1. Introduction
Dengue virus (DENV) infection is one of the most rapidly spreading arboviral diseases of humans in the world [
1]. It is a major health problem among those dwelling in tropical and subtropical areas, and it is spreading globally, with approximately 129 countries now thought to be DENV-endemic areas [
2]. The incidence of DENV infection continues to increase. An estimated 100–400 million people are infected annually, and 96 million people require medical care [
3]. There is currently no specific drug to treat DENV infection. Supportive treatment is the only treatment option for hospitalized DENV-infected patients.
DENV is a member of the Flaviviridae family, which includes four closely related serotypes, including DENV1, DENV2, DENV3, and DENV4 [
1]. All four serotypes are usually cocirculating in DENV-endemic areas, and the virus infects humans via the bite of female
Aedes aegypti or
Aedes albopictus mosquitoes, causing clinical manifestations that vary from mild to severe [
4]. Subsequent infections with serotypes different from the primary infection were reported to promote severe disease, and this is the major obstacle to DENV vaccine development [
4]. A tetravalent vaccine against all four DENV serotypes represents the ideal model for protecting against DENV infection and minimizing its severity [
5]. Attempts to develop an effective dengue vaccine have been ongoing for decades, and these attempts have yielded one currently available licensed vaccine. Sanofi Pasteur’s Dengvaxia
® vaccine has been distributed to vaccinate people against DENV infection in more than 20 countries worldwide [
6]. However, the overall efficacy of this vaccine was reported to be limited with suboptimal protection against DENV1 and DENV2 (50% and 35–42% protection, respectively) [
7,
8,
9]. Although the availability of a DENV vaccine is necessary for the prevention and control of viral infection, anti-DENV drugs for the treatment of DENV infection are also important for preventing disease progression, for reducing disease severity, and for interrupting the spread of the virus.
Biological resources in and from nature harbor large and diverse assortments of bioactive compounds [
10]. Natural compounds from herbs and medicinal plants are recognized for their safety and effectiveness, and many of them have been used as traditional medicines by people from many countries around the world. Since these natural compounds often contain a broad spectrum of biological activities, they have the potential to be further purified and developed into drugs for the treatment of many human disease conditions. Cordyceps (ascomycete) fungus has been used in Chinese medicine since ancient times [
11]. The pharmacological properties of Cordyceps extract have been studied in both infectious and noninfectious diseases [
12]. Cordycepin (3′-deoxyadenosine) is an adenosine derivative, and a major bioactive compound in
Cordyceps sinensis and
Cordyceps militaris. This compound was previously reported to have antiviral activity [
13,
14,
15,
16,
17]. However, the anti-DENV activity of cordycepin is unknown. Since cordycepin is an adenosine derivative, we hypothesized that cordycepin has anti-DENV activity and is likely to inhibit DENV replication. The results of this study clearly demonstrate that cordycepin inhibits DENV replication in African green monkey kidney (Vero) cells and also decreases DENV RNA synthesis. This inhibitory effect was also observed when a
C. militaris extract was similarly tested.
3. Discussion
DENV infection is a major public health problem in many countries worldwide. Its prevalence has also rapidly increased during the past two decades with an approximately 8-fold increase from 505,430 in 2000 to 4.2 million in 2019, as reported by the World Health Organization (WHO) [
1]. A vaccine is needed to prevent and control DENV infection in large populations; however, specific anti-DENV drugs are also needed to interrupt the infection cycle during outbreaks, to reduce disease severity, and to prevent mortality. Thus, the development of these vaccines and anti-DENV drugs is urgently required. To that end, our research group is actively working to identify anti-DENV agents that can be developed and applied for clinical use. Since cordycepin was reported to contain antiviral properties [
13,
14,
15,
16,
17], we hypothesized that cordycepin and
Cordyceps spp. extract possess anti-DENV activity. In this study, we found that cordycepin and
C. militaris extract were able to inhibit DENV infection in DENV-infected Vero cells, and these findings suggest these substances as possible anti-DENV agents.
Cordyceps spp. is primarily and mostly an entomopathogenic fungus, which has been used in traditional Chinese medicine and is well known for its nutraceutical and therapeutic properties [
18]. Cordycepin, which is a major bioactive compound isolated from
Cordyceps spp., has gained interest for its pharmaceutical uses. The pharmaceutical potential of cordycepin has been reported in several disease models. The reported actions of cordycepin include antioxidant, anti-hyperlipidemia, anti-aging, anti-diabetes, anti-cancer, anti-malarial, antifungal, and antiviral. These reported findings emphasize the potential value of cordycepin and
Cordyceps spp. extract for therapeutic applications. Cordycepin (3′-deoxyadenosine) is an adenosine derivative that lacks only a hydroxyl group at the 3′ position of the ribose moiety. Based on its basic structure, cordycepin likely binds to DNA and RNA polymerases, which are crucially required for viral replication. Cordycepin may also be incorporated into DNA and RNA strands to prematurely terminate their synthesis. We, therefore, hypothesized that cordycepin and
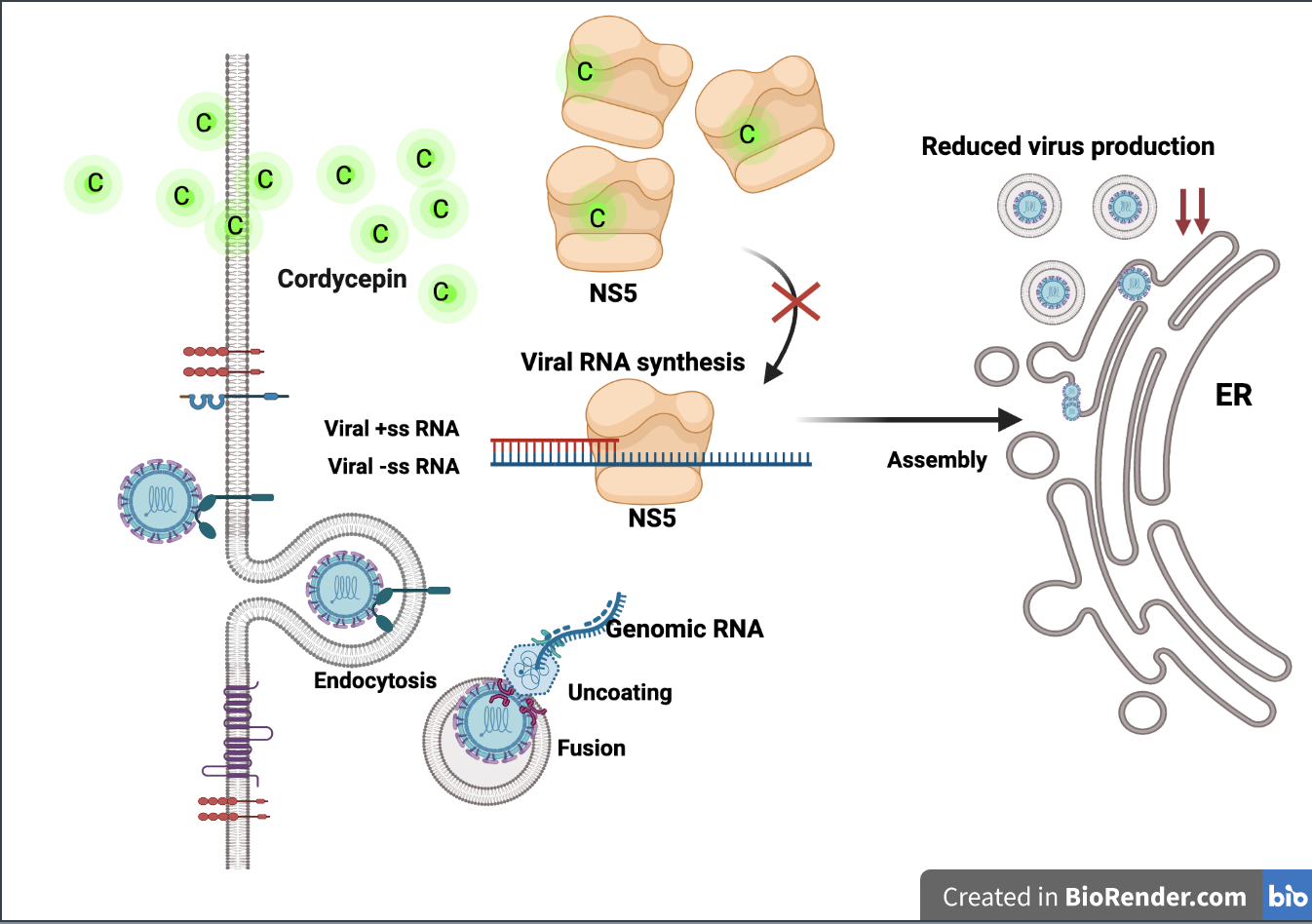
Cordyceps sp. extract are able to inhibit DENV infection and production via inhibition of DENV RNA replication. The results of this study support our hypothesis. Cordycepin could significantly reduce the intracellular DENV envelope (E) protein in a dose-dependent manner, reflecting its ability to inhibit the DENV life cycle (
Figure 1). The life cycle of DENV consists of sequential events beginning with DENV–host receptor binding, internalization, virus disassembly, protein synthesis, RNA synthesis, virus assembly, virus maturation, and ending with virus egress. Inhibition at a single step during the DENV life cycle may have an inhibitory effect on virus production. To single out the step of cordycepin action, we examined the effect of cordycepin in different sets of conditions, including preinfection, coinfection, and postinfection. The preinfection and coinfection conditions reflect the actions of cordycepin on DENV entry at the virus–host binding step and internalization, respectively, whereas the postinfection condition reflects virus disassembly and protein synthesis, and RNA replication (including the later steps of virus assembly and so on). The results of our experiments demonstrated obvious effects of cordycepin at the postinfection step (
Figure 1), which suggests that cordycepin likely acted on RNA synthesis. The effect of cordycepin at the early stage of DENV infection resulted in a reduction of intracellular E protein levels and virus production in cell culture supernatants (
Figure 2). Notably, the effect of cordycepin was absent in either preinfection or coinfection condition due to the removal of cordycepin from the system before and after the internalization step, respectively. Accordingly, this short incubation period might not be sufficient to allow the absorption of cordycepin into the cells to inhibit the RNA replication.
Based on our hypothesis that cordycepin binds to DENV RNA-dependent RNA polymerase and inhibits viral RNA synthesis, we further determined the levels of viral RNA in infected Vero cells after cordycepin treatment for 4, 8, and 12 h by the real-time RT-PCR method. In these conditions without cordycepin, the amounts of DENV2 RNA slightly changed 4 and 8 h after infection, but it significantly increased at 12 h (
Figure 3). Interestingly, the treatment of infected cells with cordycepin at concentrations of 50 and 100 μM could clearly block DENV2 RNA synthesis. Since cordycepin is an adenosine derivative, it is likely that it can bind to the DENV NS5 protein (which is a key enzyme in viral RNA synthesis), which comprises 900 amino acids and has the following two functional domains: a methyltransferase (MTase) and an RNA-dependent RNA polymerase (RdRp) at its N-terminus and C-terminus, respectively. The NS5 MTase domain (residues 1–263) functions in capping genomic RNA using S-adenosylmethionine (SAM) as a methyl donor, and it is sequentially methylated at the N-7 position of guanine (m7G) and 2′ OH of ribose of the first strictly conserved adenosine nucleotide (Am) of RNA [
19]. Another NS5 RdRp domain (residues 270–900) is responsible for de novo RNA synthesis without a primer requirement [
19]. To initially examine the possibility that cordycepin can bind to NS5 and inhibit its action, we predicted the interaction of cordycepin and pocket sites on MTase and RdRp, which have been reported as antiviral agent target sites. The molecular docking results revealed interaction between cordycepin and MTase at the pocket that is naturally occupied by
S-adenosyl-l-homocysteine (SAH) [
20], with Δ
G binding of −6.1 kcal/mol (
Figure 4a). The molecular interaction revealed that cordycepin interacted with Gly81, His110, and Glu111 via intermolecular hydrogen bonding, and that competitive binding of cordycepin might disturb the natural ligand to the SAM/SAH binding site. During the process of methylation, MTase converts SAM as the methyl donor into SAH [
21,
22], and this process is an important replication activity. The SAM/SAH binding site on MTase has been proposed as a potential target for anti-flavivirus agents in both DENV [
23] and Zika virus (ZIKV) [
24]. Thus, the binding of cordycepin to the SAM/SAH binding site on MTase may inhibit DENV RNA replication.
Furthermore, the ability of cordycepin to bind to the RdRp domain was also predicted by molecular docking. The result showed that cordycepin could bind to Arg737, Thr794, and TRP795 via hydrogen bonding, and to Arg729, Tyr766, and Ser796 via hydrophobic interactions with Δ
G binding of −5.9 kcal/mol (
Figure 4b). This binding pocket, namely N pocket, was discovered by fragment-based screening via X-ray crystallography targeting the apo-RdRp [
25,
26]. The reported active compound, namely compound 29, which specifically interacted with this pocket, was previously shown to efficiently inhibit RdRp activity, and to reduce DENV replication in BHK21, Huh7, and A549 cells [
27]. Although the Δ
G binding values of cordycepin to these two candidate sites were lower than those of the natural ligands, cordycepin probably bound to multi-target sites that might confer additive or synergistic effects that contribute to its inhibitory activity. Interestingly, the function of MTase was reported to enhance both stability and activity at the initiation and elongation steps of the RdRp domain [
28,
29]. Inhibition of these two domains that function so interdependently may strongly influence viral RNA replication. The antivirus effect of cordycepin against Epstein-Barr virus (EBV) [
13,
14], influenza viruses [
15,
16], and human immunodeficiency virus (HIV) [
17] has been reported. The mechanism of cordycepin was demonstrated to inhibit reverse transcriptase of HIV [
17], which was the direct effect on the viral protein. However, in Epstein–Barr virus, the action of cordycepin was to modulate signal transduction and epigenetic modification, which are related to the regulation of virus infection [
13], highlighting the indirect effect of cordycepin via host protein modulation. Notably, cordycepin has been studied and is known to modulate a variety of cellular signaling pathways involved in apoptosis, proliferation, angiogenesis, and inflammation [
30]. Thus, the indirect effects of cordycepin on the host proteins involved in host antiviral response cannot be excluded. Further in vitro or in vivo studies should focus on both the direct effects of cordycepin on viral proteins, and the indirect effects of cordycepin on host proteins in the inhibition of DENV infection and replication.
The amino acid sequences of the NS5 proteins from the four DENV serotypes are highly conserved. Accordingly, the cordycepin-interacting amino-acid residues on the MTase and RdRp domains are also highly conserved among the four DENV serotypes (
Figure 5a). We, therefore, examined the anti-DENV activities of cordycepin against all four DENV serotypes. The observed cordycepin-induced inhibition of virus production was shown to be dose-dependent in all four DENV serotypes. The highest inhibition of virus production was observed in DENV2, followed by DENV3 (both
p < 0.05). In contrast, the decreases in virus production were non-statistically significant in DENV1 and DENV4 (
Figure 5b). The observed differences in antiviral activity among the four DENV serotypes might be explained by variations in the amino acids lining the binding pocket. Although the key binding amino acid residues were conserved, variation in the other amino acids might affect the accessibility or stabilization of cordycepin inside the pocket. To improve binding affinity, in silico prediction of an additional chemical moiety could be applied, which is based on the size of cordycepin and the area occupied in the pocket. Other derivatives or secondary metabolites of cordycepin might also be selected as alternative molecules for a binding affinity improvement strategy.
Cordyceps spp. extract has been used in traditional medicine and nutraceuticals in Thailand and other Asian countries. Since cordycepin is known to be the major bioactive compound in
Cordyceps spp., we also conducted experiments to evaluate the effects of
C. militaris extract on anti-DENV activity. The amount of cordycepin in
C. militaris extract was examined by HPLC, and the area under the curve (AUC) was calculated. Cordycepin was present at approximately 10.72 mg/g of
C. militaris extract. Treatment with
C. militaris extract at 200 μg/mL (containing about 2 ng/mL or 7.96 μM cordycepin) significantly reduced the intracellular DENV2 E protein in the postinfection condition, which is similar to the reduction observed after cordycepin treatment. Unexpectedly, a reduction in DENV2 E protein was also observed in the coinfection condition, and this finding was not observed after treatment with cordycepin. This suggests an additional effect of
C. militaris extract, which is different from the pure cordycepin compound. Since
C. militaris extract contains a combination of many compounds, the observed inhibitory activity in the coinfection condition might be due to the collective effects of this mixture of compounds. Hence, it is possible that the anti-DENV activity of
C. militaris extract was caused by the combination of at least two or more compounds or agents that inhibited both virus entry and replication. Since DENV infection and its effects on host cells are highly complex and dynamic, the inhibition of DENV infection via different modes of action by targeting multiple steps in the DENV life cycle is a compelling strategy for developing anti-DENV drugs. For example, our group recently reported peptide inhibitors targeting DENV entry by inhibiting virus–host receptor interaction [
31,
32,
33,
34]. A combination of these peptide inhibitors and cordycepin would hypothetically enhance antiviral activity in DENV-infected cells. Further studies should be conducted to test this hypothesis.
Natural compounds from medicinal plants are excellent sources for screening anti-DENV and other antiviral drugs. Additionally, the processes of drug discovery and development can be shortened and accelerated, respectively, due to the already known therapeutic properties and proof-of-safety based on long-standing traditional use. However, the modes of action and molecular mechanisms of the bioactive compounds identified in and isolated from medicinal plants need to be thoroughly investigated before they can be assigned to specific clinical applications.
In conclusion, our study demonstrated the anti-DENV activity of cordycepin and C. militaris extract. Their inhibitory effects take place at the early stage of DENV infection, and they are involved in the step of viral RNA synthesis. The binding of cordycepin to the MTase and RdRp domains of DENV NS5 has been proposed, and is illustrated by molecular docking. The therapeutic potential of cordycepin for DENV infection should be further investigated for future clinical application as an anti-DENV drug.
4. Materials and Methods
4.1. Cell Culture and Virus Propagation
Monkey kidney epithelial Vero cells (CCL8; American Type Culture Collection (ATCC), Manassas, VA, USA) were cultured in minimal essential medium (MEM) supplemented with 10% (v/v) fetal bovine serum (FBS) (Gibco; Thermo Fisher Scientific, Waltham, MA, USA) and penicillin/streptomycin (Gibco, Thermo Fisher Scientific, Waltham, MA, USA). The cells were incubated at 37 °C in a 5% CO2 atmosphere. Four serotypes of DENV, including DENV1 (strain Hawaii originally from Hawaii, USA), DENV2 (strain 16,681 originally from Thailand), DENV3 (strain H87 originally from Thailand), and DENV4 (strain H241 originally from the Philippines), were propagated in C6/36 cells. Cell supernatants containing DENV were collected, and the virus titers were quantitated by focus-forming assay. The virus was stored at −70 °C until use.
4.2. Time of Addition Assay
Cordycepin or
C. militaris extract was tested for its mode of action during the virus life cycle using time-of-addition assay. Briefly, Vero cells were plated in 96-well format (2 × 10
4 cells per well) overnight before the experiment. At the time of experiment, cordycepin or
C. militaris extract at the indicated concentrations was added to the cells in three different conditions, including preinfection, coinfection, and postinfection (
Figure S5). To test the possibility that cordycepin or
C. militaris extract affected on the host cell receptor binding step or the virus entry step, the preinfection or coinfection condition was accordingly performed. The preinfection condition was carried out by adding cordycepin or
C. militaris extract for 30 min and by taking the culture medium out and washing before infection with 1.0 × 10
4 FFU/mL of DENV2 (100 μL), whereas in the coinfection condition, cordycepin or
C. militaris extract and DENV were simultaneously added. After incubation for 2 h to allow the virus binding and internalization, the unbound viruses and cordycepin in the extract were removed. Then, the infected cells were washed once with cold PBS and replaced with a fresh medium. The postinfection condition was performed after the cell–virus incubation for 2 h, addition of cordycepin or
C. militaris extract and its maintenance along the experiment to test the effect of cordycepin or
C. militaris extract on protein or RNA synthesis (including the later steps of virus assembly and so on) after the virus entry step. The infected cells were harvested 48 h after infection to determine the level of intracellular DENV E protein by cell-based enzyme-linked immunosorbent assay (ELISA).
4.3. Virus Titration and Focus Forming Unit (FFU) Staining
To examine the anti-DENV activity of cordycepin or C. militaris extract on virus production, the newly synthesized virus in the culture supernatant was measured by FFU staining. Vero cells were plated in 96-well format (2 × 104 cells per well) overnight before the experiment. The cells were infected with DENV1, DENV2, DENV3, or DENV4 (1.0 × 104 FFU/mL of DENV2, 100 μL). The plate was incubated for 2 h to allow the virus to enter the cells. The unbound virus was removed and a fresh medium containing 25, 50, or 100 μM of cordycepin was added to the cells. The new viral progenies were harvested at 24 or 48 h, and determined for their virus titer by FFU staining.
To determine DENV titers, the Vero cells were plated 2 × 104 cells per well in 96-well format. A 10-fold serial dilution of DENV was added to the cells. After 2 h, the infected cells were overlaid with 2% carboxymethylcellulose medium, and the plate was further incubated for 72 h. The infected cells were fixed and permeabilized using 3.8% formaldehyde and 0.2% Triton X-100, respectively. The 4G2 monoclonal antibody, which is specific to DENV E proteins, was added to the cells and the plate was incubated overnight. The cells were washed three times with phosphate-buffered saline (PBS) containing 0.1% Tween-20 (PBST) before addition of the horseradish peroxidase (HRP)-conjugated secondary antibody (at 1:2000 dilution), which was incubated at room temperature (RT) for 30 min. The DENV-infected cells shown as DENV foci were detected by adding 3,3′-diaminobenzidine (DAB) substrate (Sigma-Aldrich Corporation, St. Louis, MO, USA). The DENV foci were counted manually under a light microscope (20 × magnification) (Nikon Instruments, Inc., Melville, NY, USA).
4.4. Cell-Based Enzyme-Linked Immunosorbent Assay (ELISA)
Infected cells from the time-of-addition assay were examined for their level of intracellular DENV E protein. The infected cells were collected at the indicated time and washed once with PBS. The cells were fixed and permeabilized using 3.8% formaldehyde and 0.1% Triton X-100, respectively, before addition of the 4G2 monoclonal antibody specific to the DENV E protein. The plate was incubated overnight and washed three times with PBST. HRP-conjugated secondary antibody (dilution 1:2000) was then added to the wells and incubated at RT for 30 min. The plate was washed three times with PBST before adding the 3,3′,5,5′-tetramethylbenzidine substrate (Invitrogen Corporation, Carlsbad, CA, USA). Absorbance was detected and calculated for the percentage of DENV E protein. The absorbance percentage for infected cells without treatment (control) was set as 100%.
4.5. Immunofluorescence Assay (IFA)
The number of DENV-infected cells was detected by IFA. Briefly, the cells were plated in 24-well format (1.0 × 105 cells per well) overnight before the experiment. On the day of the experiment, the cells were infected with 1.0 × 104 FFU/mL of DENV2 (500 μL). The plate was incubated with the viruses for 2 h to allow virus entry into the cells. The unbound viruses were removed and replaced with a fresh medium containing cordycepin at concentrations of 25, 50, and 100 μM; 48 h after infection, the infected cells were harvested before being fixed and permeabilized using 3.8% formaldehyde and 0.1% Triton X-100, respectively. After that, a 4G2 monoclonal antibody specific to DENV E protein was added. The cells were washed three times with PBST, and Alexa Fluor 488 goat anti-mouse IgG (Invitrogen) was added in combination with Hoechst® 33,342 nucleic acid stain (Invitrogen). DENV-infected cells were monitored under a fluorescence microscope.
4.6. Real-Time Polymerase Chain Reaction (RT-PCR)
To investigate the effect of cordycepin on DENV replication, Vero cells were seeded at 1.5 × 105 cells per well in a 12-well plate for 24 h before addition of DENV2 for 2 h at 37 °C with 5% CO2. The unbound viruses were removed and cordycepin at concentrations of 50 and 100 µM was added. The cells were harvested at 4, 8, and 12 h, and the total RNA was extracted using TRIzol™ Reagent (Invitrogen). The RNA (1.0 µg) was converted into cDNA using a cDNA Synthesis Kit (Toyobo Life Science, Osaka, Japan). The PCR mixture contained 0.5 µg of cDNA, DENV E-specific primers [10 µM of D2R (5′-CCGGCTCTACTCCTATGATG-3′), and 10 µM of D2L (5′-ATCCAGATGTCATCAGGAAAC-3′)], and 2 × SensiFAST SYBR No-ROX Mix (Meridian Bioscience, Cincinnati, OH, USA). RT-PCR was then performed on an iCycler Thermal Cycler (Bio-Rad Laboratories, Hercules, CA, USA). The data were normalized with the housekeeping gene GAPDH and analyzed for relative value to that of non-treated control (set as 1).
4.7. Molecular Docking
Molecular docking aimed to predict the interaction between cordycepin and the structures of DENV NS5 MTase and RdRp. In this study, the three-dimensional structure of cordycepin was generated from its crystal structure (CCDC number 129360) in MOL2 format. Hydrogens were added, non-polar hydrogens were merged, and the missing atoms were added and converted from MOL2 to pdbqt format using AutoDock Tools version 1.5.6 [
35].
The crystallographic coordinates of DENV NS5 MTase and RdRp were retrieved from the Protein Data Bank (PDB ID: 1R6A, 5I3P, 5I3Q, and 5K5M). The water molecules, ions, or inhibitors in each structure were removed to prepare a receptor target model in docking simulations. Grid box sizes of 30 × 40 × 40 and 20 × 20 × 24 Å at a spacing of 1 Å were generated by defining the ligand binding site (SAM binding site, co-crystalized ligand or equivalent positions in the different protein sequences as centroid). The docking procedure was performed using Autodock Vina with rigid docking. A complex structure with favorable binding affinity was selected, and molecular interactions of the ligand and the protein were analyzed using LigPlot version 2.2.4 [
36]. The docking poses were prepared using the PyMOL Molecular Graphics System, Version 2.0 (Schrödinger, LLC.; New York, NY, USA).
4.8. Cordyceps Extraction
Dried fruiting bodies that included substrates of C. militaris were obtained from AAVA Group Co., Ltd., Bangkok, Thailand. The sample was ground and soaked with sterile distilled water at a ratio of 1:10 (w/v). The sample was macerated with distilled water at 45 °C for 3 h. The extract was filtered using Whatman No. 1 filter paper and then evaporated using a rotary evaporator at 45 °C under a reduced pressure of 50 mbar. The extract was dried by lyophilization and stored at −20 °C until use.
4.9. High Performance Liquid Chromatography (HPLC)
Crude extract of C. militaris was analyzed by high-performance liquid chromatography (HPLC). A pure compound of cordycepin (Sigma-Aldrich) was purchased to optimize the detection of the cordycepin component in C. militaris extract using an isocratic HPLC system with a conventional C18 column. The extract was filtered through a 0.45 µm microfilter, and 50 µL of the filtrate was injected into the HPLC system (Agilent Technologies, Santa Clara, CA, USA). A ZORBAX Eclipse XDB-C18 column (4.6 × 150 mm, 5 µm; Agilent Technologies) employed an ultraviolet (UV) photodiode array detector (254 nm). The HPLC system was controlled to have a flow rate of 1.0 mL per minute, and a running time of 30 min at 31 °C. Cordycepin in C. militaris extract was separated by water and methanol in the ratio of 92:8 (v/v). The bioactive compound profiles of C. militaris extract were analyzed by comparing them with those of standard cordycepin.
4.10. Statistical Analysis
The datasets were analyzed, and the results are shown as the mean ± SEM of at least three independent experiments. The statistical analysis was performed using a Student’s t-test in GraphPad Prism software, version 9.0.2 (GraphPad Software, Inc., San Diego, CA, USA). The symbols *, **, ***, **** indicate p-values of <0.05, <0.01, <0.001, and <0.0001, respectively.


 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





