

Structure of AQEE-30 of VGF Neuropeptide in Membrane-Mimicking Environments


Abstract
:1. Introduction
2. Results
2.1. Secondary Structure Studies Using CD Spectroscopy and a Helical Wheel Diagram
2.2. Resonance Assignments
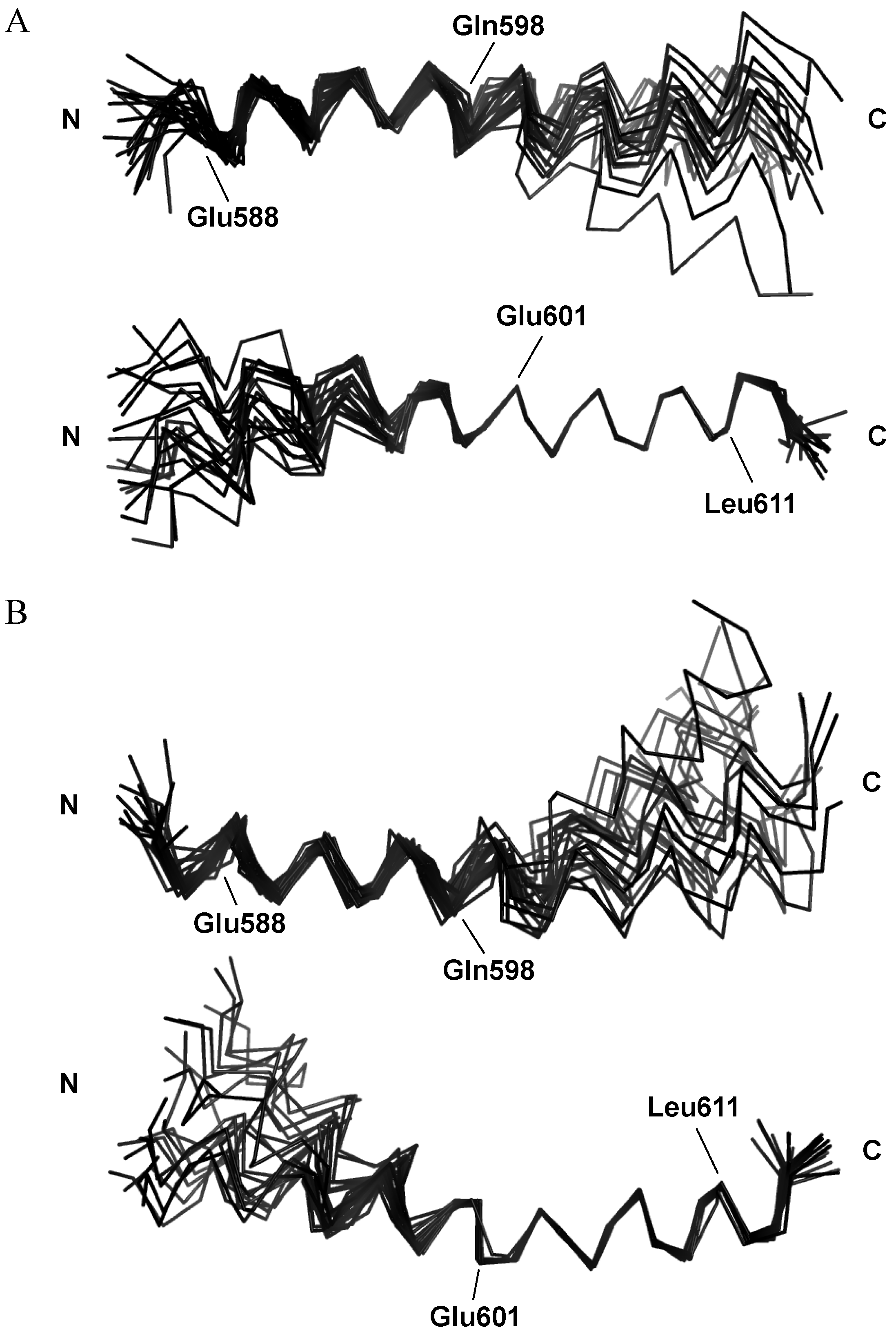
2.3. NMR Structures
3. Discussion
4. Materials and Methods
4.1. Peptide Expression and Purification
4.2. CD Spectroscopy
4.3. NMR Spectroscopy
4.4. Structure Calculations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Levi, A.; Eldridge, J.D.; Paterson, B.M. Molecular cloning of a gene sequence regulated by nerve growth factor. Science 1985, 229, 393–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alder, J.; Thakker-Varia, S.; Bangasser, D.A.; Kuroiwa, M.; Plummer, M.R.; Shors, T.J.; Black, I.B. Brain-derived neurotrophic factor-induced gene expression reveals novel actions of VGF in hippocampal synaptic plasticity. J. Neurosci. 2003, 23, 10800–10808. [Google Scholar] [CrossRef] [Green Version]
- Possenti, R.; Rinaldi, A.M.; Ferri, G.L.; Borboni, P.; Trani, E.; Levi, A. Expression, processing, and secretion of the neuroendocrine VGF peptides by INS-1 cells. Endocrinology 1999, 140, 3727–3735. [Google Scholar] [CrossRef]
- Trani, E.; Giorgi, A.; Canu, N.; Amadoro, G.; Rinaldi, A.M.; Halban, P.A.; Ferri, G.L.; Possenti, R.; Schinina, M.E.; Levi, A. Isolation and characterization of VGF peptides in rat brain. Role of PC1/3 and PC2 in the maturation of VGF precursor. J. Neurochem. 2002, 81, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prigge, S.T.; Mains, R.E.; Eipper, B.A.; Amzel, L.M. New insights into copper monooxygenases and peptide amidation: Structure, mechanism and function. Cell. Mol. Life Sci. 2000, 57, 1236–1259. [Google Scholar] [CrossRef] [PubMed]
- Hahm, S.; Fekete, C.; Mizuno, T.M.; Windsor, J.; Yan, H.; Boozer, C.N.; Lee, C.; Elmquist, J.K.; Lechan, R.M.; Mobbs, C.V.; et al. VGF is required for obesity induced by diet, gold thioglucose treatment, and agouti and is differentially regulated in pro-opiomelanocortin- and neuropeptide Y-containing arcuate neurons in response to fasting. J. Neurosci. 2002, 22, 6929–6938. [Google Scholar] [CrossRef] [Green Version]
- Hahm, S.; Mizuno, T.M.; Wu, T.J.; Wisor, J.P.; Priest, C.A.; Kozak, C.A.; Boozer, C.N.; Peng, B.; McEvoy, R.C.; Good, P.; et al. Targeted deletion of the Vgf gene indicates that the encoded secretory peptide precursor plays a novel role in the regulation of energy balance. Neuron 1999, 23, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Succu, S.; Cocco, C.; Mascia, M.S.; Melis, T.; Melis, M.R.; Possenti, R.; Levi, A.; Ferri, G.L.; Argiolas, A. Pro-VGF-derived peptides induce penile erection in male rats: Possible involvement of oxytocin. Eur. J. Neurosci. 2004, 20, 3035–3040. [Google Scholar] [CrossRef] [PubMed]
- Hunsberger, J.G.; Newton, S.S.; Bennett, A.H.; Duman, C.H.; Russell, D.S.; Salton, S.R.; Duman, R.S. Antidepressant actions of the exercise-regulated gene VGF. Nat. Med. 2007, 13, 1476–1482. [Google Scholar] [CrossRef]
- Cattaneo, A.; Sesta, A.; Calabrese, F.; Nielsen, G.; Riva, M.A.; Gennarelli, M. The expression of VGF is reduced in leukocytes of depressed patients and it is restored by effective antidepressant treatment. Neuropsychopharmacology 2010, 35, 1423–1428. [Google Scholar] [CrossRef]
- Noda, Y.; Shimazawa, M.; Tanaka, H.; Tamura, S.; Inoue, T.; Tsuruma, K.; Hara, H. VGF and striatal cell damage in in vitro and in vivo models of Huntington’s disease. Pharmacol. Res. Perspect. 2015, 3, e00140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, H.; Inagaki, S.; Morozumi, W.; Nakano, Y.; Inoue, Y.; Kuse, Y.; Mizoguchi, T.; Nakamura, S.; Funato, M.; Kaneko, H.; et al. VGF nerve growth factor inducible is involved in retinal ganglion cells death induced by optic nerve crush. Sci. Rep. 2018, 8, 16443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, Y.; Motoyama, S.; Nakamura, S.; Shimazawa, M.; Hara, H. Neuropeptide VGF-Derived Peptide LQEQ-19 has Neuroprotective Effects in an In Vitro Model of Amyotrophic Lateral Sclerosis. Neurochem. Res. 2019, 44, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Thakker-Varia, S.; Jean, Y.Y.; Parikh, P.; Sizer, C.F.; Jernstedt Ayer, J.; Parikh, A.; Hyde, T.M.; Buyske, S.; Alder, J. The neuropeptide VGF is reduced in human bipolar postmortem brain and contributes to some of the behavioral and molecular effects of lithium. J. Neurosci. 2010, 30, 9368–9380. [Google Scholar] [CrossRef] [Green Version]
- Shimazawa, M.; Tanaka, H.; Ito, Y.; Morimoto, N.; Tsuruma, K.; Kadokura, M.; Tamura, S.; Inoue, T.; Yamada, M.; Takahashi, H.; et al. An inducer of VGF protects cells against ER stress-induced cell death and prolongs survival in the mutant SOD1 animal models of familial ALS. PLoS ONE 2010, 5, e15307. [Google Scholar] [CrossRef] [Green Version]
- Schmidlin, T.; Boender, A.J.; Frese, C.K.; Heck, A.J.; Adan, R.A.; Altelaar, A.F. Diet-induced neuropeptide expression: Feasibility of quantifying extended and highly charged endogenous peptide sequences by selected reaction monitoring. Anal. Chem. 2015, 87, 9966–9973. [Google Scholar] [CrossRef]
- Riedl, M.S.; Braun, P.D.; Kitto, K.F.; Roiko, S.A.; Anderson, L.B.; Honda, C.N.; Fairbanks, C.A.; Vulchanova, L. Proteomic analysis uncovers novel actions of the neurosecretory protein VGF in nociceptive processing. J. Neurosci. 2009, 29, 13377–13388. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, T.R.; Tkalych, O.; Nanno, D.; Garcia, A.L.; Devi, L.A.; Salton, S.R. Quantification of VGF- and pro-SAAS-derived peptides in endocrine tissues and the brain, and their regulation by diet and cold stress. Brain Res. 2006, 1089, 21–32. [Google Scholar] [CrossRef]
- Chen, Y.C.; Pristera, A.; Ayub, M.; Swanwick, R.S.; Karu, K.; Hamada, Y.; Rice, A.S.; Okuse, K. Identification of a receptor for neuropeptide VGF and its role in neuropathic pain. J. Biol. Chem. 2013, 288, 34638–34646. [Google Scholar] [CrossRef] [Green Version]
- Hannedouche, S.; Beck, V.; Leighton-Davies, J.; Beibel, M.; Roma, G.; Oakeley, E.J.; Lannoy, V.; Bernard, J.; Hamon, J.; Barbieri, S.; et al. Identification of the C3a receptor (C3AR1) as the target of the VGF-derived peptide TLQP-21 in rodent cells. J. Biol. Chem. 2013, 288, 27434–27443. [Google Scholar] [CrossRef]
- Cero, C.; Vostrikov, V.V.; Verardi, R.; Severini, C.; Gopinath, T.; Braun, P.D.; Sassano, M.F.; Gurney, A.; Roth, B.L.; Vulchanova, L.; et al. The TLQP-21 peptide activates the G-protein-coupled receptor C3aR1 via a folding-upon-binding mechanism. Structure 2014, 22, 1744–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, O.-S.; Bang, J.K.; Ryu, K.-S.; Hwang, E.; Hong, K.S.; Byun, Y.; Cheong, C.; Jeon, Y.H. Structure of neuroendocrine regulatory peptide-2 in membrane-mimicking environments. Pept. Sci. 2020, 113, e24206. [Google Scholar] [CrossRef]
- Kumaran, S.; Roy, R.P. Helix-enhancing propensity of fluoro and alkyl alcohols: Influence of pH, temperature and cosolvent concentration on the helical conformation of peptides. J. Pept. Res. 1999, 53, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.K.; Park, S.H.; Won, H.S.; Na, D.S.; Lee, B.J. Solution structure and membrane-binding property of the N-terminal tail domain of human annexin I. FEBS Lett. 2000, 484, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to study proteins by circular dichroism. Biochim. Biophys. Acta 2005, 1751, 119–139. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, Y.K.; Park, J.W.; Lee, B.; Lee, B.J. Solution structure of the antimicrobial peptide gaegurin 4 by H and 15N nuclear magnetic resonance spectroscopy. Eur. J. Biochem. 2000, 267, 2695–2704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wienk, H.L.; Czisch, M.; de Kruijff, B. The structural flexibility of the preferredoxin transit peptide. FEBS Lett. 1999, 453, 318–326. [Google Scholar] [CrossRef]
- Wray, V.; Mertins, D.; Kiess, M.; Henklein, P.; Trowitzsch-Kienast, W.; Schubert, U. Solution structure of the cytoplasmic domain of the human CD4 glycoprotein by CD and 1H NMR spectroscopy: Implications for biological functions. Biochemistry 1998, 37, 8527–8538. [Google Scholar] [CrossRef]
- Gilbert, G.E.; Baleja, J.D. Membrane-binding peptide from the C2 domain of factor VIII forms an amphipathic structure as determined by NMR spectroscopy. Biochemistry 1995, 34, 3022–3031. [Google Scholar] [CrossRef]
- Wuthrich, K. NMR of Proteins and Nucleic Acids; Wiley: Hoboken, NJ, USA, 1986. [Google Scholar]
- Roccatano, D.; Fioroni, M.; Zacharias, M.; Colombo, G. Effect of hexafluoroisopropanol alcohol on the structure of melittin: A molecular dynamics simulation study. Protein Sci. 2005, 14, 2582–2589. [Google Scholar] [CrossRef]
- Gerig, J.T. Structure and solvation of melittin in 1,1,1,3,3,3-hexafluoro-2-propanol/water. Biophys. J. 2004, 86, 3166–3175. [Google Scholar] [CrossRef] [Green Version]
- Koradi, R.; Billeter, M.; Wuthrich, K. MOLMOL: A program for display and analysis of macromolecular structures. J. Mol. Graph. 1996, 14, 51–55. [Google Scholar] [CrossRef]
- Gimenez-Andres, M.; Copic, A.; Antonny, B. The Many Faces of Amphipathic Helices. Biomolecules 2018, 8, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Lutkenhaus, J. Membrane binding by MinD involves insertion of hydrophobic residues within the C-terminal amphipathic helix into the bilayer. J. Bacteriol. 2003, 185, 4326–4335. [Google Scholar] [CrossRef] [Green Version]
- Lomize, A.L.; Pogozheva, I.D.; Lomize, M.A.; Mosberg, H.I. The role of hydrophobic interactions in positioning of peripheral proteins in membranes. BMC Struct. Biol. 2007, 7, 44. [Google Scholar] [CrossRef] [Green Version]
- Fuglebakk, E.; Reuter, N. A model for hydrophobic protrusions on peripheral membrane proteins. PLoS Comput. Biol. 2018, 14, e1006325. [Google Scholar] [CrossRef]
- Reynaud, J.A.; Grivet, J.P.; Sy, D.; Trudelle, Y. Interactions of basic amphiphilic peptides with dimyristoylphosphatidylcholine small unilamellar vesicles: Optical, NMR, and electron microscopy studies and conformational calculations. Biochemistry 1993, 32, 4997–5008. [Google Scholar] [CrossRef]
- Cohen, C.; Parry, D.A. Alpha-helical coiled coils and bundles: How to design an alpha-helical protein. Proteins 1990, 7, 1–15. [Google Scholar] [CrossRef]
- Barlow, D.J.; Thornton, J.M. Helix geometry in proteins. J. Mol. Biol. 1988, 201, 601–619. [Google Scholar] [CrossRef]
- Chothia, C. Principles that determine the structure of proteins. Annu. Rev. Biochem. 1984, 53, 537–572. [Google Scholar] [CrossRef]
- Kinoshita, M.; Okamoto, Y.; Hirata, F. Peptide Conformations in Alcohol and Water: Analyses by the Reference Interaction Site Model Theory. J. Am. Chem. Soc. 2000, 122, 2773–2779. [Google Scholar] [CrossRef]
- Yoshida, K.; Kawaguchi, J.; Lee, S.; Yamaguchi, T. On the solvent role in alcohol-induced α-helix formation of chymotrypsin inhibitor 2. Pure Appl. Chem. 2008, 80, 1337–1347. [Google Scholar] [CrossRef]
- Nieto, J.L.; Rico, M.; Santoro, J.; Herranz, J.; Bermejo, F.J. Assignment and conformation of neurotensin in aqueous solution by 1H NMR. Int. J. Pept. Protein Res. 1986, 28, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Coutant, J.; Curmi, P.A.; Toma, F.; Monti, J.P. NMR solution structure of neurotensin in membrane-mimetic environments: Molecular basis for neurotensin receptor recognition. Biochemistry 2007, 46, 5656–5663. [Google Scholar] [CrossRef] [PubMed]
- Adam, G.; Delbrück, M. Reduction of dimensionality in biological diffusion processes. Struct. Chem. Mol. Biol. 1968, 198, 198–215. [Google Scholar]
- Sreerama, N.; Woody, R.W. A self-consistent method for the analysis of protein secondary structure from circular dichroism. Anal. Biochem. 1993, 209, 32–44. [Google Scholar] [CrossRef]
- Johnson, W.C. Analyzing protein circular dichroism spectra for accurate secondary structures. Proteins 1999, 35, 307–312. [Google Scholar] [CrossRef]
- Provencher, S.W.; Glockner, J. Estimation of globular protein secondary structure from circular dichroism. Biochemistry 1981, 20, 33–37. [Google Scholar] [CrossRef]
- Sreerama, N.; Woody, R.W. Estimation of protein secondary structure from circular dichroism spectra: Comparison of CONTIN, SELCON, and CDSSTR methods with an expanded reference set. Anal. Biochem. 2000, 287, 252–260. [Google Scholar] [CrossRef]
- Vranken, W.F.; Boucher, W.; Stevens, T.J.; Fogh, R.H.; Pajon, A.; Llinas, M.; Ulrich, E.L.; Markley, J.L.; Ionides, J.; Laue, E.D. The CCPN data model for NMR spectroscopy: Development of a software pipeline. Proteins 2005, 59, 687–696. [Google Scholar] [CrossRef]
- Guntert, P. Automated NMR structure calculation with CYANA. Methods Mol. Biol. 2004, 278, 353–378. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.S.; Maguire, M.L.; Stevens, T.J.; Broadhurst, R.W. DANGLE: A Bayesian inferential method for predicting protein backbone dihedral angles and secondary structure. J. Magn. Reson. 2010, 202, 223–233. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HN | Hα | Hβ | Others | |
|---|---|---|---|---|
| Ala586 | 8.96 | 4.27 | 1.55 | - |
| Gln587 | 8.53 | 4.09 | 2.20, 2.05 | Hγ 2.48; Hε 6.63, 7.43 |
| Glu588 | 7.86 | 4.15 | 2.15, 2.29 | Hγ 2.39, 2.47 |
| Glu589 | 7.92 | 4.15 | 2.22 | Hγ 2.44, 2.56 |
| Ala590 | 8.32 | 4.23 | 1.59 | - |
| Glu591 | 8.15 | 4.10 | 2.25, 2.18 | Hγ 2.38, 2.63 |
| Ala592 | 8.09 | 4.17 | 1.63 | - |
| Glu593 | 8.15 | 4.29 | 2.22, 2.25 | Hγ 2.49 |
| Glu594 | 8.31 | 4.09 | 2.27 | Hγ 2.41, 2.57 |
| Arg595 | 8.18 | 4.18 | 2.07 | Hγ 1.78, 1.88; Hδ 3.30; Hε 7.43; Hη 6.64 |
| Arg596 | 8.25 | 4.17 | 2.11 | Hγ 1.74, 1.89; Hδ 3.24, 3.34; Hε 7.55; Hη 6.71 |
| Leu597 | 8.30 | 4.20 | 1.73, 2.00 | - |
| Gln598 | 8.24 | 4.14 | 2.38, 2.28 | Hγ 2.49, 2.69; Hε 6.53, 7.33 |
| Glu599 | 8.46 | 4.12 | 2.37, 2.20 | Hγ 2.65 |
| Gln600 | 8.30 | 4.09 | 2.34 | Hγ 2.46, 2.58; Hε 6.31, 7.40 |
| Glu601 | 8.36 | 4.16 | 2.29 | Hγ 2.46, 2.61 |
| Glu602 | 8.27 | 4.20 | 2.33, 2.25 | Hγ 2.46, 2.58 |
| Leu603 | 8.37 | 4.34 | 2.03, 1.81 | - |
| Glu604 | 8.65 | 4.00 | 2.24 | - |
| Asn605 | 8.23 | 4.60 | 3.15, 2.94 | Hδ 6.77, 7.75 |
| Tyr606 | 8.50 | 4.33 | 3.49, 3.35 | - |
| Ile607 | 9.03 | 3.58 | 2.05 | - |
| Glu608 | 8.53 | 3.90 | 2.15, 1.86 | - |
| His609 | 8.04 | 4.61 | 3.33 | - |
| Val610 | 8.33 | 3.89 | 1.93 | Hγ 0.85 |
| Leu611 | 8.04 | 4.25 | 1.64, 1.96 | - |
| Leu612 | 8.02 | 4.38 | 2.00, 1.59 | - |
| Arg613 | 7.20 | 4.20 | 1.95 | Hγ 1.66; Hδ 3.24; Hε 7.16 |
| Arg614 | 8.11 | 4.70 | 1.95 | Hγ 1.77; Hδ 3.23; Hε 7.22 |
| Pro615 | - | - | - | - |
| HN | Hα | Hβ | Others | |
|---|---|---|---|---|
| Ala586 | - | 4.21 | 1.42 | - |
| Gln587 | 8.64 | 4.13 | 2.07, 1.94 | Hγ 2.35; Hε 7.76, 6.87 |
| Glu588 | 7.99 | 4.09 | 2.05 | Hγ 2.27 |
| Glu589 | 8.13 | 4.10 | 2.05 | Hγ 2.37, 2.28 |
| Ala590 | 8.18 | 4.21 | 1.44 | - |
| Glu591 | 8.24 | 4.06 | 2.04 | Hγ 2.24, 2.44 |
| Ala592 | 8.08 | 4.11 | 1.48 | - |
| Glu593 | 8.05 | 4.25 | 2.06 | Hγ 2.30 |
| Glu594 | 8.09 | 4.06 | 2.06 | Hγ 2.36 |
| Arg595 | 8.04 | 4.10 | 1.89 | Hγ 1.55; Hδ 3.21; Hε 7.41 |
| Arg596 | 8.09 | 4.10 | 1.90 | Hγ 1.72; Hδ 3.13; Hε 7.54 |
| Leu597 | 8.20 | 4.08 | 1.82, 1.58 | Hδ 0.84, 0.90 |
| Gln598 | 8.11 | 4.09 | 2.16 | Hγ 2.38, 2.51; Hε 7.68, 6.80 |
| Glu599 | 8.44 | 4.06 | 1.99, 2.17 | Hγ 2.48 |
| Gln600 | 8.20 | 4.04 | 2.16 | Hγ 2.32; Hε 6.73, 7.75 |
| Glu601 | 8.16 | 4.08 | 2.10 | Hγ 2.38 |
| Glu602 | 8.26 | 4.08 | 2.08 | Hγ 2.45, 2.28 |
| Leu603 | 8.18 | 4.19 | 1.77, 1.90 | Hγ 1.71; Hδ 0.99, 0.93 |
| Glu604 | 8.59 | 3.81 | 2.17 | Hγ 2.52 |
| Asn605 | 8.41 | 4.42 | 2.96, 2.84 | Hδ 7.79, 7.10 |
| Tyr606 | 8.21 | 4.25 | 3.25 | - |
| Ile607 | 8.54 | 3.49 | 2.00 | Hγ 0.88,1.15; Hδ 0.88 |
| Glu608 | 8.36 | 3.76 | 1.92, 1.77 | Hγ 2.22 |
| His609 | 8.16 | 4.64 | 3.26 | - |
| Val610 | 8.03 | 3.81 | 1.80 | Hγ 0.78, 0.82 |
| Leu611 | 7.74 | 4.15 | 1.52, 1.87 | Hδ 0.92, 0.84 |
| Leu612 | 7.78 | 4.27 | 1.52, 1.98 | Hγ 1.62; Hδ 0.80, 0.83 |
| Arg613 | 7.15 | 4.02 | 1.90, 1.82 | Hγ 1.53; Hδ 3.13; Hε 7.38 |
| Arg614 | 8.20 | 4.50 | 1.87, 1.57 | Hγ 1.67; Hδ 3.13; Hε 7.91 |
| Pro615 | - | 4.21 | 1.89, 2.20 | Hγ 1.96; Hδ 3.77, 3.64 |
| HFIP | DPC | |
|---|---|---|
| dihedral angles | ||
| PHI | 27 | 27 |
| PSI | 27 | 27 |
| distance restraints | ||
| short-range NOEs, |i − j| ≤ 1 | 190 | 178 |
| medium-range NOEs, 1 < |i − j| < 5 | 99 | 61 |
| long-range NOEs, |i − j| ≥ 5 | 0 | 3 |
| total | 289 | 242 |
| average target function value | 0.12 | 0.29 |
| rmsd analysis (Å) | ||
| E601-L611 backbone | 0.08 | 0.23 |
| restraints violated in 6 or more structures | ||
| violated distance restraints | 2 | 1 |
| violated van der Waals restraints | 0 | 0 |
| violated angle restraints | 0 | 0 |
| Ramachandran regions (%) | ||
| most favored | 95.2 | 94.0 |
| additionally allowed | 4.5 | 5.7 |
| generously allowed | 0.0 | 0.0 |
| disallowed | 0.3 | 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, O.-S.; Bang, J.-K.; Cheong, C.; Jeon, Y.-H. Structure of AQEE-30 of VGF Neuropeptide in Membrane-Mimicking Environments. Int. J. Mol. Sci. 2022, 23, 13953. https://doi.org/10.3390/ijms232213953
Park O-S, Bang J-K, Cheong C, Jeon Y-H. Structure of AQEE-30 of VGF Neuropeptide in Membrane-Mimicking Environments. International Journal of Molecular Sciences. 2022; 23(22):13953. https://doi.org/10.3390/ijms232213953
Chicago/Turabian StylePark, One-Sung, Jeong-Kyu Bang, Chaejoon Cheong, and Young-Ho Jeon. 2022. "Structure of AQEE-30 of VGF Neuropeptide in Membrane-Mimicking Environments" International Journal of Molecular Sciences 23, no. 22: 13953. https://doi.org/10.3390/ijms232213953