


New Methylcitrate Synthase Inhibitor Induces Proteolysis, Lipid Degradation and Pyruvate Excretion in Paracoccidioides brasiliensis

 ,
,  ,
,  , , and
, , and 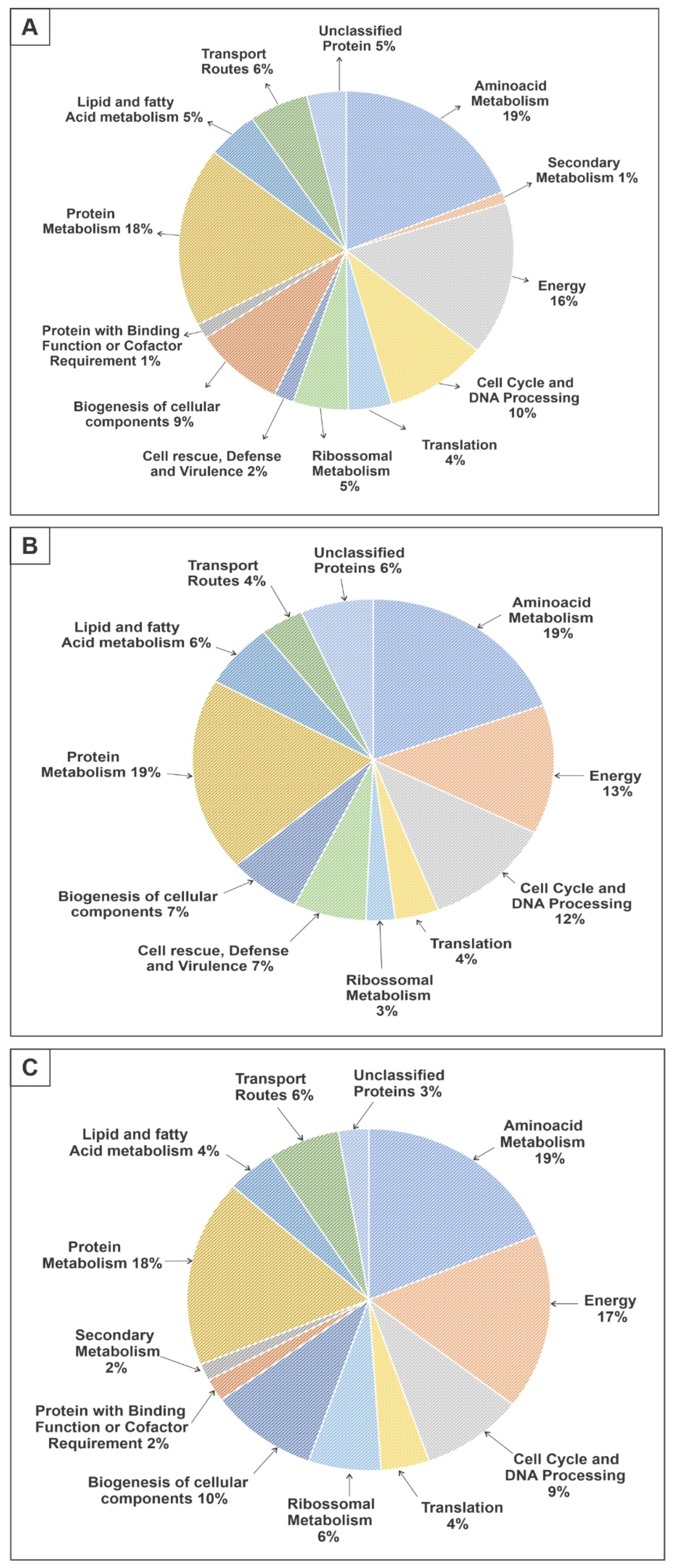{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Microorganism and Culture Conditions
2.3. Obtaining the Protein Extract
2.4. Protein Digestion and NanoUPLC-MSE Analysis
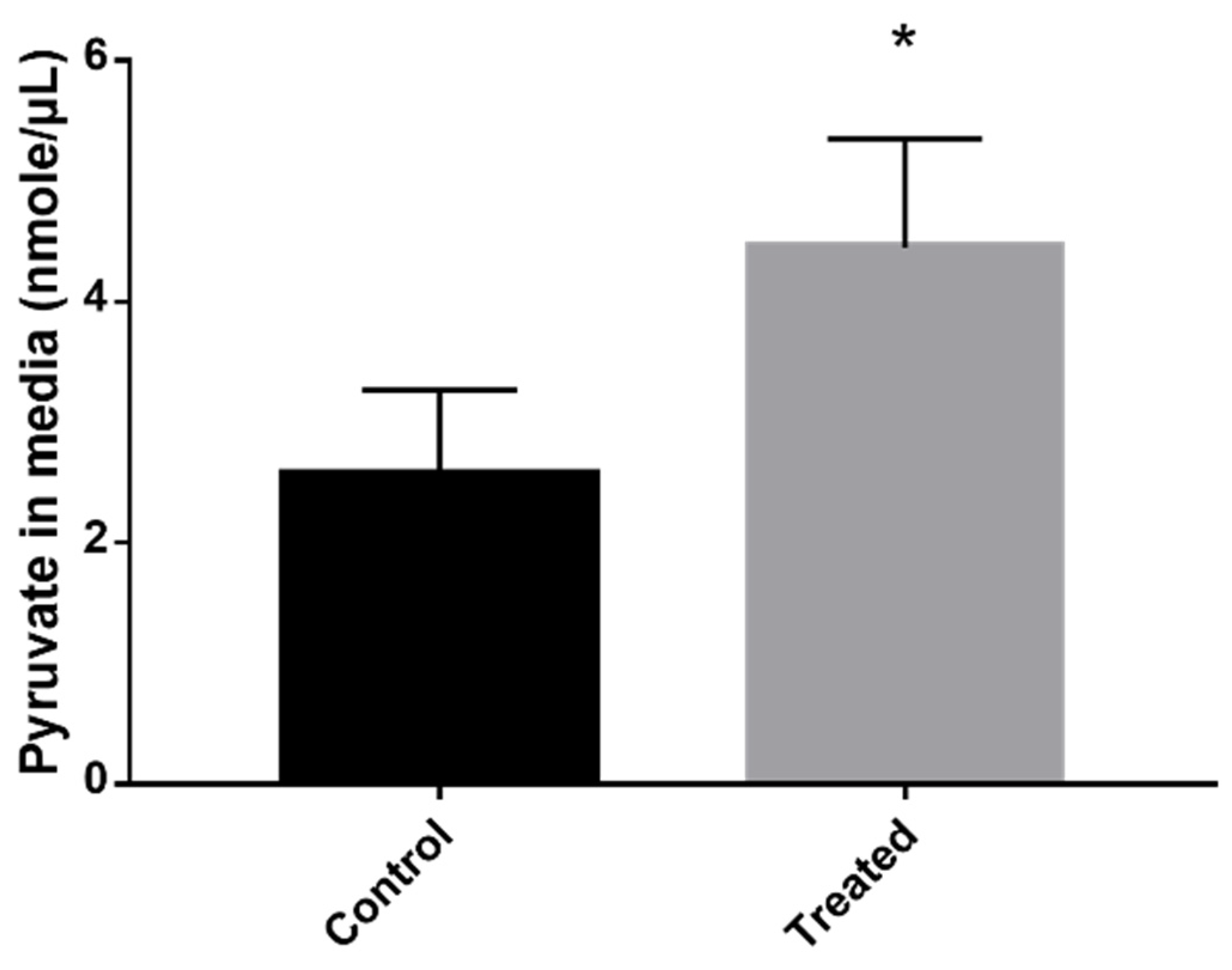
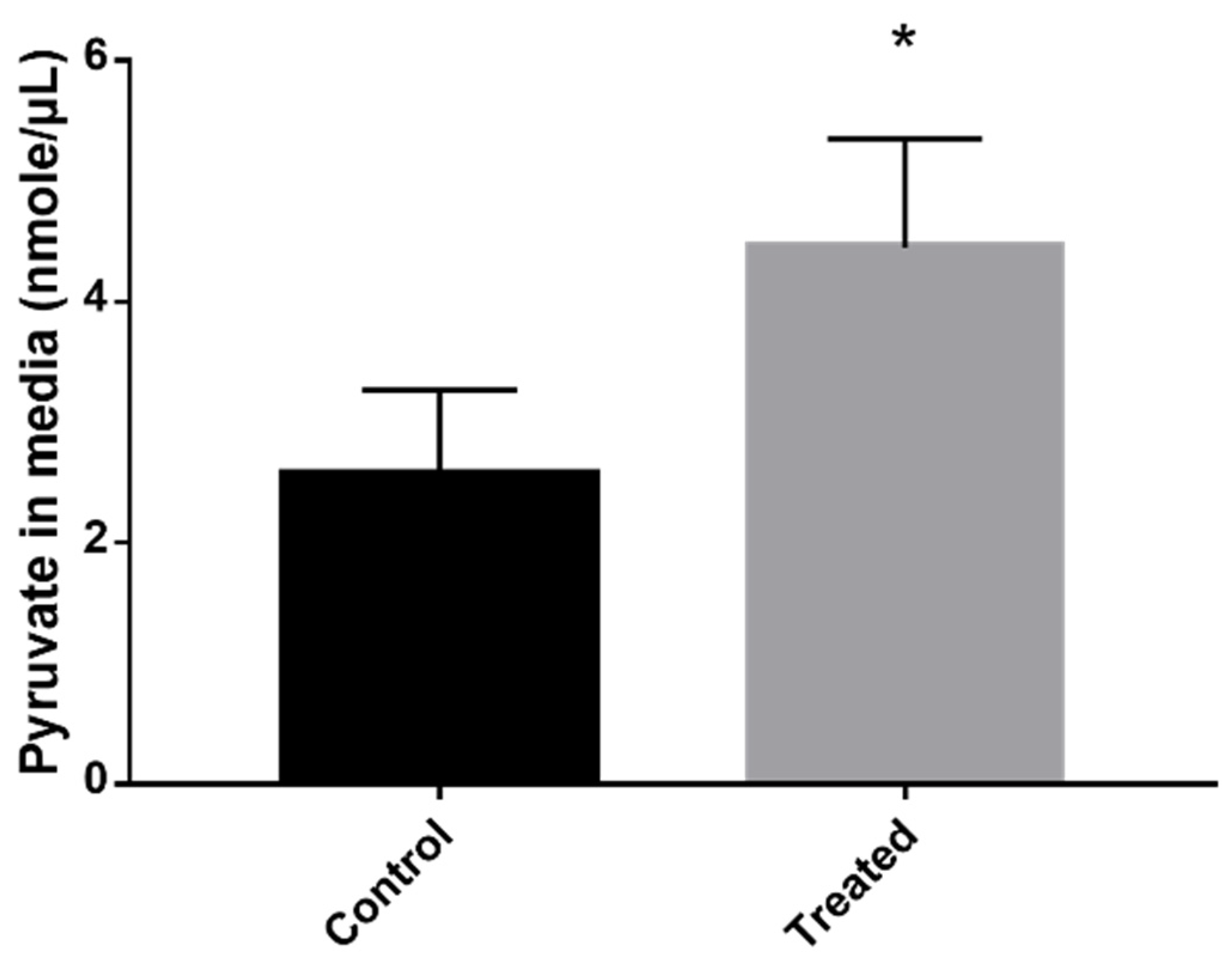
2.5. Pyruvate Dosage
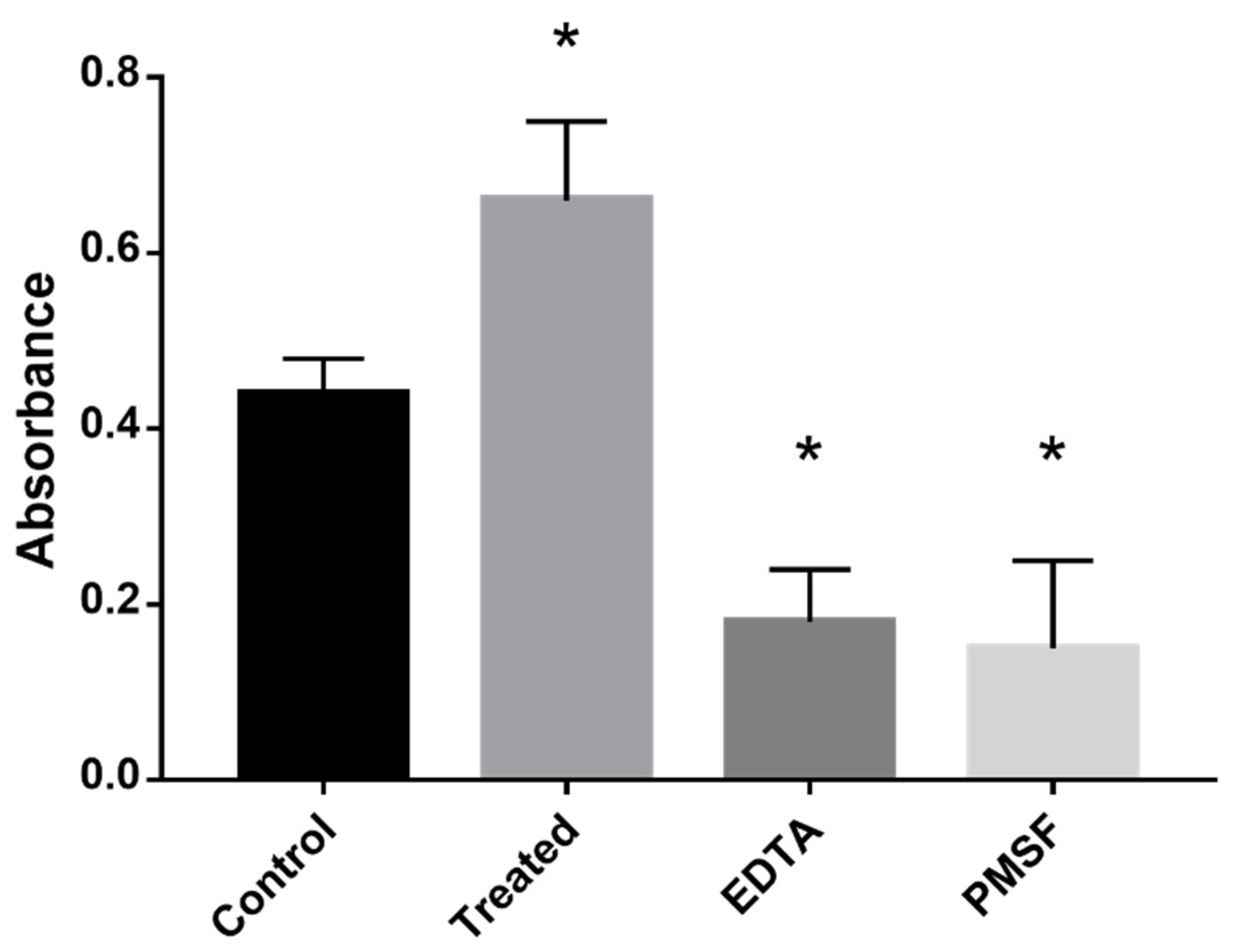
2.6. Azocasein Assay
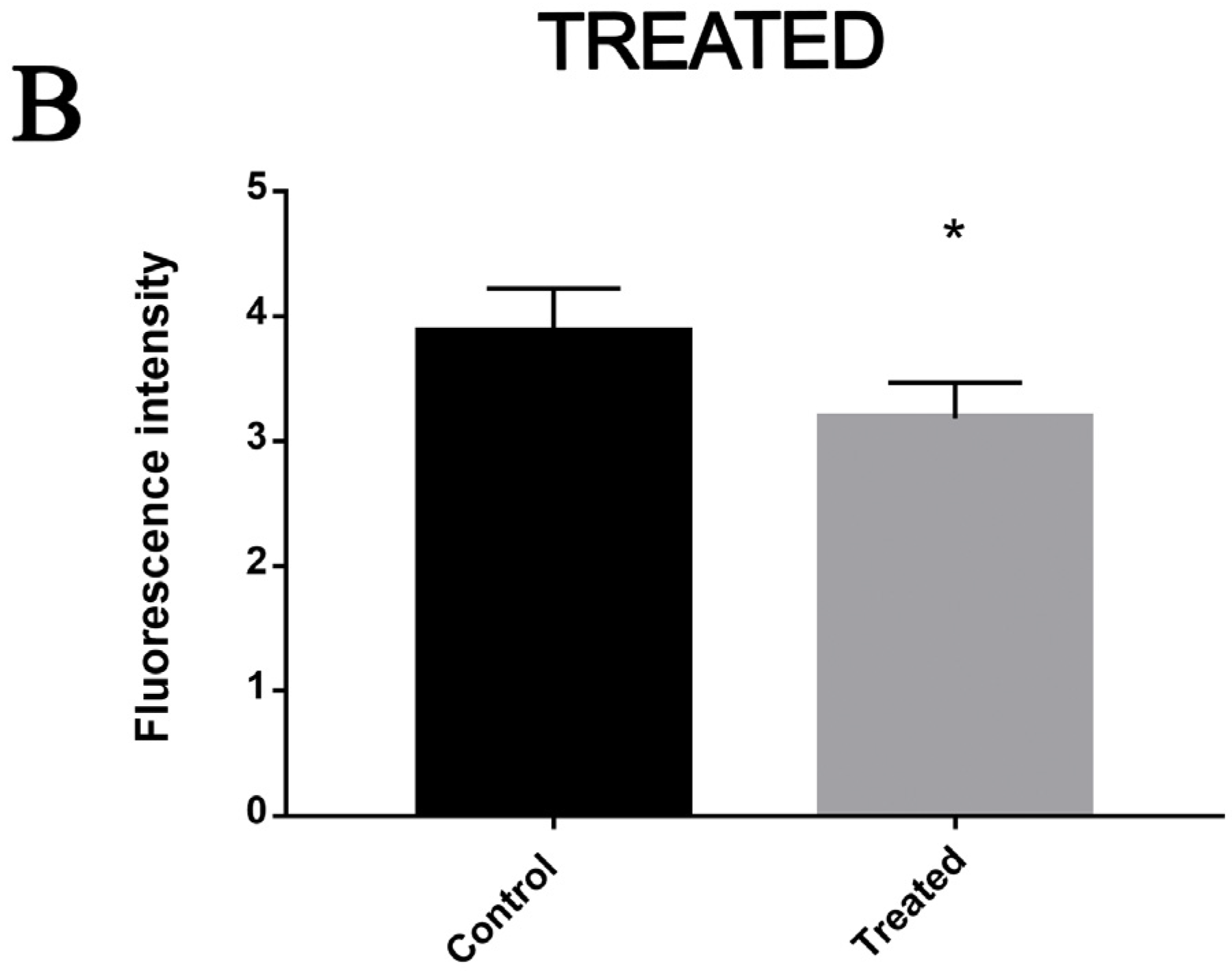
2.7. Dosage of Intracellular Lipids
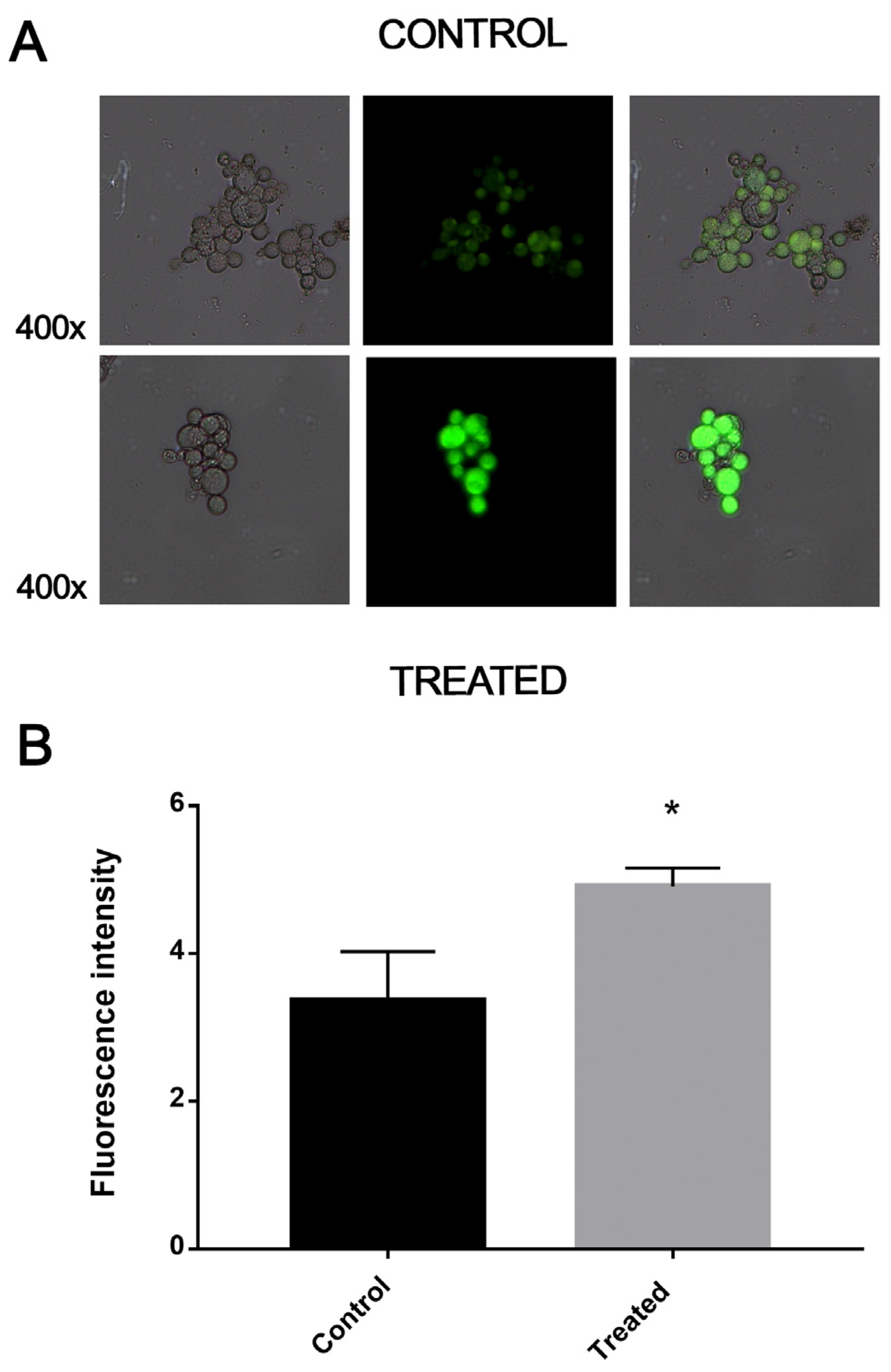
2.8. Quantification of Reactive Oxygen Species
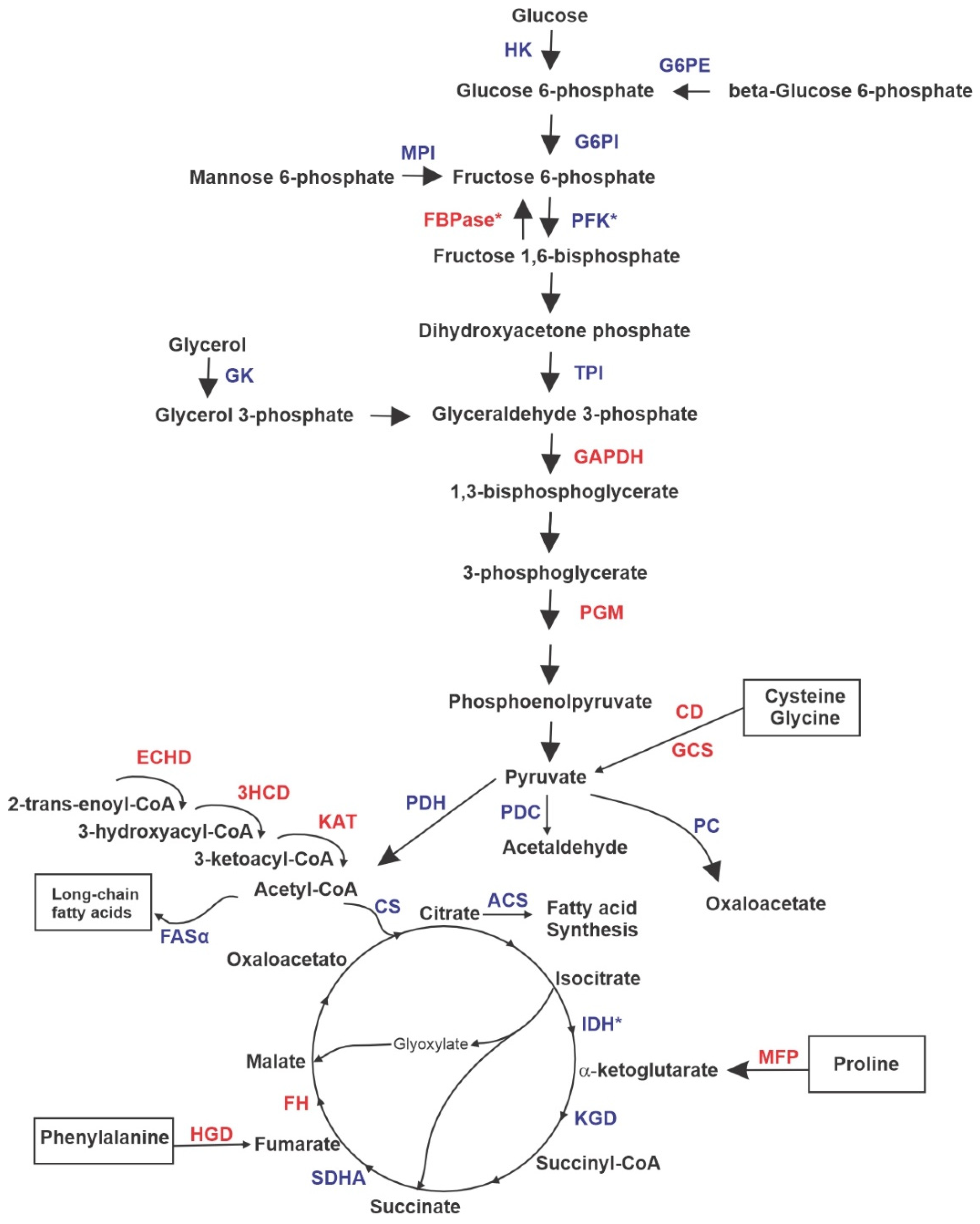
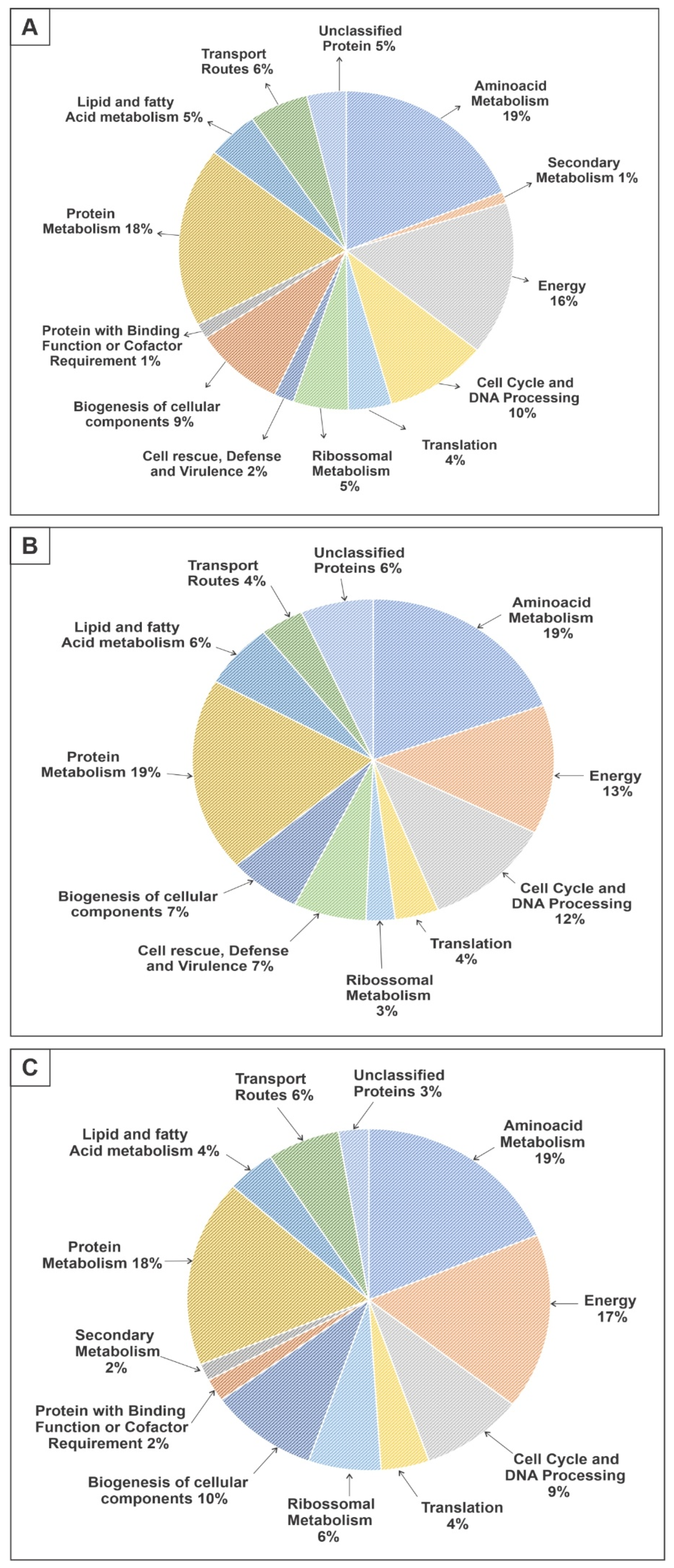
3. Results
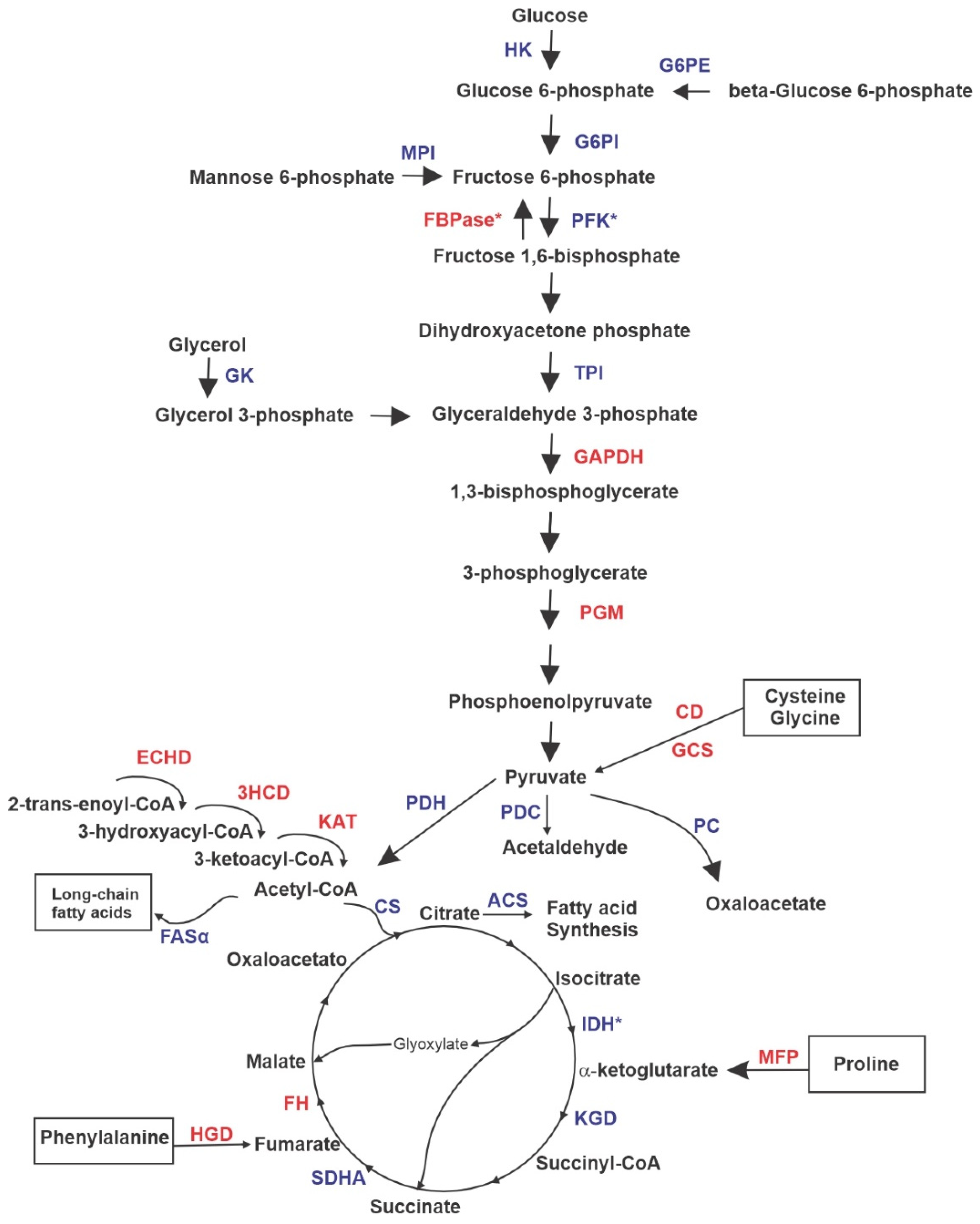
3.1. Proteomic Profile of P. brasiliensis after Treatment with ZINC08964784
3.2. Exposure to MCS Inhibitor Leads to Pyruvate Excretion
3.3. Inhibition of MCS Increases Fungal Proteolytic Activity
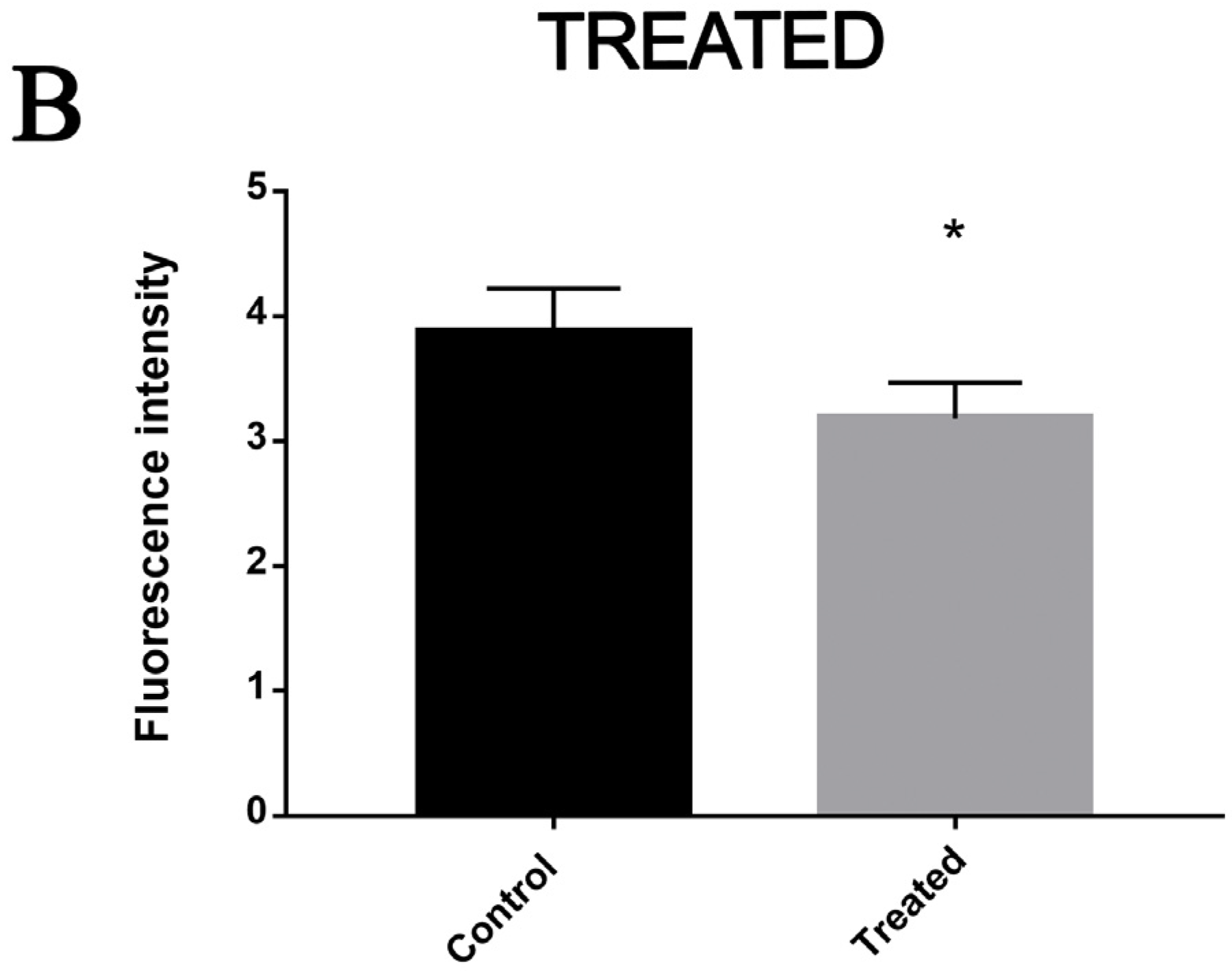
3.4. Exposure to MCS Inhibitor Increases Intracellular Lipid Degradation
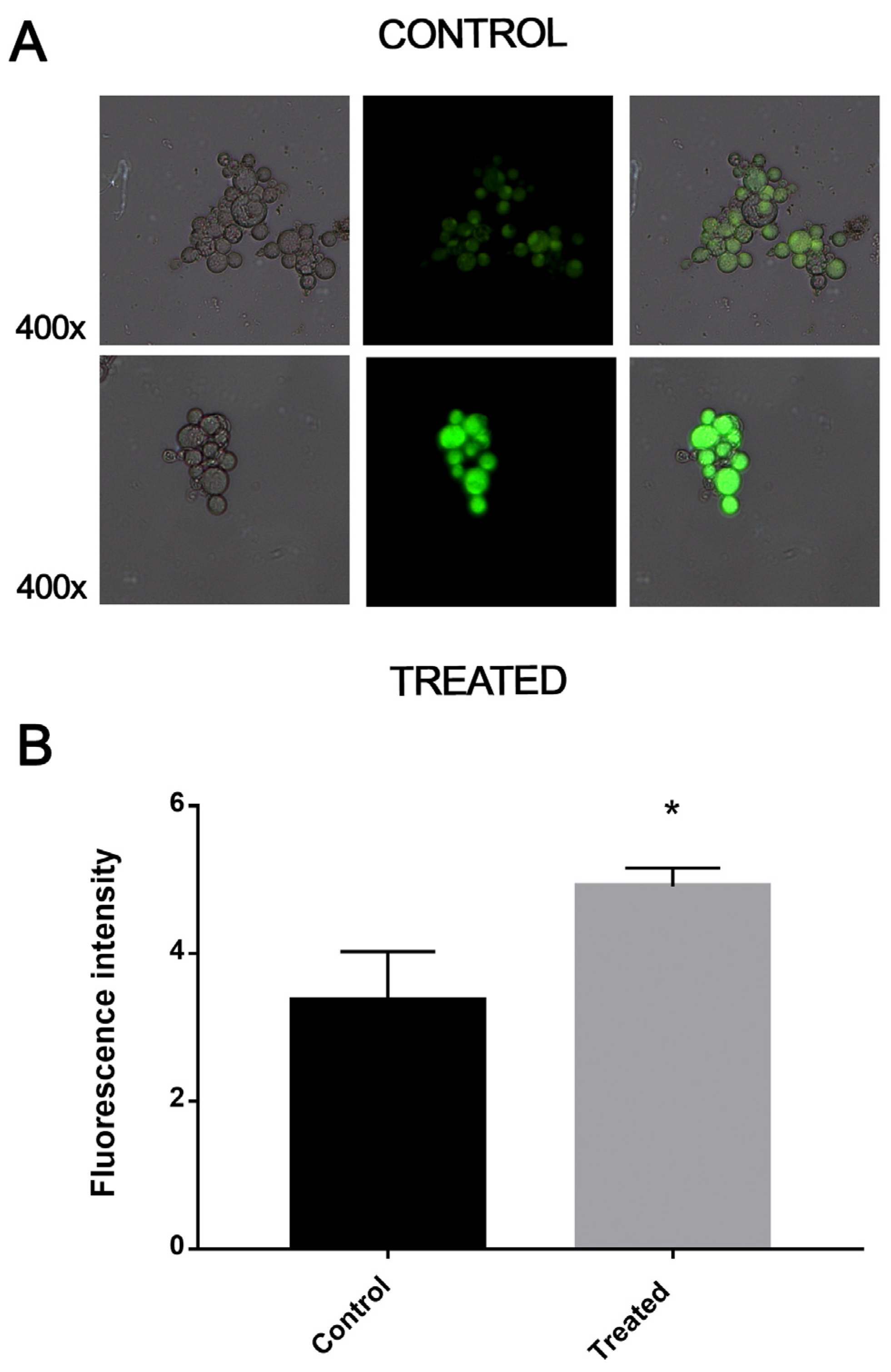
3.5. Exposure to MCS Inhibitor Causes Oxidative Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- de Oliveira, H.C.; Assato, P.A.; Marcos, C.M.; Scorzoni, L.; de Paula, E.; Silva, A.C.A.; Da Silva, J.D.F.; Singulani, J.d.L.; Alarcon, K.M.; Fusco-Almeida, A.M.; et al. Paracoccidioides-Host Interaction: An Overview on Recent Advances in the Paracoccidioidomycosis. Front. Microbiol. 2015, 6, 1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; de Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; do Valle, A.C.F.; Bagagli, E.; Benard, G.; et al. Brazilian Guidelines for the Clinical Management of Paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2017, 50, 715–740. [Google Scholar] [CrossRef] [PubMed]
- Patterson, T.F.; Thompson, G.R., III; Denning, D.W.; Fishman, J.A.; Hadley, S.; Herbrecht, R.; Kontoyiannis, D.P.; Marr, K.A.; Morrison, V.A.; Nguyen, M.H.; et al. Practice Guidelines for the Diagnosis and Management of Aspergillosis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 63, e1–e60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, U.V.; de Oliveira, S.M.d.V.L.; Chang, M.R.; Pereira, E.F.; Marques, A.P.d.C.; de Carvalho, L.R.; Mendes, R.P.; Paniago, A.M.M. Treatment Compliance of Patients with Paracoccidioidomycosis in Central-West Brazil. J. Bras. Pneumol. 2019, 45, e20180167. [Google Scholar] [CrossRef]
- Parente-Rocha, J.A.; Tomazett, M.V.; Pigosso, L.L.; Bailão, A.M.; de Souza, A.F.; Paccez, J.D.; Baeza, L.C.; Pereira, M.; Bailão, M.G.S.; Borges, C.L.; et al. In Vitro, Ex Vivo and in Vivo Models: A Comparative Analysis of Paracoccidioides spp. Proteomic Studies. Fungal Biol. 2018, 122, 505–513. [Google Scholar] [CrossRef]
- Sprenger, M.; Kasper, L.; Hensel, M.; Hube, B. Metabolic Adaptation of Intracellular Bacteria and Fungi to Macrophages. Int. J. Med. Microbiol. 2018, 308, 215–227. [Google Scholar] [CrossRef]
- Brock, M.; Maerker, C.; Schütz, A.; Völker, U.; Buckel, W. Oxidation of Propionate to Pyruvate in Escherichia Coli. Involvement of Methylcitrate Dehydratase and Aconitase. Eur. J. Biochem. 2002, 269, 6184–6194. [Google Scholar] [CrossRef]
- Zimmermann, M.; Kogadeeva, M.; Gengenbacher, M.; McEwen, G.; Mollenkopf, H.-J.; Zamboni, N.; Kaufmann, S.H.E.; Sauer, U. Integration of Metabolomics and Transcriptomics Reveals a Complex Diet of Mycobacterium Tuberculosis during Early Macrophage Infection. mSystems 2017, 2, e00057-17. [Google Scholar] [CrossRef] [Green Version]
- da Silva, L.S.; Barbosa, U.R.; Silva, L.d.C.; Soares, C.M.; Pereira, M.; da Silva, R.A. Identification of a New Antifungal Compound against Isocitrate Lyase of Paracoccidioides Brasiliensis. Future Microbiol. 2019, 14, 1589–1606. [Google Scholar] [CrossRef]
- Costa, F.G.; Neto, B.R.d.S.; Gonçalves, R.L.; da Silva, R.A.; de Oliveira, C.M.A.; Kato, L.; Freitas, C.D.S.; Giannini, M.J.S.M.; da Silva, J.d.F.; Soares, C.M.d.A.; et al. Alkaloids as Inhibitors of Malate Synthase from Paracoccidioides Spp.: Receptor-Ligand Interaction-Based Virtual Screening and Molecular Docking Studies, Antifungal Activity, and the Adhesion Process. Antimicrob. Agents Chemother. 2015, 59, 5581–5594. [Google Scholar] [CrossRef]
- Lima, R.M.; e Silva, K.S.F.; Silva, L.d.C.; Ribeiro, J.F.R.; Neves, B.J.; Brock, M.; Soares, C.M.d.A.; da Silva, R.A.; Pereira, M. A Structure-Based Approach for the Discovery of Inhibitors against Methylcitrate Synthase of Paracoccidioides Lutzii. J. Biomol. Struct. Dyn. 2021, 40, 9361–9373. [Google Scholar] [CrossRef]
- Brock, M.; Buckel, W. On the Mechanism of Action of the Antifungal Agent Propionate: Propionyl-CoA Inhibits Glucose Metabolism in Aspergillus Nidulans. Eur. J. Biochem. 2004, 271, 3227–3241. [Google Scholar] [CrossRef]
- Parente, J.A.; Costa, M.; Pereira, M.; Soares, C.M.d.A. Transcriptome Overview of Paracoccidioides Brasiliensis Proteases. Genet. Mol. Res. 2005, 4, 358–371. [Google Scholar] [PubMed]
- Goughenour, K.D.; Rappleye, C.A. Antifungal Therapeutics for Dimorphic Fungal Pathogens. Virulence 2017, 8, 211–221. [Google Scholar] [CrossRef] [Green Version]
- do Carmo Silva, L.; de Oliveira, A.A.; de Souza, D.R.; Barbosa, K.L.B.; Freitas, E.; Silva, K.S.; Júnior, M.A.B.C.; Rocha, O.B.; Lima, R.M.; Santos, T.G.; et al. Overview of Antifungal Drugs against Paracoccidioidomycosis: How Do We Start, Where Are We, and Where Are We Going? J. Fungi 2020, 6, 300. [Google Scholar] [CrossRef]
- Bueno, P.S.A.; Rodrigues, F.A.V.; Santos, J.L.; Canduri, F.; Biavatti, D.C.; Pimentel, A.L.; Bagatin, M.C.; Kioshima, É.S.; de Freitas Gauze, G.; Seixas, F.A.V. New Inhibitors of Homoserine Dehydrogenase from Paracoccidioides Brasiliensis Presenting Antifungal Activity. J. Mol. Model. 2019, 25, 325. [Google Scholar] [CrossRef] [PubMed]
- Horswill, A.R.; Dudding, A.R.; Escalante-Semerena, J.C. Studies of Propionate Toxicity in Salmonella Enterica Identify 2-Methylcitrate as a Potent Inhibitor of Cell Growth. J. Biol. Chem. 2001, 276, 19094–19101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, L.P.A.; Assunção, L.d.P.; Lima, P.d.S.; Tristão, G.B.; Brock, M.; Borges, C.L.; Silva-Bailão, M.G.; Soares, C.M.d.A.; Bailão, A.M. Propionate Metabolism in a Human Pathogenic Fungus: Proteomic and Biochemical Analyses. IMA Fungus 2020, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Ray, S.C.; Evans, H.M.; Deepe, G.S.; Rappleye, C.A. Metabolism of Gluconeogenic Substrates by an Intracellular Fungal Pathogen Circumvents Nutritional Limitations within Macrophages. mBio 2020, 11, e02712-19. [Google Scholar] [CrossRef] [Green Version]
- Armitt, S.; McCullough, W.; Roberts, C.F. Analysis of Acetate Non-Utilizing (Acu) Mutants in Aspergillus Nidulans. J. Gen. Microbiol. 1976, 92, 263–282. [Google Scholar] [CrossRef]
- Jardón, R.; Gancedo, C.; Flores, C.-L. The Gluconeogenic Enzyme Fructose-1,6-Bisphosphatase Is Dispensable for Growth of the Yeast Yarrowia Lipolytica in Gluconeogenic Substrates. Eukaryot. Cell 2008, 7, 1742–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stipanuk, M.H.; Ueki, I.; Dominy, J.E.; Simmons, C.R.; Hirschberger, L.L. Cysteine Dioxygenase: A Robust System for Regulation of Cellular Cysteine Levels. Amino Acids 2009, 37, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, G.; Motokawa, Y.; Yoshida, T.; Hiraga, K. Glycine Cleavage System: Reaction Mechanism, Physiological Significance, and Hyperglycinemia. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2008, 84, 246–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; St Leger, R.J.; Fang, W. Pyruvate Accumulation Is the First Line of Cell Defense against Heat Stress in a Fungus. mBio 2017, 8, e01284-17. [Google Scholar] [CrossRef] [Green Version]
- Eschrich, D.; Kötter, P.; Entian, K.-D. Gluconeogenesis in Candida Albicans. FEMS Yeast Res. 2002, 2, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Strijbis, K.; Distel, B. Intracellular Acetyl Unit Transport in Fungal Carbon Metabolism. Eukaryot. Cell 2010, 9, 1809–1815. [Google Scholar] [CrossRef] [Green Version]
- Kretschmer, M.; Wang, J.; Kronstad, J.W. Peroxisomal and Mitochondrial β-Oxidation Pathways Influence the Virulence of the Pathogenic Fungus Cryptococcus Neoformans. Eukaryot. Cell 2012, 11, 1042–1054. [Google Scholar] [CrossRef] [Green Version]
- Prigneau, O.; Porta, A.; Poudrier, J.A.; Colonna-Romano, S.; Noël, T.; Maresca, B. Genes Involved in β-Oxidation, Energy Metabolism and Glyoxylate Cycle Are Induced by Candida Albicans during Macrophage Infection. Yeast 2003, 20, 723–730. [Google Scholar] [CrossRef]
- Maerker, C.; Rohde, M.; Brakhage, A.A.; Brock, M. Methylcitrate Synthase from Aspergillus Fumigatus. Propionyl-CoA Affects Polyketide Synthesis, Growth and Morphology of Conidia. FEBS J. 2005, 272, 3615–3630. [Google Scholar] [CrossRef]
- Voltan, A.R.; Donofrio, F.; Miranda, E.T.; Moraes, R.A.; Mendes-Giannini, M.J.S. Induction and Secretion of Elastinolytic and Proteolytic Activity in Cultures of Paracoccidioides Brasiliensis. Rev. De Ciências Farm. Básica E Apl. 2008, 29, 1. [Google Scholar]
- Brock, M.; Fischer, R.; Linder, D.; Buckel, W. Methylcitrate Synthase from Aspergillus Nidulans: Implications for Propionate as an Antifungal Agent. Mol. Microbiol. 2000, 35, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Elías, E.J.; Upton, A.M.; Cherian, J.; McKinney, J.D. Role of the Methylcitrate Cycle in Mycobacterium Tuberculosis Metabolism, Intracellular Growth, and Virulence. Mol. Microbiol. 2006, 60, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Dantas, A.d.S.; Day, A.; Ikeh, M.; Kos, I.; Achan, B.; Quinn, J. Oxidative Stress Responses in the Human Fungal Pathogen, Candida Albicans. Biomolecules 2015, 5, 142–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nittler, M.P.; Hocking-Murray, D.; Foo, C.K.; Sil, A. Identification of Histoplasma Capsulatum Transcripts Induced in Response to Reactive Nitrogen Species. Mol. Biol. Cell 2005, 16, 4792–4813. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha, O.B.; Freitas e Silva, K.S.; Santos, T.G.; Moraes, D.; Assunção, L.d.P.; Bailão, A.M.; Soares, C.M.d.A.; Pereira, M. New Methylcitrate Synthase Inhibitor Induces Proteolysis, Lipid Degradation and Pyruvate Excretion in Paracoccidioides brasiliensis. J. Fungi 2023, 9, 108. https://doi.org/10.3390/jof9010108
Rocha OB, Freitas e Silva KS, Santos TG, Moraes D, Assunção LdP, Bailão AM, Soares CMdA, Pereira M. New Methylcitrate Synthase Inhibitor Induces Proteolysis, Lipid Degradation and Pyruvate Excretion in Paracoccidioides brasiliensis. Journal of Fungi. 2023; 9(1):108. https://doi.org/10.3390/jof9010108
Chicago/Turabian StyleRocha, Olivia Basso, Kleber Santiago Freitas e Silva, Thaynara Gonzaga Santos, Dayane Moraes, Leandro do Prado Assunção, Alexandre Melo Bailão, Célia Maria de Almeida Soares, and Maristela Pereira. 2023. "New Methylcitrate Synthase Inhibitor Induces Proteolysis, Lipid Degradation and Pyruvate Excretion in Paracoccidioides brasiliensis" Journal of Fungi 9, no. 1: 108. https://doi.org/10.3390/jof9010108
APA StyleRocha, O. B., Freitas e Silva, K. S., Santos, T. G., Moraes, D., Assunção, L. d. P., Bailão, A. M., Soares, C. M. d. A., & Pereira, M. (2023). New Methylcitrate Synthase Inhibitor Induces Proteolysis, Lipid Degradation and Pyruvate Excretion in Paracoccidioides brasiliensis. Journal of Fungi, 9(1), 108. https://doi.org/10.3390/jof9010108