3.1. Bogdanov–Takens Bifurcation in Conductance-Based Models
We are looking for a critical point that encloses most of the diverse, but planar (two variables) dynamics observed in single neurons. In electrophysiological experiments, when the transmembrane current
I is increased, neuron dynamics abruptly transit from a fixed point (rest potential) to a limit cycle (spiking). This transition to relaxation oscillations is the landmark of single neuron dynamics. Often, different classes of neurons have different types of spiking dynamics. The rest potential can exhibit an exponential decay after being perturbed (node) or damped oscillations (focus). In addition, the transition to spiking can occur via many dynamical mechanisms as saddle-node homoclinic bifurcation, bistability between a focus and limit cycle, subcritical Andronov–Hopf bifurcation between others. These dynamical mechanisms are seldom controlled by the experimenter, but they depend on the specific class of neuron, that is its specific set of ionic conductances and the values of its parameters. When the current
I is increased, depending on the type of neuron, the local bifurcations observed are usually saddle-node or Andronov–Hopf. Then, the first condition that we will impose for a critical point that encloses the diversity observed in single neurons dynamics is to have both local saddle-node and Andronov–Hopf bifurcation in its neighborhood. This must be a bifurcation point of at least codimension two, since we need one parameter (usually
I) to reach the bifurcation and another to transit from saddle-node to Andronov–Hopf (or
vice versa). We exclude the bifurcation arising from the eigenvalues
,
i.e., a simultaneous saddle-node and Andronov–Hopf bifurcation, since it leads to a three-variable dynamics. A low codimension candidate for this is a double zero bifurcation (two eigenvalues equal to zero). Slight departures in the parameter space from this point will transform this double zero in a saddle-node or an Andronov–Hopf bifurcation locally. Using the characteristic polynomial (Equation (
15)), we can find analytically the conditions to have double zero eigenvalues:
Since the relaxation times of the gating variables (
) are always positive quantities, then these two conditions are equivalent to:
Using Expressions (
6) and (
7), we obtain that the previous expression is equivalent to:
At this point, it is necessary to define the following quantity:
The function
can be interpreted as the relaxation time of the ohmic part of the I-V curve. The function
, called the gating function, will quantify the linear contribution of a given gating variable
j to the overall CB model dynamics. The sign of
will determine if a gating variable is an amplifier (
i.e., contributes to the dynamics as a positive feedback, is negative) or resonant (
i.e., contribute to the dynamics as negative feedback, is positive) according to the classification used in computational neuroscience [
7]. This naturally leads us to a sign criteria to designate an amplifier or a resonant gating variable. Using these definitions, we can write the double zero condition in terms of the gating functions and relaxation times:
Since the relaxation times (
and
) are positive quantities, then after some simple algebra, we can conclude that the necessary conditions for having a double zero bifurcation are: (1) at least one amplifier and one resonant gating variable (
i.e., at least one
must have a different sign); (2) at least one gating variables relaxation time is different. In other words, a conductance-based model must have gating variables that produce both positive and negative feedback, and there must be a time scale separation between at least one of them. Furthermore, we showed that if the system has two time scales, the fastest gating variables must be an amplifier and the slower resonant (
Supplementary Materials, Section 2). This result is surprising, since the conditions mentioned have been reported as a key dynamical mechanism for neurons to exhibit excitable behavior [
15,
16]. In addition, we showed that in CB models, a double zero bifurcation is always a BT bifurcation (
Supplementary Materials, Section 1.1). Additionally, we calculated the complete Jordan base (
Supplementary Materials, Section 1.2) and showed that generically, a Jordan block will arise. An example of the BT bifurcation in the widely-known Hodgkin and Huxley CB model [
15] is shown in
Figure 1B.
3.2. Additional Condition to the Bogdanov–Takens Bifurcation
We already identified BT as a critical point of interest. However, there is an important feature that is commonly observed in neurons and neuron models that was not included in the later discussion. It has been observed in experiments and simulations that under realistic parameter values, the I-V curve is usually either monotonically increasing or has two changes in its concavity (see
Figure 1A). As we discussed in
Section 2, the zeros of the I-V curve determine the fixed points of the dynamics. Then, if the I-V curve is monotonically increasing, the dynamics will have just one fixed point, and if its concavity changes just twice, it has three. Nevertheless, the stability of the fixed point depends on the eigenvalues of the linear matrix, which can be calculated using the expression for the characteristic polynomial in Equation (
15), and they do not depend explicitly on the second derivative of the I-V curve. Since we want to include both scenarios (one fixed point and three fixed points) in the neighborhood of the critical point, then we need to impose that:
In addition to the two BT conditions, this third condition puts the critical point in the transition point between one to three fixed points. Then, in its neighborhood, one finds many of the local bifurcation scenarios previously observed in neuron models: Andronov–Hopf bifurcation with one fixed point, Andronov–Hopf bifurcation with three fixed points and a saddle-node with three fixed points. When the BT normal form is calculated in this point, we obtain surprising results regarding its universality to reproduce single neuron dynamics.
Figure 1.
(A) The I-V curve (i.e., function ) for the Hodgkin and Huxley model for three different values of the potassium conductance . A red dot indicates a fixed point in the dynamics. (B) The Bogdanov–Takens (BT) bifurcation in the Hodgkin and Huxley model. The function is shown in purple. The function is shown in green. The plots in black frames show the eigenvalues of the linear matrix in the complex plane. We omitted the greater in absolute value eigenvalue (far to the left) in order to better visualize the two critical ones. Tags 1–3 indicate the corresponding values of the variable . The plot in a red dashed frame shows the eigenvalues of the linear matrix in the complex plane when the model undergoes a BT bifurcation.
Figure 1.
(A) The I-V curve (i.e., function ) for the Hodgkin and Huxley model for three different values of the potassium conductance . A red dot indicates a fixed point in the dynamics. (B) The Bogdanov–Takens (BT) bifurcation in the Hodgkin and Huxley model. The function is shown in purple. The function is shown in green. The plots in black frames show the eigenvalues of the linear matrix in the complex plane. We omitted the greater in absolute value eigenvalue (far to the left) in order to better visualize the two critical ones. Tags 1–3 indicate the corresponding values of the variable . The plot in a red dashed frame shows the eigenvalues of the linear matrix in the complex plane when the model undergoes a BT bifurcation.
3.3. The Cubic Bogdanov–Takens Normal Form
Using the results in Elphick, Tirapegui, Brachet, Coullet and Iooss in [
2], it is possible to characterize and calculate explicitly all of the non-linear terms for the BT normal form. In the case of the BT bifurcation, the equations obtained can be satisfied in more than one way, leaving an inherent freedom to incorporate the normal from (see Section 3,
Supplementary Materials). We have two extreme choices: Arnold’s or the Takens choice [
22]. Since in Arnold’s choice, we gain a Hamiltonian intuition, we make this choice here and use the Arnold form of the cubic BT normal form, which can be written as:
We will call the function the force and the function the friction using the obvious analogy to mechanics. More explicitly, , and . The unfolding coefficients are α, β and . When they vanish, we are at the critical point, and when they take small values, we are in the neighborhood of the critical point, where we expect the normal form to be valid quantitatively. Outside this neighborhood, one expects a qualitative description. We have three unfolding parameters, since the codimension is three. We can define a potential , such that the force is given by , where . Then, we can define the “energy” of the BT normal form as and conclude that . This last equation shows that the BT normal form does not conserve the energy. The reader should notice that the BT normal form can undergo an injection or dissipation of energy depending on the sign of , and this sign can of course change when u changes.
The most common scaling studied in the literature is quadratic,
i.e.,
and
. The complete bifurcation diagram is known, and it exhibits saddle-node, Andronov–Hopf and homoclinic bifurcations [
22,
23]. However, the dynamics observed in this normal form is not consistent with the dynamics observed in single neurons. First, it is not bounded in the complete phase space, which means that for some regions in the phase space,
u can go to infinity (see
Figure 2 left plot). Second, it does not display many of the distinctive global bifurcations observed in neurons as the saddle-node homoclinic bifurcation or the saddle homoclinic bifurcation.
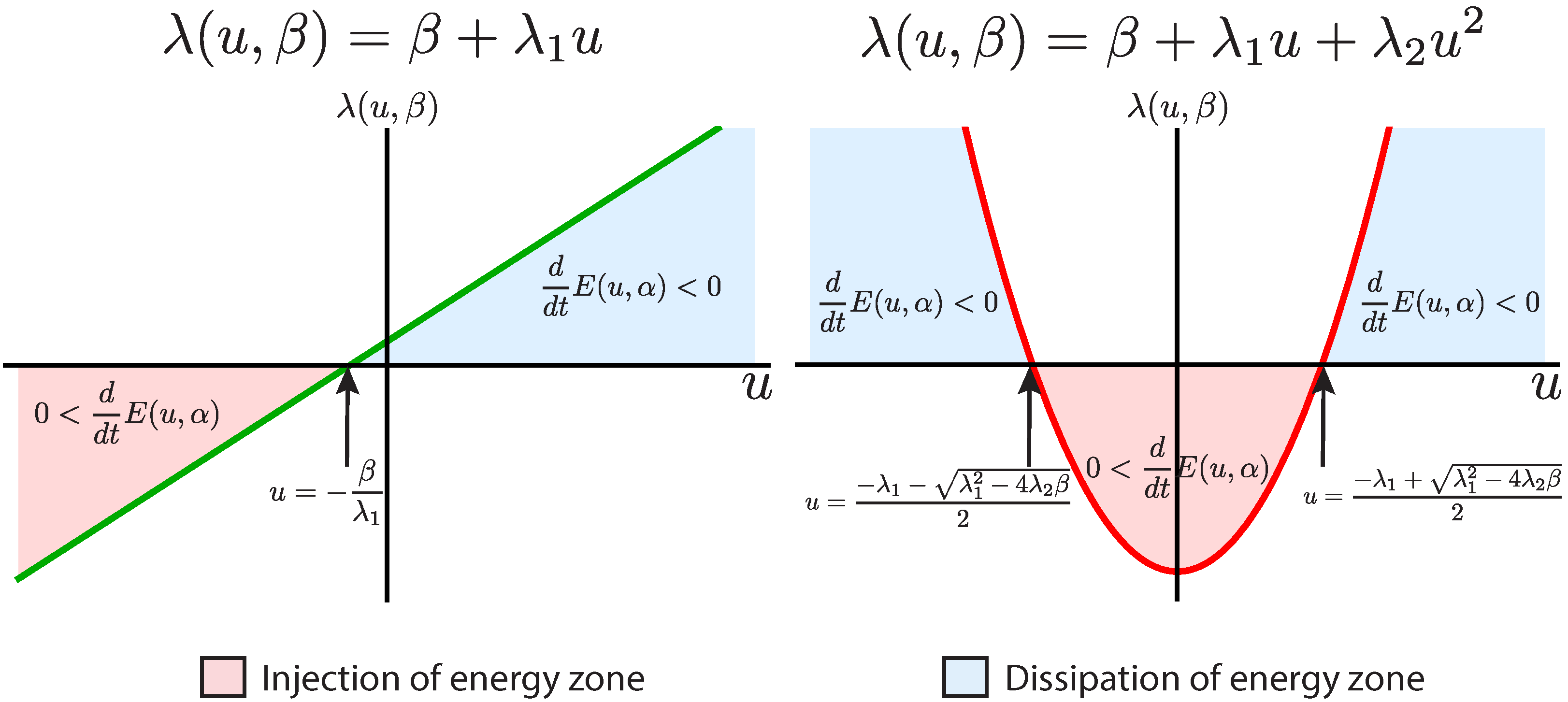
Figure 2.
The left graphic shows in green a linear friction curve. The right graphic shows in red a quadratic friction curve with . The arrow indicates the points where the friction changes the sign in both cases. In pale red, we indicate the region where the system undergoes an injection of energy and in pale blue the region where the system undergoes a dissipation of energy .
Figure 2.
The left graphic shows in green a linear friction curve. The right graphic shows in red a quadratic friction curve with . The arrow indicates the points where the friction changes the sign in both cases. In pale red, we indicate the region where the system undergoes an injection of energy and in pale blue the region where the system undergoes a dissipation of energy .
Since we know analytically the Jordan basis, we are able to calculate explicitly the cubic BT normal form using the methods developed in [
2]; that is, to calculate for any model of the form of Equation (
1), the explicit dependence of the unfolding parameters (
α,
β,
) and the coefficients (
,
,
) on the parameters of the model. We find that the critical variables of the normal form can be chosen to be
, where
u is the dimensionless membrane potential and
its time derivative (see Equation (1), Section 1.1,
Supplementary Materials). The calculations, which are standard after the results in [
2], are presented for the quadratic coefficient
(see Section 3,
Supplementary Materials). Very surprisingly we found that the quadratic term of the force is proportional to the second derivative of the I-V curve,
i.e.:
This relation is very important, since it shows that in order to have one or three fixed points (which is what is observed), then in the unfolding of the normal form, we have to impose an additional condition, namely that the second derivative of the I-V function with respect to
u vanishes. This means that in the normal form, the calculated
will be very small (see Equation (
18)). Therefore, we have to look for another scaling. The next scaling to consider is a quadratic friction,
i.e.,
. Since we are looking for a bounded dynamics, we must consider
. As is shown in
Figure 2 (right plot), for some values of
u,
i.e.,
, there will be only the injection of energy, and it can be shown analytically that the system cannot have a stable fixed point in this region, no matter the form of the force. Therefore, all of the orbits will escape from this region, but the orbits will not go to infinity, because as soon as they reach the regions
or
, the system will dissipate energy quadratically with the variable
u and the dynamics will be bounded to a certain region of phase space. Then this scaling will present a bounded dynamics for sensible force functions. Moreover, it has been shown that the BT normal form with a quadratic friction and cubic force presents a rich set of global bifurcations, some of which are characteristic of single neuron dynamics [
24]. The cubic BT normal form has the following form:
This equation corresponds to the normal form of a particular BT bifurcation point of codimension three described by Equations (
16)–(
18). We expect that this normal form displays at least some of the fundamental aspects of single neuron dynamics, since we construct this critical point imposing constrains widely observed among neurons. However, it is not clear if this equation is able to reproduce the diversity in the observed dynamics of neurons. We show that it does in the next section.


{kind=link}
{kind=link}
{kind=link}
{kind=link}



