K-th Nearest Neighbor (KNN) Entropy Estimates of Complexity and Integration from Ongoing and Stimulus-Evoked Electroencephalographic (EEG) Recordings of the Human Brain

Abstract
1. Introduction
1.1. Information-Theoretic Measures of Brain Integration and Complexity
1.2. Ongoing Versus Stimulus-Elicited EEG Integration and Complexity
1.3. K-th Nearest Neighbor Entropy Estimation for EEG Integration and Complexity
1.4. The Present Study
2. Materials and Methods
2.1. Participants
2.2. Resting State EEG
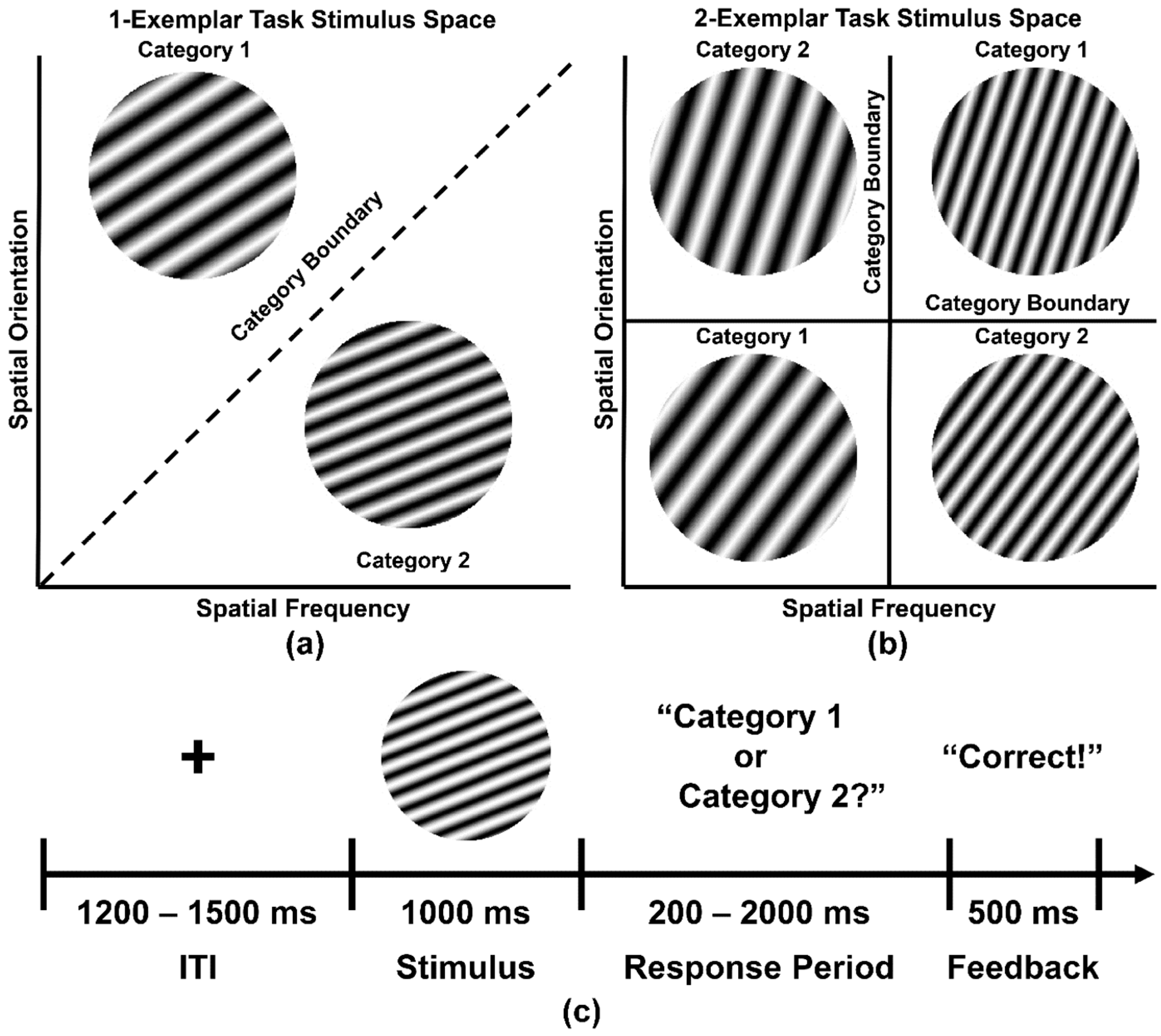
2.3. Categorization Task
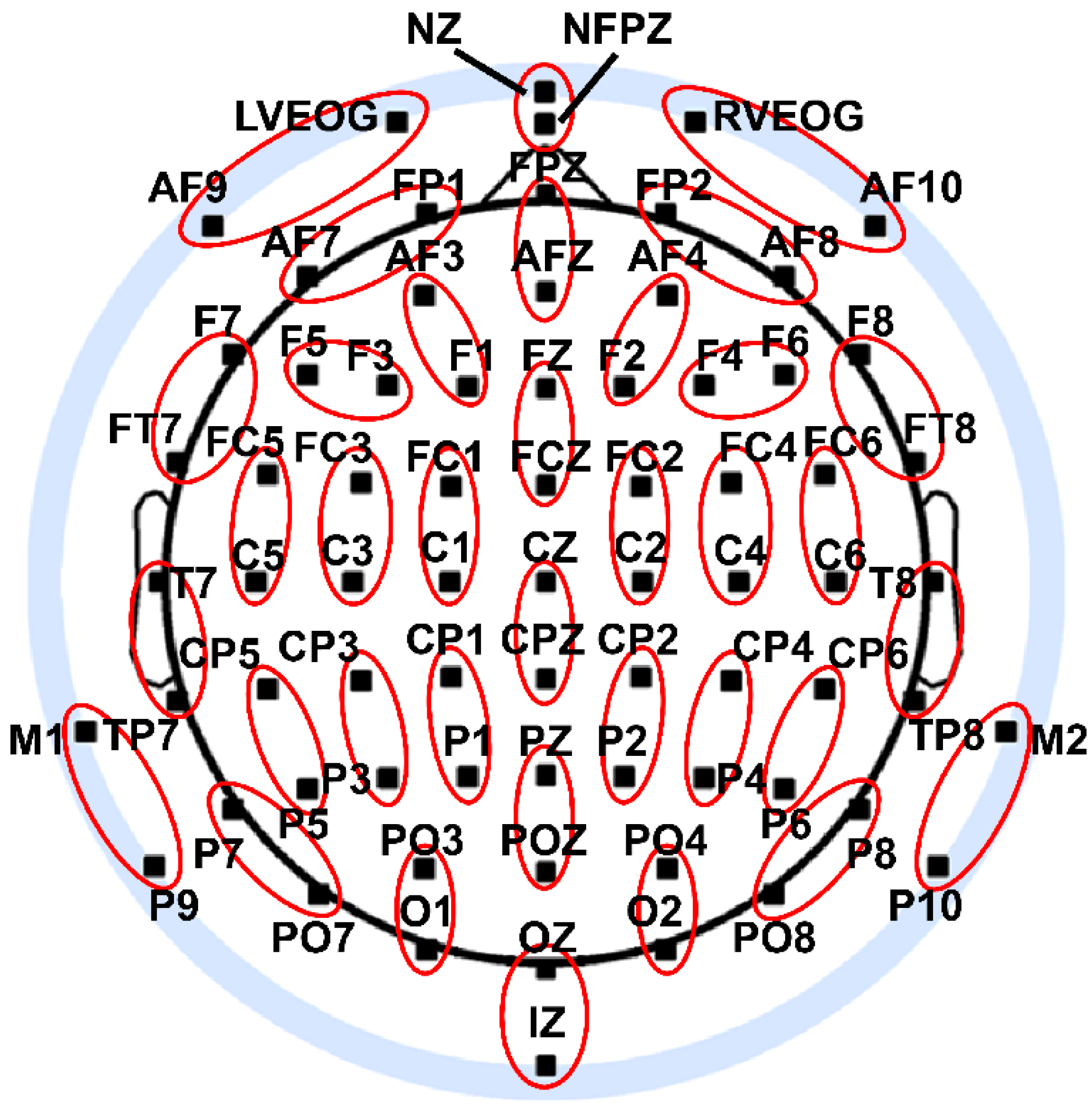
2.4. EEG Recording and Preprocessing
2.5. Estimation of Induced and Evoked Categorization Task EEG Signals
2.6. Computation of EEG Integration and Complexity
2.7. Computation of EEG Power and Phase Synchronization
2.7.1. EEG Power
2.7.2. EEG Synchronization
2.8. Statistical Analysis of Observed EEG Measures
2.8.1. Parametric Statistics
2.8.2. Surrogate Statistics
2.9. Dipole Modeling of Categorization Task EEG Data
3. Results
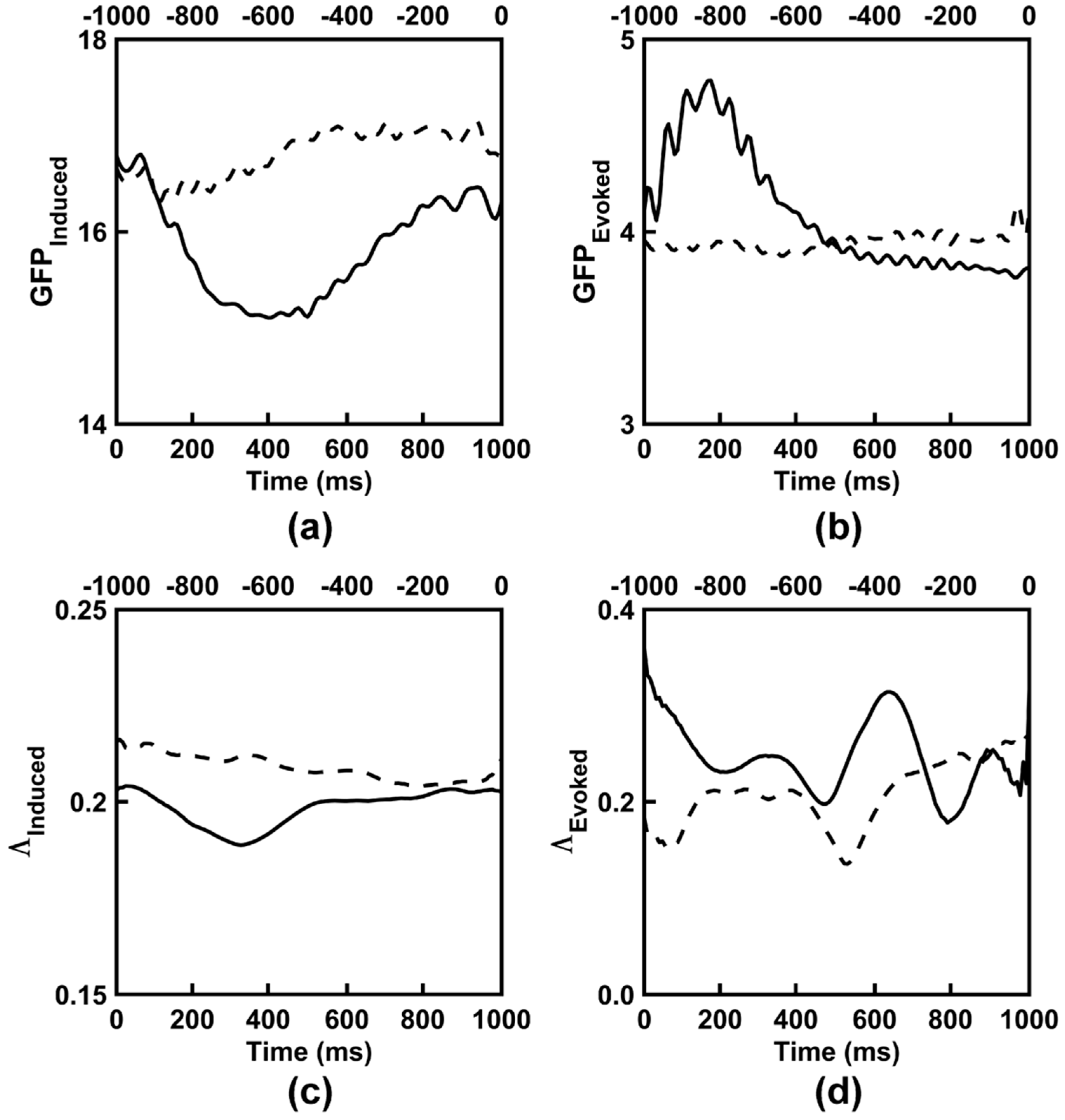
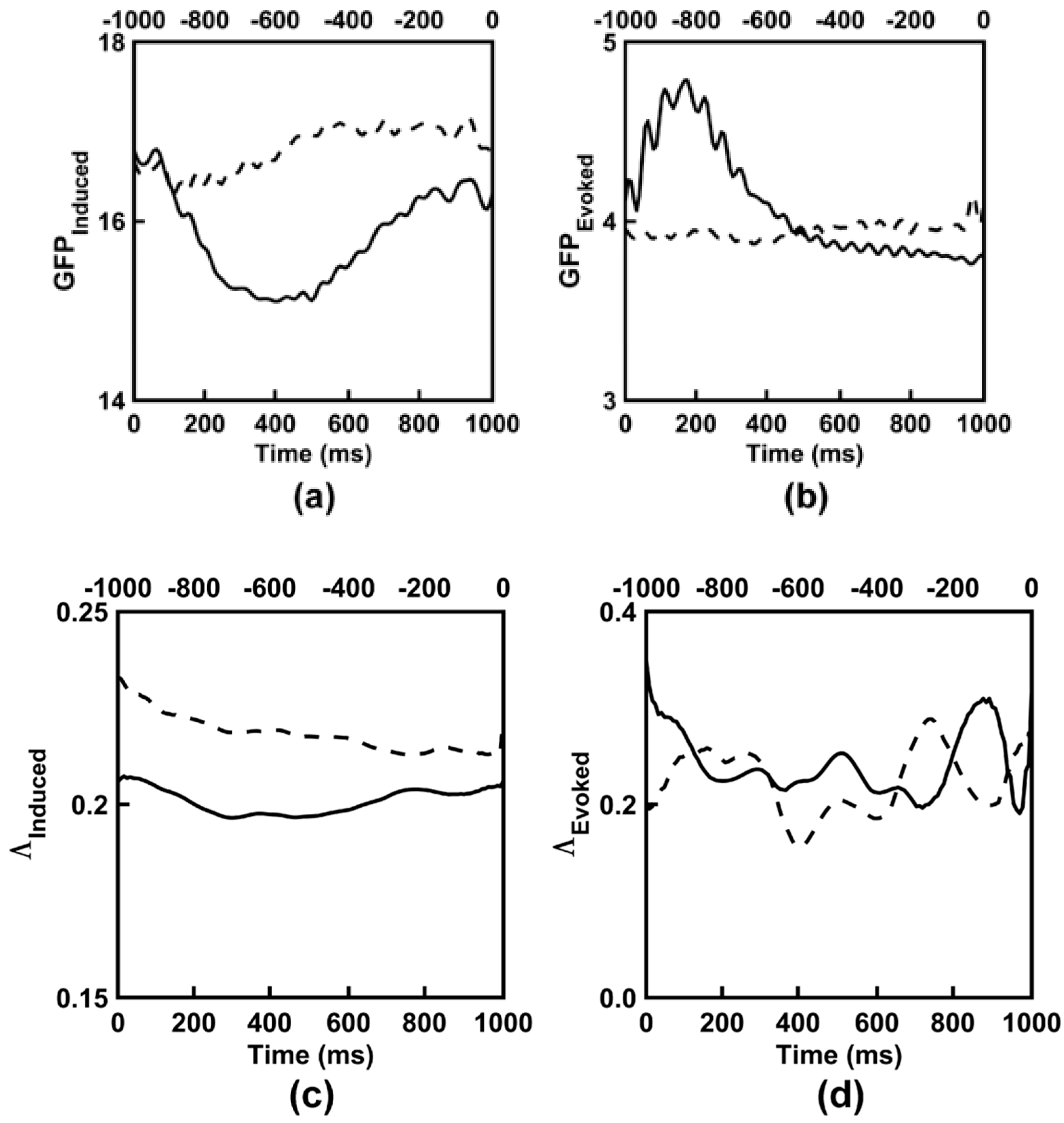
3.1. Observed Induced EEG Activity Integration and Complexity
3.2. Observed Evoked EEG Activity Integration and Complexity
3.3. Observed EEG Power and Synchronization
3.4. Categorization Task Behavior
3.5. Dipole Simulation of Categorization Task EEG
3.6. Comparison of KNN-based and Gaussian-Based EEG Integration and Complexity Estimation
4. Discussion
4.1. Stimulus-Induced and Stimulus-Evoked Changes in Scalp EEG Integration and Complexity
4.2. Resting State Scalp EEG Integration and Complexity
4.3. Advantages and Disadvantages of KNN Metrics for EEG Entropy Estimation
4.4. Study Limitations
5. Conclusions
Supplementary Materials
Funding
Conflicts of Interest
References
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Fair, D.A.; Cohen, A.L.; Power, J.D.; Dosenbach, N.U.F.; Church, J.A.; Miezin, F.M.; Schlaggar, B.L.; Petersen, S.E. Functional brain networks develop from a “local to distributed” organization. PLoS Comput. Biol. 2009, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Tononi, G.; Edelman, G.M.; Sporns, O. Complexity and coherency: Integrating information in the brain. Trends Cogn. Sci. 1998, 2, 474–484. [Google Scholar] [CrossRef]
- Tononi, G.; McIntosh, A.R.; Russell, D.P.; Edelman, G.M. Functional clustering: Identifying strongly interactive brain regions in neuroimaging data. Neuroimage 1998, 7, 133–149. [Google Scholar] [CrossRef] [PubMed]
- Tononi, G.; Sporns, O.; Edelman, G.M. A measure for brain complexity: Relating functional segregation and integration in the nervous system. Proc. Natl. Acad. Sci. USA 1994, 91, 5033–5037. [Google Scholar] [CrossRef] [PubMed]
- Tononi, G.; Sporns, O.; Edelman, G.M. A complexity measure for selective matching of signals by the brain. Proc. Natl. Acad. Sci. USA 1996, 93, 3422–3427. [Google Scholar] [CrossRef] [PubMed]
- Bastos, A.M.; Schoffelen, J.-M. A tutorial review of functional connectivity analysis methods and their interpretational pitfalls. Front. Syst. Neurosci. 2016, 9, 175. [Google Scholar] [CrossRef]
- Rapp, P.E.; Cellucci, C.J.; Watanabe, T.A.A.; Albano, A.M. Quantitative characterization of the complexity of multichannel human EEGs. Int. J. Bifurcat. Chaos 2005, 15, 1737–1744. [Google Scholar] [CrossRef]
- Zhai, Y.; Kiss, I.Z.; Daido, H.; Hudson, J.L. Extracting order parameters from global measurements with application to coupled electrochemical oscillators. Physical D 2005, 205, 57–69. [Google Scholar] [CrossRef]
- Trujillo, L.T.; Stanfield, C.T.; Vela, R.D. The effect of electroencephalogram (EEG) reference choice on information-theoretic measures of the complexity and integration of EEG signals. Front. Neurosci. 2017, 11, 425. [Google Scholar] [CrossRef] [PubMed]
- Burgess, A.P.; Rehman, J.; Williams, J.D. Changes in neural complexity during the perception of 3D images using random dot stereograms. Int. J. Psychophysiol. 2003, 48, 35–42. [Google Scholar] [CrossRef]
- Gu, F.; Meng, X.; Shen, E.; Cai, A. Can we measure consciousness with EEG compexities? Int. J. Bifurcat. Chaos 2003, 13, 733–742. [Google Scholar] [CrossRef]
- Jin, S.-H.; Kim, S.Y.; Park, K.H.; Lee, K.-J. Differences in EEG between gifted and average students: Neural complexity and functional cluster analysis. Int. J. Neurosci. 2007, 117, 1167–1184. [Google Scholar] [CrossRef] [PubMed]
- Papadelis, C.; Kourtidou-Papadeli, C.; Bamidis, P.D.; Maglaveras, N.; Pappas, K. The effect of hypobaric hypoxia on multichannel EEG signal complexity. Clin. Neurophysiol. 2007, 118, 31–52. [Google Scholar] [CrossRef] [PubMed]
- Van Cappellen van Walsum, A.-M.; Pijnenburg, Y.A.L.; Berendse, H.W.; van Dijk, B.W.; Knol, D.L.; Scheltens, P.; Stam, C.J. A neural complexity measure applied to MEG data in Alzheimer’s disease. Clin. Neurophysiol. 2003, 114, 1034–1040. [Google Scholar] [CrossRef]
- Van Putten, M.J.A.M.; Stam, C.J. Application of a neural complexity measure to multichannel EEG. Phys. Lett. A 2001, 281, 131–141. [Google Scholar] [CrossRef]
- Branston, N.M.; El-Deredy, W.; McGlone, F.P. Changes in neural complexity of the EEG during a visual oddball task. Clin. Neurophysiol. 2005, 116, 151–159. [Google Scholar] [CrossRef]
- Hermann, C.S.; Grigutsch, M.; Busch, N.A. EEG oscillations and wavelet analysis. In Event-Related Potentials: A Methods Handbook; Handy, T.C., Ed.; MIT Press: Cambridge, MA, USA, 2005; pp. 229–260. ISBN 0-262-08333-7. [Google Scholar]
- Klimesch, W.; Russegger, H.; Doppelmayr, M.; Pachinger, T. A method for the calculation of induced band power: Implications for the significance of brain oscillations. Electroencephalogr. Clin. Neurophysiol. 1998, 108, 123–130. [Google Scholar] [CrossRef]
- Luck, S.J. An Introduction to the Event-Related Potential Technique; MIT Press: Cambridge, MA, USA, 2005; ISBN 0-262-62196-7. [Google Scholar]
- Kipiński, L.; König, R.; Sielużycki, C.; Kordecki, W. Application of modern tests for stationarity to single-trial MEG data. Transferring powerful statistical tools to neuroscience. Biol. Cybern. 2011, 105, 183–195. [Google Scholar] [CrossRef]
- Elul, R. Gaussian behavior of the electroencephalogram: Changes during performance of mental task. Science 1969, 164, 328–331. [Google Scholar] [CrossRef]
- Xiong, W.; Faes, L.; Ivanov, P.C. Entropy measures, entropy estimators, and their performance in quantifying complex dynamics: Effects of artifacts, nonstationarity, and long-range correlations. Phys. Rev. E 2017, 95, 062114. [Google Scholar] [CrossRef]
- Wollstadt, P.; Martínez-Zarzuela, M.; Vicente, R.; Díaz-Pernas, F.J.; Wibral, M. Efficient transfer entropy analysis of non-stationary neural time series. PLoS ONE 2014, 9, e102833. [Google Scholar] [CrossRef] [PubMed]
- Charsyńska, A.; Gambin, A. Improvement of the k-NN entropy estimator with applications in systems biology. Entropy 2015, 18, 13. [Google Scholar] [CrossRef]
- Kozachenko, L.; Leonenko, N. Sample Estimate of the Entropy of a Random Vector. Probl. Inf. Transm. 1987, 23, 95–101. [Google Scholar]
- Kraskov, A.; Stögbauer, H.; Grassberger, P. Estimating mutual information. Phys. Rev. E 2004, 69, 066138. [Google Scholar] [CrossRef] [PubMed]
- Lord, W.M.; Sun, J.; Bollt, E.M. Geometric k-nearest neighbor estimation of entropy and mutual information. Chaos 2018, 28, 033114. [Google Scholar] [CrossRef]
- Singh, H.; Misra, N.; Hnizdo, V.; Fedorodicz, A.; Demchuk, E. Nearest neighbor estimates of entropy. Am. J. Math. Mangag. Sci. 2003, 23, 301–321. [Google Scholar] [CrossRef]
- Sanguinetti, J.L.; Trujillo, L.T.; Schnyer, D.M.; Allen, J.J.B.; Peterson, M.A. Increased alpha band activity indexes inhibitory competition acorss a border during figure assignment. Vis. Res. 2016, 126, 120–130. [Google Scholar] [CrossRef]
- Klimesch, W.; Doppelmayr, M.; Röhm, D.; Pöllhuber, D.; Stadler, W. Simultaneous desynchronization and synchronization of different alpha responses in the human electroencephalogram: A neglected paradox? Neuroci. Lett. 2000, 284, 97–100. [Google Scholar] [CrossRef]
- Trujillo, L.T.; Allen, J.J.B. Theta EEG dynamics of the error-related negativity. Clin. Neurophysiol. 2007, 118, 645–668. [Google Scholar] [CrossRef] [PubMed]
- Cuffin, B.N.; Cohen, D. Comparison of the magnetoencephalogram and electroencephalogram. Electroencephalogr. Clin. Neurophysiol. 1979, 47, 132–146. [Google Scholar] [CrossRef]
- Mosher, J.C.; Spencer, M.E.; Leahy, R.M.; Lewis, P.S. Error bounds for EEG and MEG source localization. Electroencephalogr. Clin. Neurophysiol. 1993, 86, 303–321. [Google Scholar] [CrossRef]
- Tenke, C.E.; Kayser, J. Surface Laplacians (SL) and phase properties of EEG rhythms: Simulated generators in a volume-conduction model. Int. J. Psychophysiol. 2015, 97, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Picton, T.W.; Bentin, S.; Berg, P.; Donchin, E.; Hillyard, S.A.; Johnson, R., Jr.; Miller, G.A.; Ritter, W.; Ruchkin, D.S.; Rugg, M.D.; et al. Guidelines for using human event-related potentials to study cognition: Recording standards and publication criteria. Psychophysiology 2000, 37, 127–152. [Google Scholar] [CrossRef] [PubMed]
- Perrin, F.; Pernier, J.; Bertrand, O.; Echallier, J.F. Spherical splines for scalp potential and current density mapping. Electroencephalogr. Clin. Neurophysiol. 1989, 72, 184–187. [Google Scholar] [CrossRef]
- Law, S.K.; Nunez, P.L.; Wijesinghe, R.S. High resolution EEG using spline generated surface laplacians on spherical and ellipsoidal surfaces. IEEE Trans. Biomed. Eng. 1993, 40, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- Gao, S.; Ver Steeg, G.; Galstyan, A. Efficient estimation of mutual information for strongly dependent variables. arXiv, 2014; arXiv:1411.2003. [Google Scholar]
- Khan, S.; Bandyopadhyay, S.; Ganguly, A.R.; Saigal, S.; Erickson, D.J.; Protopopescu, V.; Ostrouchov, G. Relative performance of mutual information estimation methods for quantifying the dependence among short and noisy data. Phys. Rev. E 2007, 76, 026209. [Google Scholar] [CrossRef]
- Pfurtscheller, G. EEG event-related desynchronization (ERD) and event-related synchronization (ERS). In Electroencephalography: Basic Principles, Clinical Application, and Related Fields, 4th ed.; Niedermeyer, E., Lopes Da Silva, F., Eds.; Williams & Wilkins: Baltimore, MA, USA, 1999; pp. 958–967. ISBN 0-683-30284-1. [Google Scholar]
- Murray, M.M.; Brunet, D.; Michel, C.M. Topographic ERP analyses: A step-by-step tutorial review. Brain Topogr. 2008, 20, 249–264. [Google Scholar] [CrossRef]
- Glass, G.V.; Peckham, P.D.; Sanders, J.R. Consequences of failure to meet assumptions underlying the fixed effects analyses of variance and covariance. Rev. Educ. Res. 1972, 42, 237–288. [Google Scholar] [CrossRef]
- Harwell, M.R.; Rubinstein, E.N.; Hayes, W.S.; Olds, C.C. Summarizing Monte Carlo results in methodological research: The one- and two-factor fixed effects ANOVA cases. J. Educ. Stat. 1992, 17, 315–339. [Google Scholar] [CrossRef]
- Lix, L.M.; Keselman, J.C.; Keselman, H.J. Consequences of assumption violations revisited: A quantitative review of alternatives to the one-way analysis of variance “F” test. Rev. Educ. Res. 1996, 66, 579–619. [Google Scholar] [CrossRef]
- Greenhouse, S.W.; Geisser, S. On methods in the analysis of profile data. Psychometrika 1959, 24, 95–112. [Google Scholar] [CrossRef]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar] [CrossRef]
- Gardiner, J.C.; Luo, Z.; Roman, L.A. Fixed effects, random effects and GEE: What are the differences? Stat. Med. 2009, 28, 221–239. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Mazumdar, M.; Memtsoudis, S.G. Beyond repeated measures ANOVA: Advanced statistical methods for the analysis of longitudinal data in anesthesia research. Reg. Anesth. Pain Med. 2012, 37, 99–105. [Google Scholar] [CrossRef]
- Hubbard, A.E.; Ahern, J.; Fletscher, N.L.; Van der Laan, M.; Lippman, S.A.; Jewell, N.; Bruckner, T.; Satariano, W.A. To GEE or not to GEE: Comparing Population average and mixed models for estimating the associations between neighborhood risk factors and health. Epidemiology 2010, 21, 467–474. [Google Scholar] [CrossRef]
- Liang, K.Y.; Zeger, S.L. Longitudinal data analysis using generalized linear models. Biometrika 1986, 73, 13–22. [Google Scholar] [CrossRef]
- Lachaux, J.-P.; Rodriguez, E.; Le Van Quyen, M.; Lutz, A.; Martinerie, J.; Varela, F.J. Studying single-trials of phase synchronous activity in the brain. Int. J. Bifurcat. Chaos 2000, 10, 2429–2439. [Google Scholar] [CrossRef]
- Lachaux, J.-P.; Rodriguez, E.; Martinerie, J.; Varela, F.J. Measuring phase synchrony in brain signals. Hum. Brain Mapp. 1999, 8, 194–208. [Google Scholar] [CrossRef]
- Nunez, P.L.; Srinivasan, R.; Westdorp, A.F.; Wijesinghe, R.S.; Tucker, D.M.; Silberstein, R.B.; Cadusch, P.J. EEG coherency I: Statistics, reference electrode, volume conduction, Laplacians, cortical imaging, and interpretation at multiple scales. Electroencephalogr. Clin. Neurophysiol. 1997, 103, 499–515. [Google Scholar] [CrossRef]
- Shahbazi, F.; Ewald, A.; Ziehe, A.; Nolte, G. Constructing surrogate data to control for artifacts of volume conduction for functional connectivity measures. In Proceedings of the 17th International Conference on Biomagnetism Advances in Biomagnetism—Biomag 2010 (IFMBE Proceedings), Dubrovnik, Croatia, 28 March–1 April 2010; Supek, S., Sušac, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 28, pp. 207–210, ISBN 978-3-642-12196-8. [Google Scholar]
- Theiler, J.; Eubank, S.; Longtin, A.; Galdrikian, B.; Farmer, J.D. Testing for nonlinearity in time series: The method of surrogate data. Physica D 1992, 58, 77–94. [Google Scholar] [CrossRef]
- Lee, T.-W.; Girolami, M.; Sejnowski, T.J. Independent component analysis using an extended infomax algorithm for mixed sub-gaussian and super-gaussian sources. Neural Comput. 1999, 11, 417–441. [Google Scholar] [CrossRef] [PubMed]
- Efron, B.; Tibshirani, R. Bootstrap methods for standard errors, confidence intervals, and other measures of statistical accuracy. Stat. Sci. 1986, 1, 54–77. [Google Scholar] [CrossRef]
- Jensen, O.; Hesse, C. Estimating distributed representations of evoked responses and oscillatory brain activity. In MEG: An Introduction to Methods; Hansen, P.C., Kringelbach, M.L., Salmelin, R., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 156–185. ISBN 978-0-19-530723-8. [Google Scholar]
- Acebrón, J.A.; Bonilla, L.L.; Vicente, C.J.P.; Ritort, F.; Spigler, R. The Kuramoto model: A simple paradigm for synchronization phenomena. Rev. Mod. Phys. 2005, 77, 137–185. [Google Scholar] [CrossRef]
- Trujillo, L.T. Trujillo (2019) Entropy Journal Article Dataverse; Texas State University: San Marcos, TX, USA, 2019. [Google Scholar]
- Kornguth, S.; Steinberg, R.; Schnyer, D.M.; Trujillo, L.T. Integrating the human into the total system: Degradation of performance under stress. Naval Eng. J. 2013, 125, 85–90. [Google Scholar]
- Witkowski, S.; Trujillo, L.T.; Sherman, S.M.; Carter, P.; Matthews, M.D.; Schnyer, D.M. An examination of the association between chronic sleep restriction and electrocortical arousal in college students. Clin. Neurophysiol. 2015, 126, 549–557. [Google Scholar] [CrossRef]
- Van Albada, S.J.; Robinson, P.A. Transformation of arbitrary distributions to the normal distribution with application to EEG test–retest reliability. J. Neurosci. Methods 2007, 161, 205–211. [Google Scholar] [CrossRef]
- Jarque, C.M.; Bera, A.K. A test for normality of observations and regression residuals. Int. Stat. Rev. 1987, 55, 163–172. [Google Scholar] [CrossRef]
- Royston, J.P. Some techniques for assessing multivarate normality based on the Shapiro-Wilk W. J. R. Stat. Soc. Ser. C Appl. Stat. 1983, 32, 121–133. [Google Scholar] [CrossRef]
- Trujillo-Ortiz, A.; Hernandez-Walls, R.; Barba-Rojo, K.; Cupul-Magana, L. Roystest: Royston’s Multivariate Normality Test. A MATLAB File. 2007. Available online: https://www.researchgate.net/publication/255982178_ROYSTEST_Royston's_Multivariate_Normality_Test (accessed on 11 January 2019).
- Ince, R.A.A.; Giordano, B.L.; Kayser, C.; Rousselet, G.A.; Gross, J.; Schyns, P.G. A statistical framework for neuroimaging data analysis based on mutual information estimated via a gaussian copula. Hum. Brain Mapp. 2017, 38, 1541–1573. [Google Scholar] [CrossRef] [PubMed]
- Norwich, K.H. Information, Sensation, and Perception; Academic Press. Inc.: San Diego, CA, USA, 1993. [Google Scholar]
- Magri, C.; Whittinstall, K.; Singh, V.; Logothetis, N.K.; Panzeri, S. A toolbox for the fast information analysis of multiple-site LFP, EEG and spike train recordings. BMC Neurosci. 2009, 10. [Google Scholar] [CrossRef] [PubMed]
- Misra, N.; Singh, H.; Demchuk, E. Estimation of the entropy of a multivariate normal distribution. J. Multivar. Anal. 2005, 92, 324–342. [Google Scholar] [CrossRef]
- Pola, G.; Schultz, S.R.; Petersen, R.S.; Panzeri, S. A practical guide to information analysis of spike trains. In Neuroscience Databases: A Practical Guide; Kötter, R., Ed.; Springer Science+Business Media: New York, NY, USA, 2003; pp. 139–154. [Google Scholar]
- Pfurtscheller, G. Functional brain imaging based on ERD/ERS. Vis. Res. 2001, 41, 1257–1260. [Google Scholar] [CrossRef]
- Romei, V.; Brodbeck, V.; Michel, C.; Amedi, A.; Pascual-Leone, A.; Thut, G. Spontaneous Fluctuations in Posterior α-Band EEG Activity Reflect Variability in Excitability of Human Visual Areas. Cereb. Cortex 2008, 18, 2010–2018. [Google Scholar] [CrossRef]
- Dumermuth, G. Variance spectra of electroencephalogram in twins. A contribution to the problem of quantification of EEG background activity in childhood. In Clinical Electroencephalography in Childhood; Kellaway, P., Petersén, I., Eds.; Almqvist & Wiksell: Stockholm, Sweden, 1968; pp. 119–154. [Google Scholar]
- Dumermuth, G.; Walz, W.; ScolloLavizzari, G.; Kleiner, B. Spectral analysis of EEG activity during sleep stages in normal adults. Eur. Neurol. 1972, 7, 265–296. [Google Scholar] [CrossRef]
- Pollock, V.E.; Schneider, L.S.; Lyness, S.A. EEG amplitudes in healthy, late-middle-aged and elderly adults: Normality of the distributions and correlations with age. Electroencephalogr. Clin. Neurophysiol. 1990, 75, 276–288. [Google Scholar] [CrossRef]
- Atasoy, S.; Roseman, L.; Kaelen, M.; Kringelbach, M.L.; Deco, G.; Carhart-Harris, R.L. Connectome-harmonic decomposition of human brain activity reveals dynamical repertoire re-organization under LSD. Sci. Rep. 2017, 7, 17661. [Google Scholar] [CrossRef]
- Bak, P.; Tang, C.; Wiesenfeld, K. Self-organized criticality: An explanation of 1/f noise. Phys. Rev. Lett. 1987, 59, 381–384. [Google Scholar] [CrossRef]
- Priesemann, V.; Wibral, M.; Valderrama, M.; Pröpper, R.; Le Van Quyen, M.; Geisel, T.; Triesch, J.; Nikolić, D.; Munk, M.H.J. Spike avalanches in vivo suggest a driven, slightly subcritical brain state. Front. Syst. Neurosci. 2014, 8, 108. [Google Scholar] [CrossRef] [PubMed]
- Shew, W.L.; Plenz, D. The functional benefits of criticality in the cortex. Neuroscientist 2013, 19, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Hesse, J.; Gross, T. Self-organized criticality as a fudamental property of neural systems. Front. Syst. Neurosci. 2014, 8, 166. [Google Scholar] [CrossRef] [PubMed]
- Beggs, J.M. The criticality hypothesis: How local cortical networks might optimize information processing. Phil. Trans. R. Soc. Lond. A Math. Phys. Eng. Sci. 2008, 366, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Allegrini, P.; Paradisi, P.; Menicucci, D.; Gemignani, A. Fractal complexity in spontaneous eeg metastable-state transitions: New vistas on integrated neural dynamics. Front. Physiol. 2010, 1, 128. [Google Scholar] [CrossRef] [PubMed]
- Expert, P.; Lambiotte, R.; Chialvo, D.R.; Christensen, K.; Jensen, H.J.; Sharp, D.J.; Turkheimer, F. Self-similar correlation function in brain resting-state functional magnetic resonance imaging. J. R. Soc. Interface 2011, 8, 472–479. [Google Scholar] [CrossRef]
- Haimovici, A.; Tagliazucchi, E.; Balenzuela, P.; Chialvo, D.R. Brain organization into resting state networks emerges at criticality on a model of the human connectome. Phys. Rev. Lett. 2013, 110, 178101. [Google Scholar] [CrossRef]
- Kitzbichler, M.G.; Smith, M.L.; Christensen, S.R.; Bullmore, E. Broadband criticality of human brain network synchronization. PLoS Comput. Biol. 2009, 5, e1000314. [Google Scholar] [CrossRef]
- Linkenkaer-Hansen, K.; Nikouline, V.V.; Palva, J.M.; Ilmoniemi, R.J. Long-range temporal correlations and scaling behavior in human brain oscillations. J. Neurosci. 2001, 21, 1370–1377. [Google Scholar] [CrossRef]
- Palva, J.M.; Zhigalov, A.; Hirvonen, J.; Korhonen, O.; Linkenkaer-Hansen, K.; Palva, S. Neuronal long-range temporal correlations and avalanche dynamics are correlated with behavioral scaling laws. Proc. Natl. Acad. Sci. USA 2013, 110, 3585–3590. [Google Scholar] [CrossRef]
- Shriki, O.; Alstott, J.; Carver, F.; Holroyd, T.; Henson, R.N.; Smith, M.L.; Coppola, R.; Bullmore, E.; Plenz, D. Neuronal avalanches in the resting meg of the human brain. J. Neurosci. 2013, 33, 7079–7090. [Google Scholar] [CrossRef] [PubMed]
- Clauset, A.; Shalizi, C.R.; Newman, M.E.J. Power-law distributions in empirical data. SIAM Rev. 2009, 51, 661–703. [Google Scholar] [CrossRef]
- Panzeri, S.; Senatore, R.; Montemurro, M.A.; Petersen, R.S. Correcting for the sampling bias problem in spike train information measures. J. Neurophysiol. 2007, 98, 1064–1072. [Google Scholar] [CrossRef] [PubMed]
- Ayyildiz, E.; Gazu, V.P.; Wit, E. A short note in resolving singularity problems in covariance matrices. Int. J. Stat. Probab. 2012, 1, 113–118. [Google Scholar] [CrossRef]
- Nunez, P.L.; Srinivasan, R. Electric Fields of the Brain: The Neurophysics of EEG, 2nd ed.; Oxford University Press, Inc.: New York, NY, USA, 2006. [Google Scholar]
- Nunez, P.L.; Wingeier, B.M.; Silberstein, R.B. Spatial-temporal structures of human alpha rhythms: Theory, microcurrent sources, multiscale measurements, and global binding of local networks. Hum. Brain Mapp. 2001, 13, 125–164. [Google Scholar] [CrossRef]
- Srinivasan, R.; Winter, W.R.; Nunez, P.L. Source analysis of EEG oscillations using high-resolution EEG and MEG. Prog. Brain Res. 2006, 159, 29–42. [Google Scholar] [CrossRef]
- Groppe, D.M.; Bickel, S.; Keller, C.J.; Jain, S.K.; Hwang, S.T.; Harden, C.; Mehta, A.D. Dominant frequencies of resting human brain activity as measured by the electrocorticogram. Neuroimage 2013, 79, 223–233. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
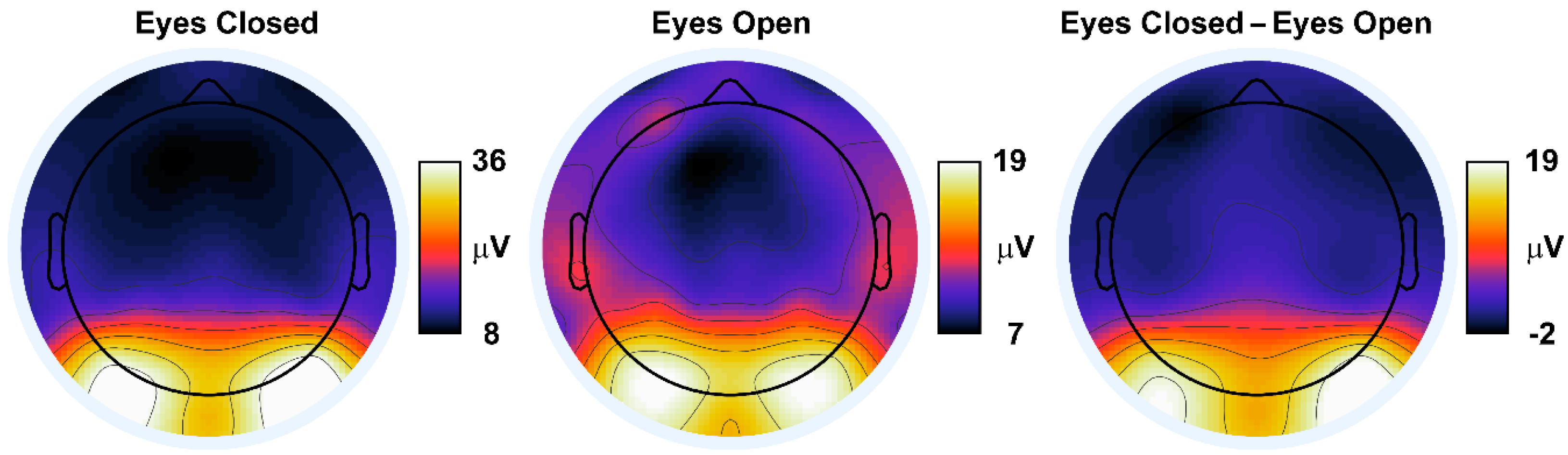{kind=link}
{kind=link}
| Observed Data | Surrogate Data | ||||
|---|---|---|---|---|---|
| Task | Condition | I(X) | CI(X) | I(X) | CI(X) |
| 1-Exemplar Task | Prestimulus | 214.59 (1.07) | 258.41 (0.83) | 102.54 (100.00, 105.09) | 239.95 (237.65, 242.25) |
| Poststimulus | 219.71 (1.39) | 254.24 (0.94) | 101.74 (99.23, 104.24) | 240.99 (238.30, 243.68) | |
| 2-Exemplar Task | Prestimulus | 214.21 (1.09) | 258.49 (0.78) | 102.72 (100.02, 105.42) | 239.66 (237.31, 242.00) |
| Poststimulus | 218.66 (1.30) | 254.65 (0.88) | 101.28 (98.90, 103.66) | 240.26 (237.63, 242.88) | |
| Resting Task | Eyes Open | 228.82 (1.20) | 242.49 (0.89) | 106.29 (103.56,109.01) | 244.89 (242.51,247.27) |
| Eyes Closed | 232.22 (1.08) | 240.73 (0.83) | 101.74 (107.90,113.88) | 244.82 (242.45,247.18) | |
| Task | EEG Measure | Effect | F | P | ε | η2P |
|---|---|---|---|---|---|---|
| Categorization | I(X) | Task | 1.58 | 0.228 | – | 0.10 |
| TI | 92.59 | 0.001 | – | 0.86 | ||
| Task x TI | 3.3 | 0.097 | – | 0.17 | ||
| CI(X) | Task | 0.41 | 0.532 | – | 0.03 | |
| TI | 190.59 | 0.001 | – | 0.93 | ||
| Task x TI | 2.10 | 0.168 | – | 0.12 | ||
| Resting State | I(X) | RS | 34.3 | 0.001 | – | 0.70 |
| CI(X) | RS | 22.55 | 0.001 | – | 0.60 | |
| Categorization vs. Resting State | I(X) | DC | 502.99 | * 0.001 | 0.94 | 0.97 |
| CI(X) | DC | 1738.45 | * 0.001 | 0.83 | 0.99 |
| Observed Data | Surrogate Data | ||||
|---|---|---|---|---|---|
| Task | Condition | I(X) | CI(X) | I(X) | CI(X) |
| 1-Exemplar Task | Prestimulus | 213.56 (1.81) | 259.75 (1.47) | 103.01 (100.48, 105.55) | 233.07 (229.53, 236.60) |
| Poststimulus | 242.20 (3.11) | 241.34 (2.36) | 110.73 (107.68, 113.78) | 232.90 (229.71, 236.09) | |
| 2-Exemplar Task | Prestimulus | 212.49 (1.45) | 258.67 (1.59) | 102.73 (100.08, 105.38) | 233.34 (230.14, 236.54) |
| Poststimulus | 246.66 (1.83) | 238.09 (2.44) | 112.05 (108.75, 115.36) | 233.06 (228.83, 237.29) | |
| EEG Type | EEG Measure | Effect | F | P | η2P |
|---|---|---|---|---|---|
| Evoked | I(X) | Task | 2.91 | 0.109 | 0.16 |
| TI | 282.84 | 0.001 | 0.95 | ||
| Task x TI | 9.04 | 0.009 | 0.38 | ||
| CI(X) | Task | 2.90 | 0.109 | 0.16 | |
| TI | 64.49 | 0.001 | 0.81 | ||
| Task x TI | 0.91 | 0.356 | 0.06 | ||
| Induced vs.Evoked | EEG | 36.74 | 0.001 | 0.71 | |
| I(X) | TI | 427.41 | 0.001 | 0.97 | |
| EEG x TI | 162.93 | 0.001 | 0.92 | ||
| EEG | 21.56 | 0.001 | 0.59 | ||
| CI(X) | TI | 103.38 | 0.001 | 0.87 | |
| EEG x TI | 36.30 | 0.001 | 0.71 |
| Task | Data Condition | GFPInduced | GFPEvoked | ΛInduced | ΛEvoked |
|---|---|---|---|---|---|
| 1-Exemplar Task | Prestimulus | 16.82 (1.48) | 3.95 (0.36) | 0.209 (0.006) | 0.208 (0.015) |
| Poststimulus | 15.83 (1.38) | 4.09 (0.37) | 0.199 (0.006) | 0.248 (0.015) | |
| 2-Exemplar Task | Prestimulus | 17.51 (1.68) | 4.37 (0.36) | 0.218 (0.007) | 0.224 (0.010) |
| Poststimulus | 15.97 (1.49) | 4.47 (0.37) | 0.201 (0.006) | 0.243 (0.016) | |
| Resting Task | Eyes Open | 23.50 (2.07) | – | 0.202 (0.007) | – |
| Eyes Closed | 20.88 (1.79) | – | 0.223 (0.010) | – |
| Task | EEG Measure | Effect | F | P | ε | η2P |
|---|---|---|---|---|---|---|
| Categorization | GFPInduced | Task | 0.46 | 0.508 | – | 0.03 |
| TI | 41.41 | 0.001 | – | 0.73 | ||
| Task x TI | 7.02 | 0.018 | – | 0.32 | ||
| ΛInduced | Task | 3.16 | 0.096 | – | 0.17 | |
| TI | 31.05 | 0.001 | – | 0.67 | ||
| Task x TI | 6.80 | 0.020 | – | 0.31 | ||
| Resting State | GFPInduced | RS | 33.42 | 0.001 | – | 0.69 |
| ΛInduced | RS | 6.79 | 0.020 | – | 0.31 | |
| Categorization vs. Resting State | GFPInduced | DC | 29.94 | * 0.001 | 0.35 | 0.67 |
| ΛInduced | DC | 6.47 | * 0.010 | 0.49 | 0.30 |
| EEG Type | EEG Measure | Effect | F | P | η2P |
|---|---|---|---|---|---|
| Evoked | GFPEvoked | Task | 4.98 | 0.041 | 0.25 |
| TI | 8.14 | 0.012 | 0.35 | ||
| Task x TI | 1.15 | 0.300 | 0.07 | ||
| ΛEvoked | Task | 0.83 | 0.378 | 0.05 | |
| TI | 5.98 | 0.027 | 0.29 | ||
| Task x TI | 0.94 | 0.349 | 0.06 | ||
| Induced vs. Evoked | EEG | 72.47 | 0.001 | 0.83 | |
| GFPEvoked | TI | 33.71 | 0.001 | 0.69 | |
| EEG x TI | 45.87 | 0.001 | 0.75 | ||
| EEG | 5.63 | 0.031 | 0.27 | ||
| ΛEvoked | TI | 1.36 | 0.262 | 0.08 | |
| EEG x TI | 14.76 | 0.002 | 0.50 |
| IV | DV | β | Wald χ2 | P |
|---|---|---|---|---|
| GFPInduced | I(X) | 0.56 (0.11) | 27.90 | 0.004 |
| CI(X) | −0.52 (0.11) | 21.28 | 0.004 | |
| ΛInduced | I(X) | −0.27 (0.08) | 11.12 | 0.004 |
| CI(X) | 0.33 (0.07) | 22.18 | 0.004 | |
| GFPEvoked | I(X) | 0.28 (0.04) | 40.44 | 0.004 |
| CI(X) | −0.29 (0.08) | 14.18 | 0.004 | |
| ΛEvoked | I(X) | −0.001 (0.11) | 0.00 | 0.965 |
| CI(X) | 0.08 (0.13) | 0.36 | 0.546 |
| Data Condition | Prestimulus | Poststimulus | F | p | η2P |
|---|---|---|---|---|---|
| I(X)Induced | 221.08 (0.09) | 237.25 (0.20) | 7007.88 | 0.001 | 0.99 |
| CI(X)Induced | 253.43 (0.16) | 244.24 (0.18) | 1849.51 | 0.001 | 0.99 |
| I(X)Evoked | 221.50 (1.08) | 246.78 (1.54) | 313.07 | 0.001 | 0.95 |
| CI(X)Evoked | 255.28 (1.05) | 240.36 (1.81) | 48.87 | 0.001 | 0.77 |
| GFPInduced | 30.63 (0.15) | 27.83 (0.09) | 310.58 | 0.001 | 0.95 |
| GFPEvoked | 9.40 (0.37) | 9.64 (0.39) | 33.03 | 0.001 | 0.69 |
| ΛInduced | 0.195 (0.001) | 0.152 (0.001) | 1996.46 | 0.001 | 0.99 |
| ΛEvoked | 0.173 (0.012) | 0.225 (0.007) | 16.55 | 0.001 | 0.53 |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trujillo, L.T. K-th Nearest Neighbor (KNN) Entropy Estimates of Complexity and Integration from Ongoing and Stimulus-Evoked Electroencephalographic (EEG) Recordings of the Human Brain. Entropy 2019, 21, 61. https://doi.org/10.3390/e21010061
Trujillo LT. K-th Nearest Neighbor (KNN) Entropy Estimates of Complexity and Integration from Ongoing and Stimulus-Evoked Electroencephalographic (EEG) Recordings of the Human Brain. Entropy. 2019; 21(1):61. https://doi.org/10.3390/e21010061
Chicago/Turabian StyleTrujillo, Logan T. 2019. "K-th Nearest Neighbor (KNN) Entropy Estimates of Complexity and Integration from Ongoing and Stimulus-Evoked Electroencephalographic (EEG) Recordings of the Human Brain" Entropy 21, no. 1: 61. https://doi.org/10.3390/e21010061
APA StyleTrujillo, L. T. (2019). K-th Nearest Neighbor (KNN) Entropy Estimates of Complexity and Integration from Ongoing and Stimulus-Evoked Electroencephalographic (EEG) Recordings of the Human Brain. Entropy, 21(1), 61. https://doi.org/10.3390/e21010061