
Antiproliferative, Cytotoxic, and Apoptotic Activity of Steroidal Oximes in Cervicouterine Cell Lines

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
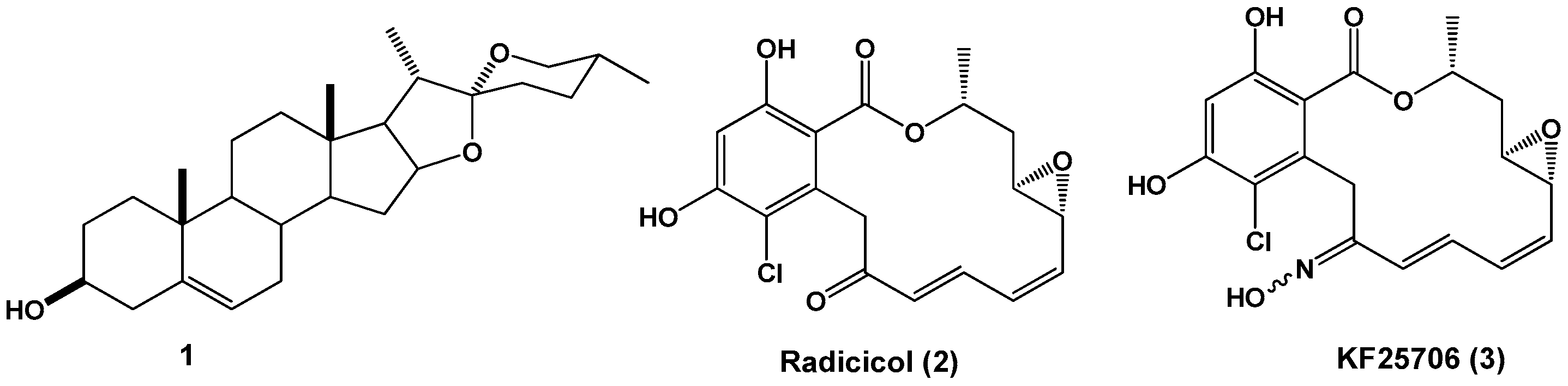
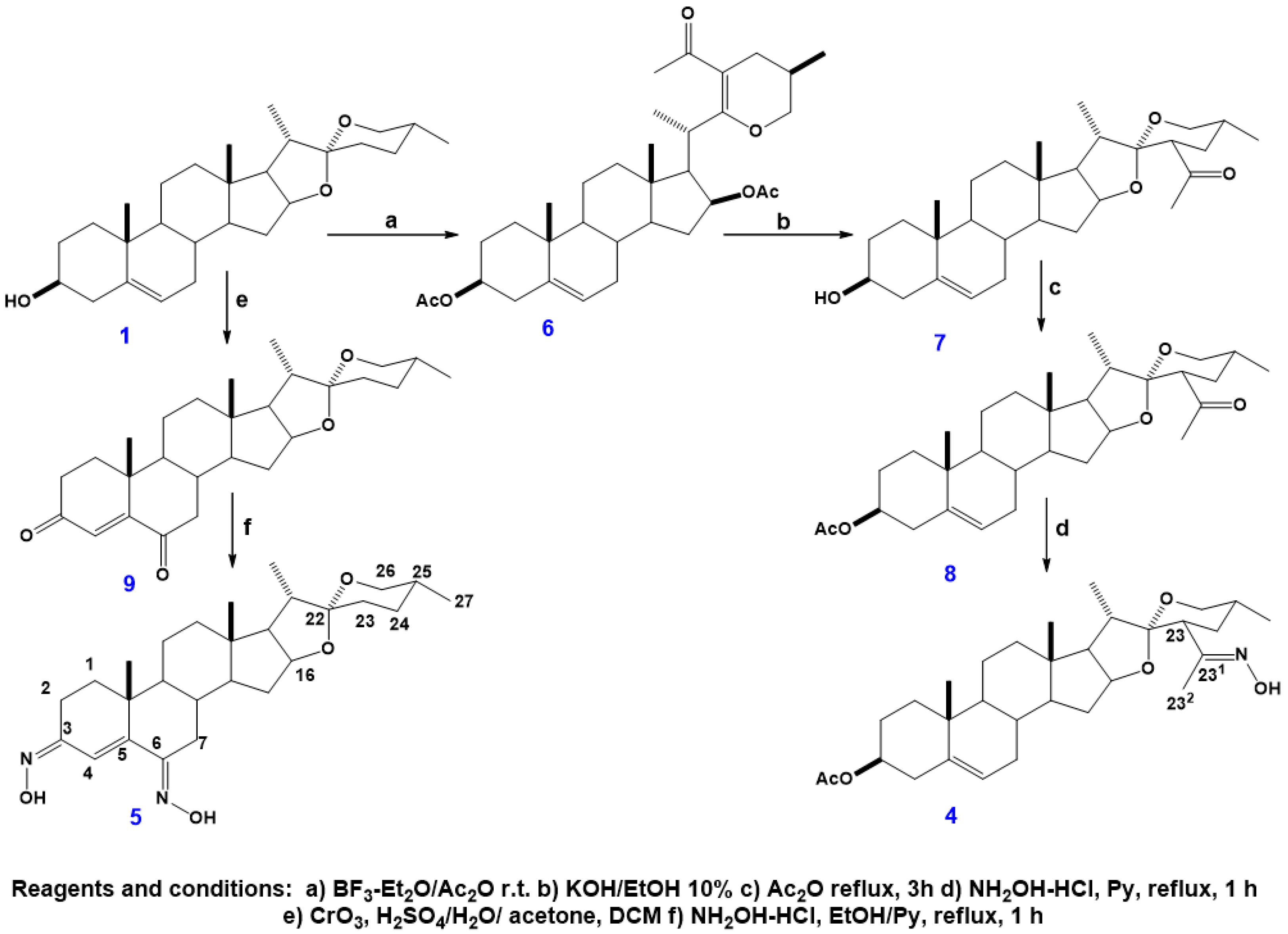
2.1. Synthesis
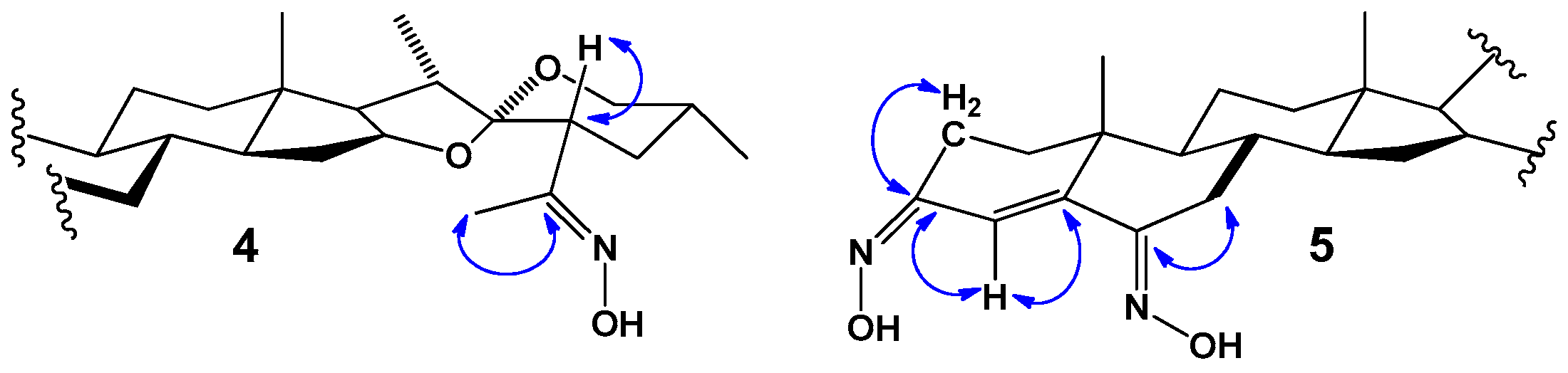
2.2. NMR Analysis
2.3. Biological Activity
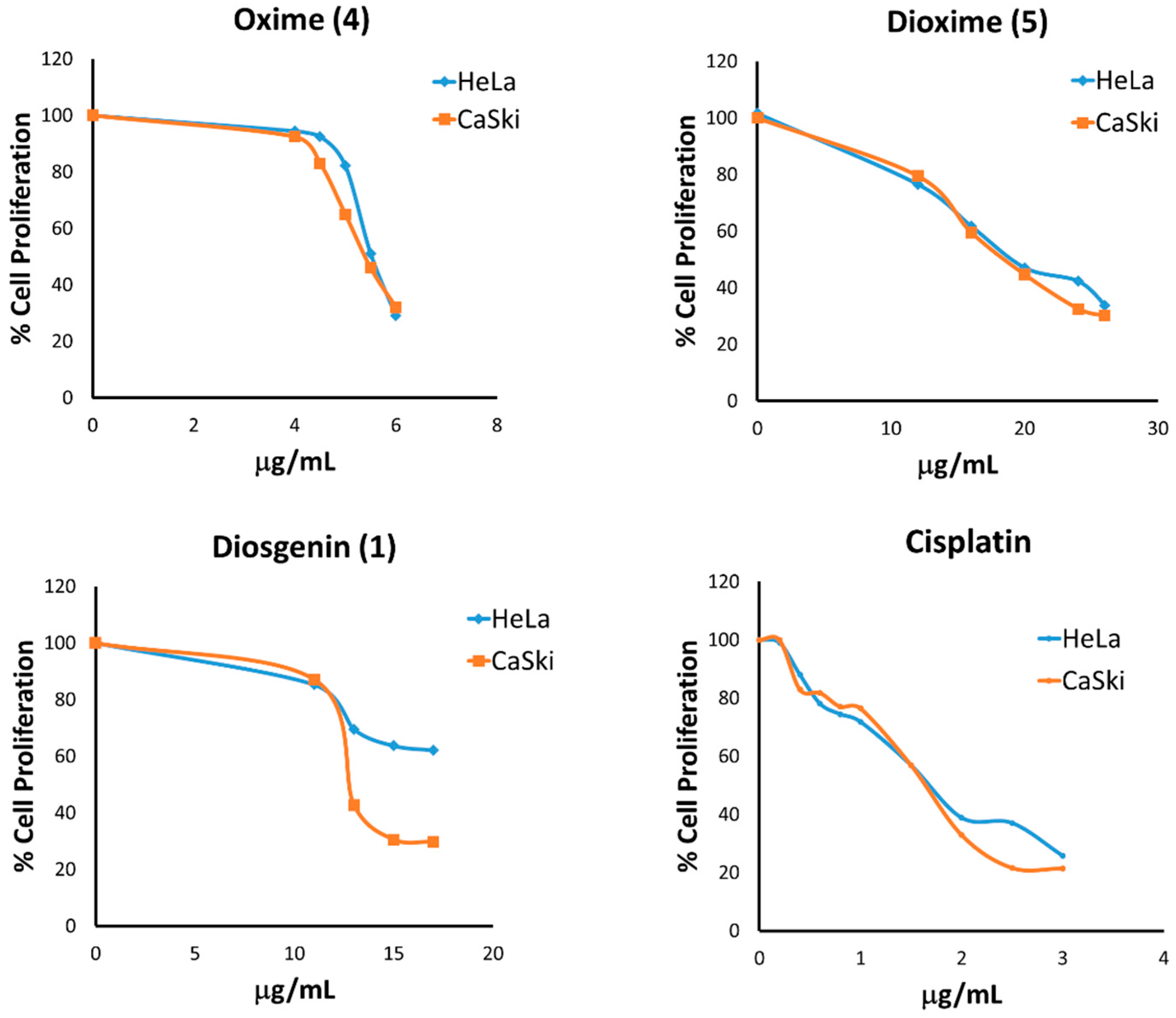
2.3.1. Antiproliferative Activity on HeLa and CaSki Cervical Cancer Cell Lines
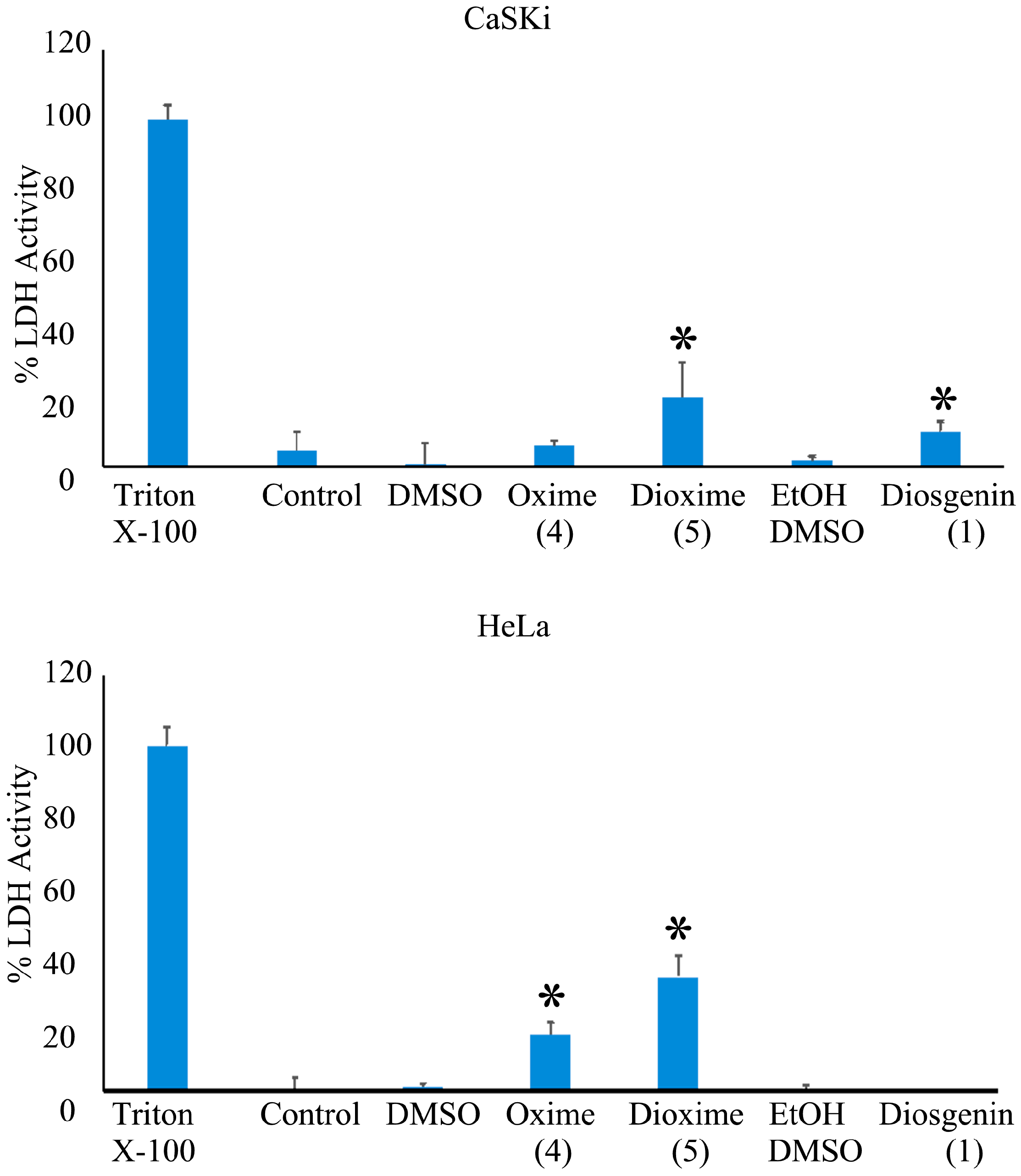
2.3.2. Determination of Cytotoxic Activity on Cervicouterine Cancer Cells
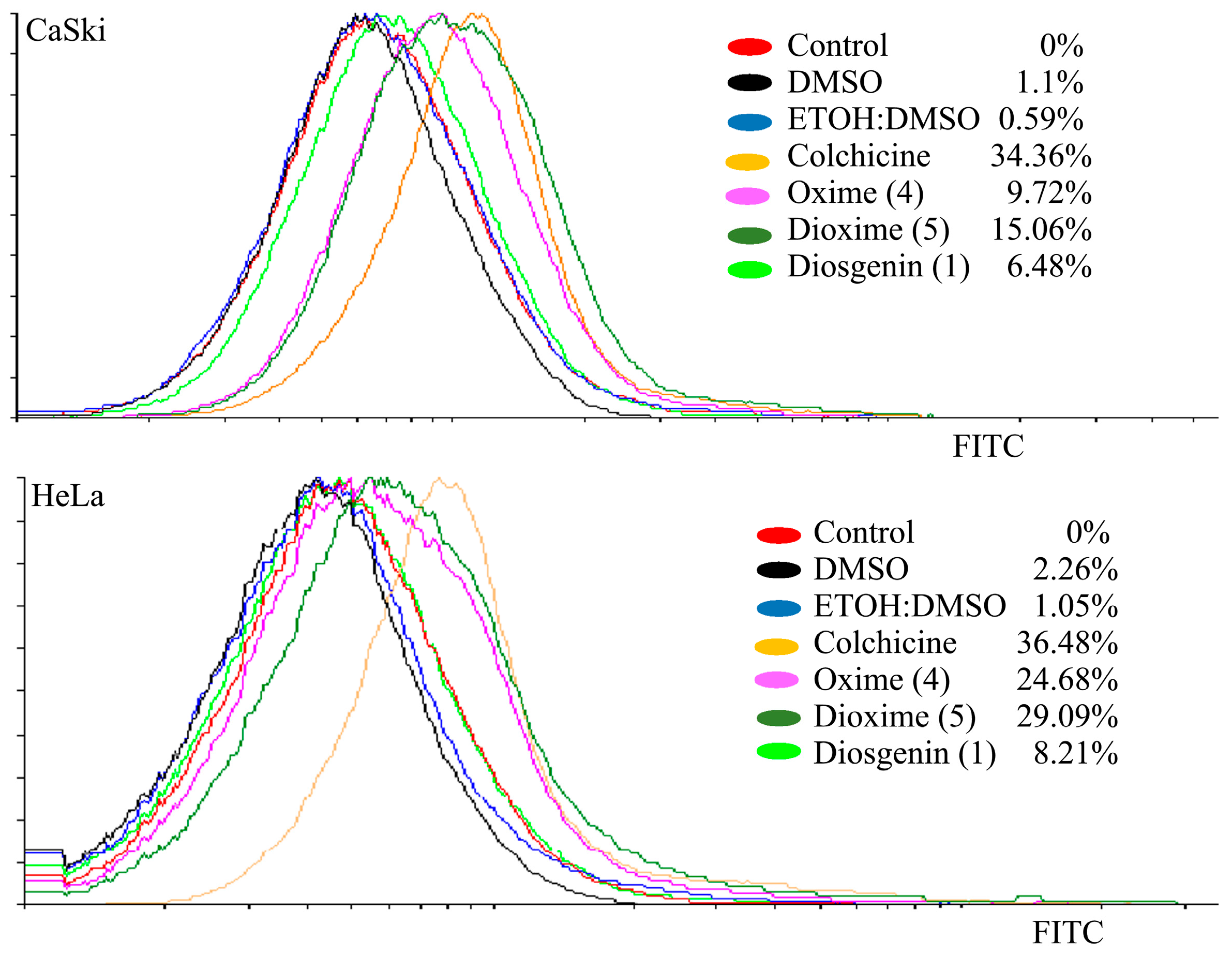
2.3.3. Apoptotic Effect on Tumor Cells
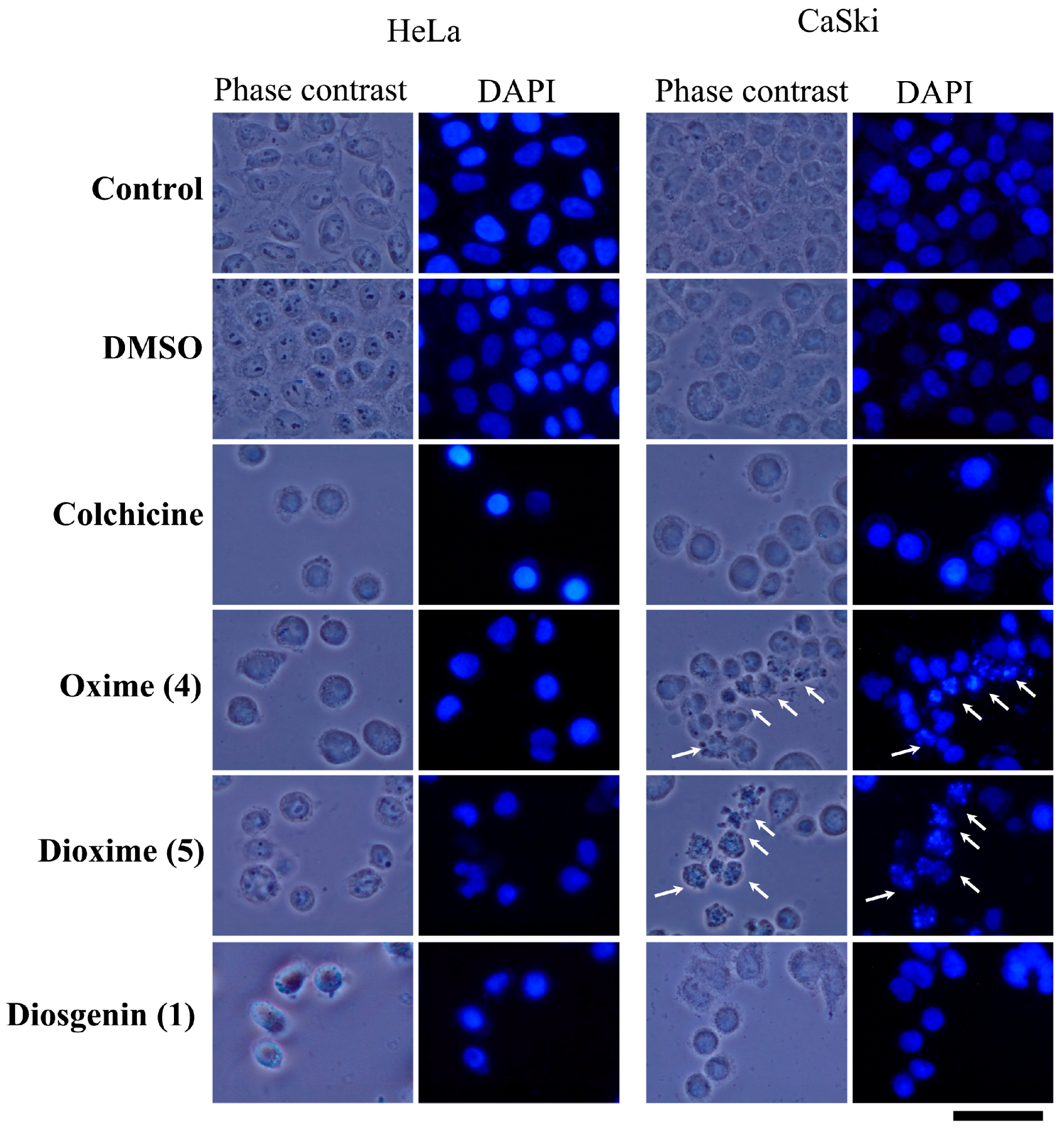
2.3.4. Apoptotic Bodies-DAPI Staining
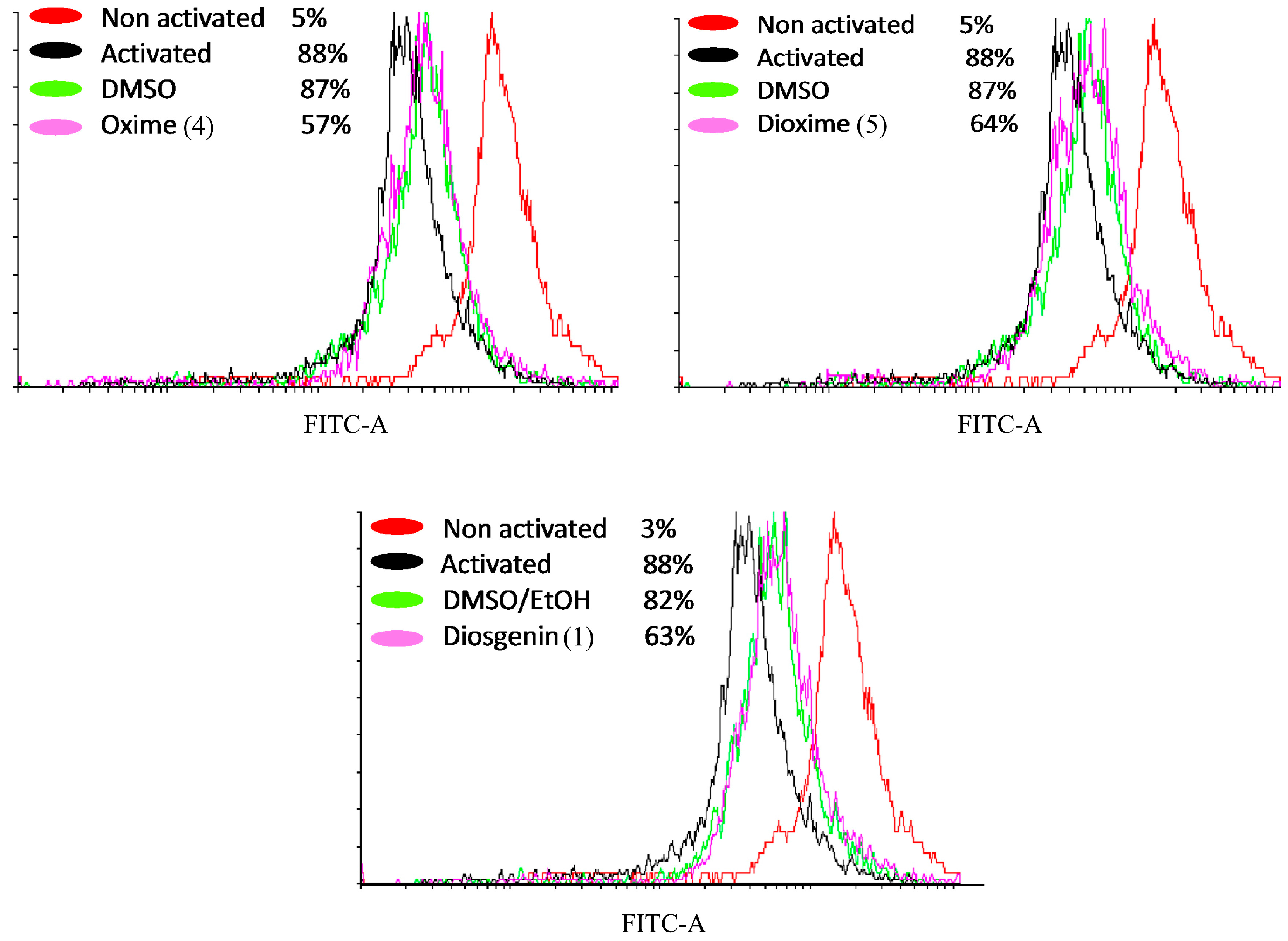
2.3.5. Detection of Active Caspase-3
2.3.6. Evaluation of Antiproliferative Activity on Non-Tumoral Cells
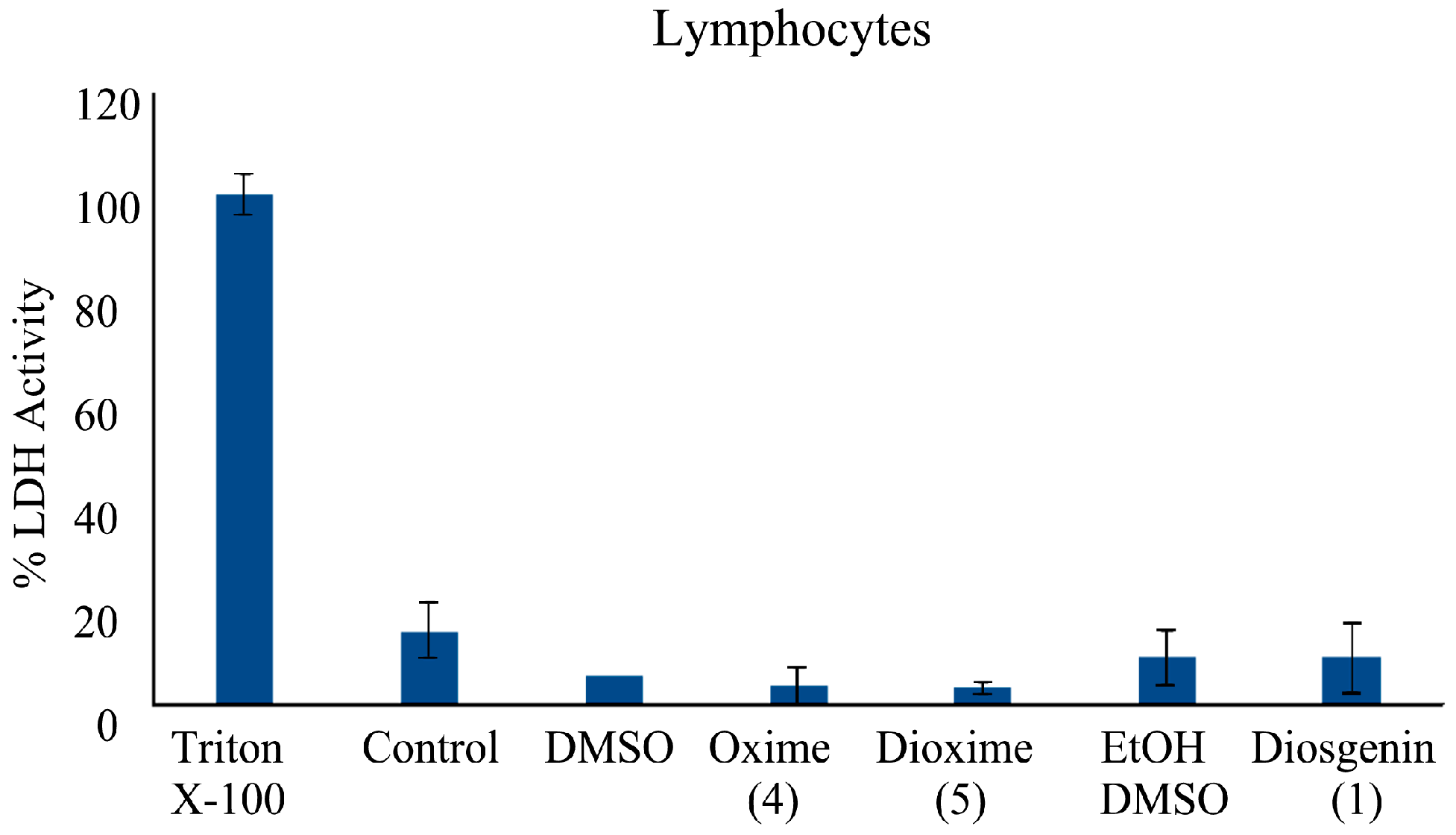
2.3.7. Determination of Cytotoxic Activity on Lymphocyte Cells
3. Materials and Methods
3.1. General Procedures and Materials
3.2. Chemical Synthesis and Characterization
3.2.1. 23-Acetyldiosgenin Acetate 8
3.2.2. Oxime of 23-Acetyldiosgenin Acetate 4
3.2.3. (25R)-Spirost-4-en-3,6-dione 9
3.2.4. Dioxime of (25R)-Spirost-4-en-3,6-dione 5
3.3. Biological Evaluation
3.3.1. Cell Cultures
3.3.2. Cell Proliferation Assays
3.3.3. Determination of Cytotoxicity
3.3.4. CFSE-Labeling Assay
3.3.5. Detection of Active Caspase-3
3.3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Enger, E.D.; Ross, F.C.; Bailey, D.B. Concepts in Biology, 12th ed.; McGraw-Hill: New York, NY, USA, 2007; pp. 171–173. [Google Scholar]
- Kato, A.; Miura, T.; Fukunaga, T. Effects of steroidal glycosides on blood glucose in normal and diabetic mice. Biol. Pharm. Bull. 1995, 18, 167–168. [Google Scholar] [CrossRef] [PubMed]
- Gurib-Fakim, A. Medicinal plants: Traditions of yesterday and drugs of tomorrow. Mol. Asp. Med. 2006, 27, 1–93. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S. Natural products to drugs: Natural product derived compounds in clinical trials. Nat. Prod. Rep. 2005, 22, 162–195. [Google Scholar] [CrossRef] [PubMed]
- Man, S.; Gao, W.; Zhang, Y.; Huang, L.; Liu, C. Chemical study and medical application of saponins as anti-cancer agents. Fitoterapia 2010, 81, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Arunasalam, K.; Yeung, D.; Kakuda, Y.; Mittal, G.; Jiang, Y. Saponins from edible legumes: Chemistry, processing, and health benefits. J. Med. Food 2004, 7, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Djerassi, C.; Rosenkranz, G.; Pataki, J.; Kaufmann, S. Steroids, XXVII. Synthesis of allopregnane-3β, 11β, 17α-, 20β, 21-pentol from cortisone and diosgenin. J. Biol. Chem. 1952, 194, 115–118. [Google Scholar] [PubMed]
- Raju, J.; Mehta, R. Cancer chemopreventive and therapeutic effects of diosgenin, a food saponin. Nutr. Cancer 2008, 61, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Chen, L.; Kou, J.P.; Yu, B.Y. Recent advances of diosgenin in its pharmacological activities and mechanism. Chin. J. Clin. Pharmacol. Ther. 2010, 2, 233–237. [Google Scholar]
- Corbière, C.; Liagre, B.; Bianchi, A.; Bordji, K.; Dauça, M.; Netter, P.; Beneytout, J.L. Different contribution of apoptosis to the antiproliferative effects of diosgenin and other plant steroids, hecogenin and tigogenin, on human 1547 osteosarcoma cells. Int. J. Oncol. 2003, 22, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Corbière, C.; Liagre, B.; Terro, F.; Beneytout, J.L. Induction of antiproliferative effect by diosgenin through activation of p53, release of apoptosis-inducing factor (AIF) and modulation of caspase-3 activity in different human cancer cells. Cell Res. 2004, 14, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.C.; Liu, S.F.; Chang, W.T.; Shiue, Y.L.; Hsieh, P.F.; Hung, T.J.; Hung, C.Y.; Hung, Y.J.; Chen, M.F.; Yang, Y.L. The effects of diosgenin in the regulation of renal proximal tubular fibrosis. Exp. Cell Res. 2014, 323, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.S.; Shih, Y.W.; Huang, H.C.; Cheng, H.W. Diosgenin, a steroidal saponin, inhibits migration and invasion of human prostate cancer PC-3 cells by reducing matrix metalloproteinases expression. PLoS ONE 2011, 6, e20164. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.J.; Tang, Q.J.; Zhang, C.A.; Qin, Z.F.; Pang, B.; Wei, P.W.; Liu, B.; Chou, Y.N. Anti-proliferation and anti-invasion effects of diosgenin on gastric cancer BGC-823 cells with HIF-1α shRNAs. Int. J. Mol. Sci. 2012, 13, 6521–6533. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.H.; Moon, E.; Hong, B.N.; Choi, S.Z.; Son, M.; Park, J.H.; Kim, S.Y. Diosgenin from Dioscorea nipponica ameliorates diabetic neuropathy by inducing nerve growth factor. Biol. Pharm. Bull. 2011, 34, 1493–1498. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Ku, C.Y.; Jan, T.R. Diosgenin attenuates allergen-induced intestinal inflammation and IgE production in a murine model of food allergy. Planta Med. 2009, 75, 1300–1305. [Google Scholar] [CrossRef] [PubMed]
- Jan, T.R.; Wey, S.P.; Kuan, C.C.; Liao, M.H.; Wu, H.Y. Diosgenin, a steroidal sapogenin, enhances antigen-specific IgG2a and interferon-γ expression in ovalbumin-sensitized BALB/c mice. Planta Med. 2007, 73, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Liu, D.Z.; Jan, T.R. Diosgenin, a plant-derived sapogenin, enhances regulatory T-cell immunity in the intestine of mice with food allergy. J. Nat. Prod. 2010, 73, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Chen, L.; Yu, H.; Sun, Q.; Kou, J.; Yu, B. Diosgenin down-regulates NF-κB p65/p50 and p38MAPK pathways and attenuates acute lung injury induced by lipopolysaccharide in mice. Int. Immunopharmacol. 2013, 15, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Huang, B.; Du, D.; Guo, X.; Xin, G.; Xing, Z.; Liang, L.; Chen, Y.; Chen, Q.; He, Y.; et al. Anti-thrombosis effect of diosgenyl saponins in vitro and in vivo. Steroids 2013, 78, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Tian, Y.; Zhang, X.; Bing, B.; Zhang, L.; Wang, H.; Zhao, W. Anti-tumour and immunomodulating activities of diosgenin, a naturally occurring steroidal saponin. Nat. Prod. Res. 2012, 26, 2243–2246. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, T.; Ueno, Y.; Ohta, Z. On the constitution of diosgenin. I. Glucoside of Dioscorea tokoro makino. J. Pharm. Soc. Jpn. 1936, 56, 931–940. [Google Scholar]
- Taylor, W.G.; Elder, J.L.; Chang, P.R.; Richards, K.W. Microdetermination of diosgenin from fenugreek (Trigonella foenum-graecum) seeds. J. Agric. Food Chem. 2000, 48, 5206–5210. [Google Scholar] [CrossRef] [PubMed]
- Hooker, E. Final report of the amended safety assessment of Dioscorea villosa (Wild Yam) root extract. Int. J. Toxicol. 2004, 23, 49–54. [Google Scholar]
- Marker, R.E. Sterols, C.V. The preparation of testosterone and related compounds from sarsasapogenin and diosgenin. J. Am. Chem. Soc. 1940, 62, 2543–2547. [Google Scholar] [CrossRef]
- Tietze, L.F.; Bell, H.P.; Chandrasekhar, S. Natural product hybrids as new leads for drug discovery. Angew. Chem. Int. Ed. 2003, 42, 3996–4028. [Google Scholar] [CrossRef] [PubMed]
- Komesaroff, P.A.; Black, C.V.; Cable, V.; Sudhir, K. Effects of wild yam extract on menopausal symptoms, lipids and sex hormones in healthy menopausal women. Climacteric 2001, 4, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; You-Mei, T.; Su-Lan, Y.; Yu-Wei, H.; Jun-Ping, K.; Bao-Lin, L.; Bo-Yang, Y. Advances in the pharmacological activities and mechanisms of diosgenin. Chin. J. Nat. Med. 2015, 13, 578–587. [Google Scholar]
- Juarez-Oropeza, M.A.; Diaz-Zagoya, J.C.; Rabinowitz, J.L. In vivo and in vitro studies of hypocholesterolemic effects of diosgenin in rats. Int. J. Biochem. 1987, 19, 679–683. [Google Scholar] [CrossRef]
- Huo, R.; Zhou, Q.; Wang, B.; Tashiro, S.; Onodera, S.; Ikejima, T. Diosgenin induces apoptosis in HeLa cells via activation of caspase pathway. Acta Pharmacol. Sin. 2004, 25, 1077–1082. [Google Scholar]
- Raju, J.; Bird, R.P. Diosgenin, a naturally occurring furostanol saponin suppresses 3-hydroxy-3-methylglutaryl CoA reductase expression and induces apoptosis in HCT-116 human colon carcinoma cells. Cancer Lett. 2007, 255, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Moalic, S.; Liagre, B.; Corbière, C.; Bianchi, A.; Dauça, M.; Bordji, K.; Beneytout, J.L. A plant steroid, diosgenin, induces apoptosis, cell cycle arrest and COX activity in osteosarcoma cells. FEBS Lett. 2001, 506, 225–230. [Google Scholar] [CrossRef]
- Li, F.; Fernandez, P.P.; Rajendran, P.; Hui, K.M.; Sethi, G. Diosgenin, a steroidal saponin, inhibits STAT3 signaling pathway leading to suppression of proliferation and chemosensitization of human hepatocellular carcinoma cells. Cancer Lett. 2010, 292, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, X.; Guo, M.; Liu, Y.; Liu, S.; Yao, S. Electrochemical study of breast cancer cells MCF-7 and its application in evaluating the effect of diosgenin. Anal. Sci. 2005, 21, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Bansal, R.; Acharya, P.C. Man-made cytotoxic steroids: Exemplary agents for cancer therapy. Chem. Rev. 2014, 114, 6986–7005. [Google Scholar] [CrossRef] [PubMed]
- Díaz, J.; Durán, N.; Robles, J.; Loaiza, A. Síntesis y evaluación “in vitro” de la actividad antifúngica de oximas, éteres de oxima e isoxazoles. Univ. Sci. 2011, 16, 294–302. [Google Scholar] [CrossRef]
- Soga, S.; Neckers, I.M.; Schulte, T.W.; Shiotsu, Y.; Akasaka, K.; Narumi, H.; Agatsuma, T.; Ikuina, Y.; Murakata, C.; Tamaoki, T.; et al. KF25706, a novel oxime derivative of Radicicol, exhibits in vivo antitumor activity via selective depletion of Hsp90 binding signaling molecules. Cancer Res. 1999, 59, 2931–2938. [Google Scholar] [PubMed]
- Sato, H.; Yamada, R.; Yanagihara, M.; Okuzawa, H.; Iwata, H.; Kurosawa, A.; Ichinomiya, S.; Suzuki, R.; Okabe, H.; Yano, T.; et al. New 2-aryl-1,4-naphthoquinone-1-oxime methyl ether compound induces microtubule depolymerization and subsequent apoptosis. J. Pharmacol. Sci. 2012, 118, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Fan, L.; Huang, Y.; Xin, Y.; Zhou, A. Synthesis and evaluation of some steroidal oximes as cytotoxic agents: Structure/activity studies (I). Steroids 2009, 74, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Fan, L.; Huang, Y.; Xin, Y.; Zhou, A. Synthesis and evaluation of some steroidal oximes as cytotoxic agents: Structure/activity studies (II). Steroids 2009, 74, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.; Nuñez, L.; Peixinho, S.; Jiménez, C. Isolation and synthesis of the first natural 6-hydroximino 4-en-3-one-steroids from the sponges Cinachyrella spp. Tetrahedron Lett. 1997, 38, 1833–1836. [Google Scholar] [CrossRef]
- Ning-Juan, F.; Jiang-Jiang, T.; He, L.; Xiao-Jun, L.; Bo, L.; Jin-Ming, G. Synthesis and cytotoxicity of some novel steroidal C-17 pyrazolinyl derivatives. Eur. J. Med. Chem. 2013, 69, 182–190. [Google Scholar]
- Ning-Juan, F.; Shao-Peng, W.; Jin-Ming, G.; Jiang-Jiang, T. Potential insecticidal activity of steroidal C-17 pyrazolinyl derivatives. J. Braz. Chem. Soc. 2015, 26, 389–392. [Google Scholar]
- Meza-Reyes, S.; Sandoval-Ramírez, J.; Montiel-Smith, S.; Hernández-Linares, G.; Viñas-Bravo, O.; Martínez-Pascual, R.; Fernández-Herrera, M.A.; Vega-Báez, J.L.; Merino-Montiel, P.; Santillán, R.L.; et al. β-Alkoxy-α,β-unsaturated ketone systems in steroidal frameworks, and their conversion to 23,24-bisnorcholane lactones. Arkivoc 2005, vi, 307–320. [Google Scholar]
- Kovganko, N.V.; Chernov, Y.G. 13C-NMR Spectra of 6-hydroximinosteroids of the stigmastane series. Chem. Nat. Compd. 2001, 37, 351–355. [Google Scholar] [CrossRef]
- Cui, J.; Huang, L.; Fan, L.; Zhou, A. A facile and efficient synthesis of some (6E)-hydroximino-4-en-3-one steroids, steroidal oximes from Cinachyrella spp. sponges. Steroids 2008, 73, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Kovganko, N.B.; Chernov, Y.G. Novel synthesis of (24R,6E)-24-ethylcholest-6-hydroxyimino-4-en-3-one, a steroidal oxime from Cinachyrella spp. sponges. Chem. Nat. Compd. 2000, 36, 189–191. [Google Scholar] [CrossRef]
- Holland, H.L.; Kumaresan, S.; Tan, L.; Njar, V.C.O. Synthesis of 6-hydroximino-3-oxo steroids, a new class of aromatase inhibitor. J. Chem. Soc. Perkin Trans. 1 1992, 5, 585–587. [Google Scholar] [CrossRef]
- Deive, N.; Rodríguez, J.; Jiménez, C. Synthesis of cytotoxic 6E-hydroximino-4-ene steroids: Structure/activity studies. J. Med. Chem. 2001, 44, 2612–2618. [Google Scholar] [CrossRef] [PubMed]
- Poza, J.; Rega, M.; Paz, V.; Alonso, B.; Rodríguez, J.; Salvador, N.; Fernández, A.; Jiménez, C. Synthesis and evaluation of new 6-hydroximinosteroid analogs as cytotoxic agents. Bioorg. Med. Chem. 2007, 15, 4722–4740. [Google Scholar] [CrossRef] [PubMed]
- Legrand, C.; Bour, J.M.; Jacob, C.; Capiaumont, J.; Martial, A.; Marc, A.; Wudtke, M.; Kretzmer, G.; Demangel, C.; Duval, D.; et al. Lactate dehydrogenase (LDH) activity of the number of dead cells in the medium of cultured eukaryotic cells as marker. J. Biotechnol. 1992, 25, 231–243. [Google Scholar] [CrossRef]
- Koff, J.L.; Ramachandiran, S.; Bernal-Mizrachi, L. A Time to Kill: Targeting Apoptosis in Cancer. Int. J. Mol. Sci. 2015, 16, 2942–2955. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S. Targeting extrinsic apoptosis in cancer: Challenges and opportunities. Semin. Cell Dev. Biol. 2015, 39, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, H.M. Practical Flow Cytometry, 2nd ed.; Alan R. Liss, Inc.: New York, NY, USA, 1988. [Google Scholar]
- Hartwell, L.H.; Weinert, T.A. Checkpoints: Controls that ensure the order of cell cycle events. Science 1989, 246, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Darzynkiewicz, Z. Nucleic Acid Analysis; Robinson, Ed.; Current Protocols in Cytometry, John Wiley & Sons, Inc.: New York, NY, USA, 1997; Chapter 7. (In Japanese) [Google Scholar]
- Carlson, B.; Lahusen, T.; Singh, S.; Loaiza-Perez, A.; Worland, P.J.; Pestell, R.; Albanese, C.; Sausville, E.A.; Senderowicz, A.M. Down-regulation of cyclin D1 by transcriptional repression in MCF-7 human breast carcinoma cells induced by flavopiridol. Cancer Res. 1999, 59, 4634–4641. [Google Scholar] [PubMed]
- Sherr, C.J. The Pezcoller lecture: Cancer cell cycles revisited. Cancer Res. 2000, 60, 3689–3695. [Google Scholar] [PubMed]
- Senderowicz, A.M.; Sausville, E.A. Preclinical and clinical development of cyclin-dependent kinase modulators. J. Natl. Cancer Inst. 2000, 92, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.R.; Gilmartin, A.; Imburgia, C.; Winkler, J.D.; Marshall, L.A.; Roshak, A. An indolocarbazole inhibitor of human checkpoint kinase (Chk1) abrogates cell cycle arrest caused by DNA damage. Cancer Res. 2000, 60, 566–572. [Google Scholar] [PubMed]
- Hirose, Y.; Berger, M.S.; Pieper, R.O. Abrogation of the Chk1-mediated G2 checkpoint pathway potentiates temozolomide-induced toxicity in a p53- independent manner in human glioblastoma cells. Cancer Res. 2001, 61, 5843–5849. [Google Scholar] [PubMed]
- McLaughlin, F.; Finn, P.; La Thangue, N.B. The cell cycle, chromatin and cancer: Mechanism-based therapeutics come of age. Drug Discov. Today 2003, 8, 793–802. [Google Scholar] [CrossRef]
- Krajewska, M.; Wang, H.G.; Krajewski, S.; Zapata, J.M.; Shabaik, A.; Gascoyne, R.; Reed, J.C. Immunohistochemical analysis of in vivo patterns of expression of CPP32 (Caspase-3), a cell death protease. Cancer Res. 1997, 57, 1605–1613. [Google Scholar] [PubMed]
- Li, Y.; Yao, J.; Chang, M.; Cuendet, M.; Bolton, J.L. Altered apoptotic response in MCF 10A cells treated with the equine estrogen metabolite, 4-hydroxyequilenin. Toxicol. Lett. 2004, 154, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Zhag, H.; Qian, Z.D.; Sun-Tan, Y.; Lee, K.; Gao, P.; Ren, R.Y.; Rey, S.; Hammers, H.; Chang, D.; Pili, R.; et al. Digoxin and other cardiac glycosides inhibit HIL-1 synthesis and block tumor growth. Med. Sci. 2008, 105, 19579–19586. [Google Scholar]
- Dimas, K.; Papadopoulou, N.; Baskakis, C.; Prousis, C.K.; Tsakos, M.; Alkahtani, S.; Honisch, S.; Lang, F.; Calogeropoulou, T.; Alevizopoulos, K.; et al. Steroidal cardiac Na+/K+ ATPase inhibitors exhibit stong anti-cancer potential in vitro and in prostate and lung cancer xenegrafts in vivo. Anti-Cancer Agents Med. Chem. 2014, 14, 762–770. [Google Scholar] [CrossRef]
- Alevizopoulos, K.; Calogeropoulou, T.; Lang, F.; Stournaras, C. Na+/K+ ATPase Inhibitors in Cancer. Curr. Drug Targets 2014, 15, 988–1000. [Google Scholar] [PubMed]
- Gobbini, M.; Armanoli, S.; Banfi, L.; Benicchio, A.; Carzana, G.; Fedrizzzi, G.; Ferrari, P.; Giacalone, G.; Giubileo, M.; Narazzi, G.; et al. Novel analogues of Istaroxime a potent inhibitor of Na+/K+-ATPase: Synthesis and structure-activity relationship. J. Med. Chem. 2008, 51, 4601–4608. [Google Scholar] [CrossRef] [PubMed]
- Alevizopoulos, K.; Dimas, K.; Papadopoulou, N.; Schmidt, E.M.; Tsapara, A.; Alkahtani, S.; Honisch, S.; Prousis, C.K.; Alarifi, S.; Calogeropoulou, T.; et al. Funtional characterization and anti-cancer action of the clinical phase II cardiac Na+/K+ ATPase inhibitor istaroxime: In vitro and in vivo properties and cross talk with the membrane androgen receptor. Oncotarget 2016, 7, 24415–24428. [Google Scholar] [PubMed]
- Raju, J.; Rao, C.V. Diosgenin, a Steroid Saponin Constituent of Yams and Fenugreek: Emerging Evidence for Applications in Medicine. Bioactive Compounds in Phytomedicine, Prof; Rasooli, I., Ed.; InTech.: Rijeka, Croatia, 2012; pp. 125–142. [Google Scholar]
- Kueng, W.; Silber, E.; Eppenberger, U. Quantification of cells cultured on 96-well plates. Anal. Biochem. 1989, 182, 16–19. [Google Scholar] [CrossRef]
- Lyons, A.B.; Hasbold, J.; Hodgkin, P.D. Chapter 17 Flow cytometric analysis of cell division history using dilution of carboxyfluorescein diacetate succinimidyl ester, a stably integrated fluorescent probe. Methods Cell Biol. 2001, 63, 375–398. [Google Scholar] [PubMed]
- Thareja, S. Steroidal 5α-Reductase inhibitors: A comparative 3D-QSAR study review. Chem. Rev. 2015, 115, 2883–2894. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds 1, 4–9 are available from the authors.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell line | IC50 μg/ML (μM) | ||
|---|---|---|---|
| Diosgenin (1) | Oxime (4) | Dioxime (5) | |
| HeLa | 15 (36.17) | 5.6 (10.9) | 19 (41.61) |
| CaSki | 13 (31.35) | 5.4 (10.51) | 22 (48.18) |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Sánchez, L.; Hernández-Linares, M.G.; Escobar, M.L.; López-Muñoz, H.; Zenteno, E.; Fernández-Herrera, M.A.; Guerrero-Luna, G.; Carrasco-Carballo, A.; Sandoval-Ramírez, J. Antiproliferative, Cytotoxic, and Apoptotic Activity of Steroidal Oximes in Cervicouterine Cell Lines. Molecules 2016, 21, 1533. https://doi.org/10.3390/molecules21111533
Sánchez-Sánchez L, Hernández-Linares MG, Escobar ML, López-Muñoz H, Zenteno E, Fernández-Herrera MA, Guerrero-Luna G, Carrasco-Carballo A, Sandoval-Ramírez J. Antiproliferative, Cytotoxic, and Apoptotic Activity of Steroidal Oximes in Cervicouterine Cell Lines. Molecules. 2016; 21(11):1533. https://doi.org/10.3390/molecules21111533
Chicago/Turabian StyleSánchez-Sánchez, Luis, María Guadalupe Hernández-Linares, María L. Escobar, Hugo López-Muñoz, Edgar Zenteno, María A. Fernández-Herrera, Gabriel Guerrero-Luna, Alan Carrasco-Carballo, and Jesús Sandoval-Ramírez. 2016. "Antiproliferative, Cytotoxic, and Apoptotic Activity of Steroidal Oximes in Cervicouterine Cell Lines" Molecules 21, no. 11: 1533. https://doi.org/10.3390/molecules21111533