU5982312.1 | Cysteine synthase A (EC 2.5.1.47) that catalyzes the formation of cysteine from O-acetylserine through the elimination of acetate and addition of hydrogen sulfide. | Metabolism | E | None |
| MBU5982880.1 | Ornithine carbamoyltransferase (EC 2.1.3.3) that catalyzes the transfer of the carbamoyl group from carbamoyl phosphate to ornithine; produces L-citrulline | Metabolism | E | None |
| MBU5982566.1 | Bacterial solute-binding protein (family 3) that participates in the active transport of amino acids across the cytoplasmic membrane via their delivery to the active-transport system. | Metabolism/Transport | E/T | Classical |
| MBU5982057.1 | Ribonucleoside hydrolase RihC (EC 3.2.2.1) that catalyzes the hydrolysis of ribonucleosides with the formation of free ribose and the corresponding base. | Metabolism | F | None |
| MBU5982646.1 | Nucleotidase (EC 3.1.3.5) that catalyzes the hydrolysis of ribonucleotides to the corresponding ribonucleosides. | Metabolism | F | Classical |
| MBU5983020.1 | Ribonucleoside reductase (EC 1.17.4.1) that catalyzes the oxidation of ribonucleoside-diphosphates to the corresponding deoxyribonucleotides. | Metabolism | F | Non-classical |
| MBU5982282.1 | Phosphoglycerate mutase (EC 5.4.2.11) that catalyzes conversion of bisphosphoglycerate to 3-phosphoglycerate at the eighth step of glycolysis. | Metabolism | G | None |
| MBU5982130.1 | Citrate CoA-transferase (EC 2.8.3.10) that catalyzes the transfer of CoA from Acetyl-CoA to citrate. The enzyme is a component of EC 4.1.3.6 [citrate(pro-3-S)-lyase], which produces acetate and oxaloacetate from citrate. | Metabolism | H | None |
| MBU5982114.1 | Chaperonin GroEL that prevents misfolding and promotes the refolding and proper assembly of unfolded polypeptides generated under stress conditions. | Protein folding | O | None |
| MBU5983126.1 | Peptidase P60 (NlpC/P60; peptidase family C40; EC 3.4...) that catalyzes the hydrolysis of MurNAc-(L-alanine) bonds in peptidoglyucan and/or acts as γ-glutamyl DL-endopeptidase. | Cell wall degradation | M | Classical |
| MBU5982939.1 | Metalloendopeptidase (peptidase family M23; EC 3.4...) that catalyzes the preferential hydrolysis of the glycine–glycine bonds in peptidoglycan. | Cell wall degradation | M | Classical |
| MBU5982718.1 | Mannosyl-glycoprotein endo-beta-N-acetylglucosaminidase (GH73; EC 3.2.1.96) that catalyzes the hydrolysis of the bonds between GlcNAc residues and contiguous monosaccharides in peptidoglycan (contains carbohydrate-binding module family 50; CBM50). | Cell wall degradation | M | Classical |
| MBU5983247.1/MBU5983476.1 | Putative adhesin; contains YSIRK-type signal peptide and LPXTG cell wall anchor domain; has multiple Rib/alpha-like repeats. | Cell wall adhesion | M | Classical |
| MBU5981906.1 | Putative adhesin; contains LPXTG cell wall anchor domain. | Cell wall adhesion | M | Classical |
| Lacticaseibacillus rhamnosus F | ||||
| Protein ID | Functional Description/Possible Function | Functional Category | COG | Secretion |
| MBU5979615.1 | L-lactate dehydrogenase (EC 1.1.1.27) that catalyzes the oxidation of (S)-lactate to the pyruvate. | Metabolism | C | None |
| MBU5977590.1 | D-lactate dehydrogenase (EC 1.1.1.28) that catalyzes the oxidation of (R)-lactate to the pyruvate. | Metabolism | C | None |
| MBU5977926.1 | Malate dehydrogenase (EC 1.1.1.38) that catalyzes the oxidative decarboxylation of malate into pyruvate. | Metabolism | C | None |
| MBU5978913.1 | Glucose-6-phosphate isomerase (EC 5.3.1.9) that catalyzes the isomerization of D-glucose 6-phosphate into D-fructose 6-phosphate at the second step of glycolysis. | Metabolism | G | None |
| MBU5977496.1 | β-galactosidase (GH59; EC 3.2.1.23) that catalyzes the hydrolysis of β-galactosides into monosaccharides (contains FIVAR domain). | Metabolism | G | Classical |
| MBU5978705.1 | Glyceraldehyde-3-phosphate dehydrogenase (type I; EC 1.2.1.12) that catalyzes interconversion of glyceraldehyde-3-phosphate and 1,3-diphosphoglycerate at the sixth step of glycolysis. | Metabolism | G | Non-classical |
| MBU5978932.1 | Zn-dependent protease (peptidase family M10) that is homologous to the human matrix metallopeptidase capable of degrading extracellular matrix proteins. | Proteolysis | NA | Classical |
| MBU5978576.1 | Cell envelope proteinase PrtR (peptidase family S8; EC 3.4.21.96) that degrades the extracellular proteins into oligopeptides. | Proteolysis | O | Classical |
| MBU5978925.1 | Cold-shock protein that binds single-stranded nucleic acids and functions in a variety of processes that are related, for the most part, to post-translational gene regulation | Transcription | K | Non-classical |
| MBU5979459.1 | Cell wall hydrolase P75 (NlpC/P60; peptidase family C40; EC 3.4...) that catalyzes the hydrolysis of MurNAc-(L-alanine) bonds in peptidoglyucan and/or acts as γ-glutamyl DL-endopeptidase. | Cell wall degradation | M | Classical |
| MBU5977346.1 | Lytic transglycosylase (3D domain) that catalyzes cleavage of beta-1-4 bond between MurNAc and GlcNAc. | Cell wall degradation | M | Non-classical |
| MBU5978877.1 | Viral capsid protein derived from the prophage integrated into the bacterial chromosome. | Unknown | NA | Non-classical |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savinova, O.S.; Glazunova, O.A.; Moiseenko, K.V.; Begunova, A.V.; Rozhkova, I.V.; Fedorova, T.V. Exoproteome Analysis of Antagonistic Interactions between the Probiotic Bacteria Limosilactobacillus reuteri LR1 and Lacticaseibacillus rhamnosus F and Multidrug Resistant Strain of Klebsiella pneumonia. Int. J. Mol. Sci. 2021, 22, 10999. https://doi.org/10.3390/ijms222010999
Savinova OS, Glazunova OA, Moiseenko KV, Begunova AV, Rozhkova IV, Fedorova TV. Exoproteome Analysis of Antagonistic Interactions between the Probiotic Bacteria Limosilactobacillus reuteri LR1 and Lacticaseibacillus rhamnosus F and Multidrug Resistant Strain of Klebsiella pneumonia. International Journal of Molecular Sciences. 2021; 22(20):10999. https://doi.org/10.3390/ijms222010999
Chicago/Turabian StyleSavinova, Olga S., Olga A. Glazunova, Konstantin V. Moiseenko, Anna V. Begunova, Irina V. Rozhkova, and Tatyana V. Fedorova. 2021. "Exoproteome Analysis of Antagonistic Interactions between the Probiotic Bacteria Limosilactobacillus reuteri LR1 and Lacticaseibacillus rhamnosus F and Multidrug Resistant Strain of Klebsiella pneumonia" International Journal of Molecular Sciences 22, no. 20: 10999. https://doi.org/10.3390/ijms222010999