Structural Patterns of Rhamnogalacturonans Modulating Hsp-27 Expression in Cultured Human Keratinocytes

Abstract
:Introduction
Results and Discussion

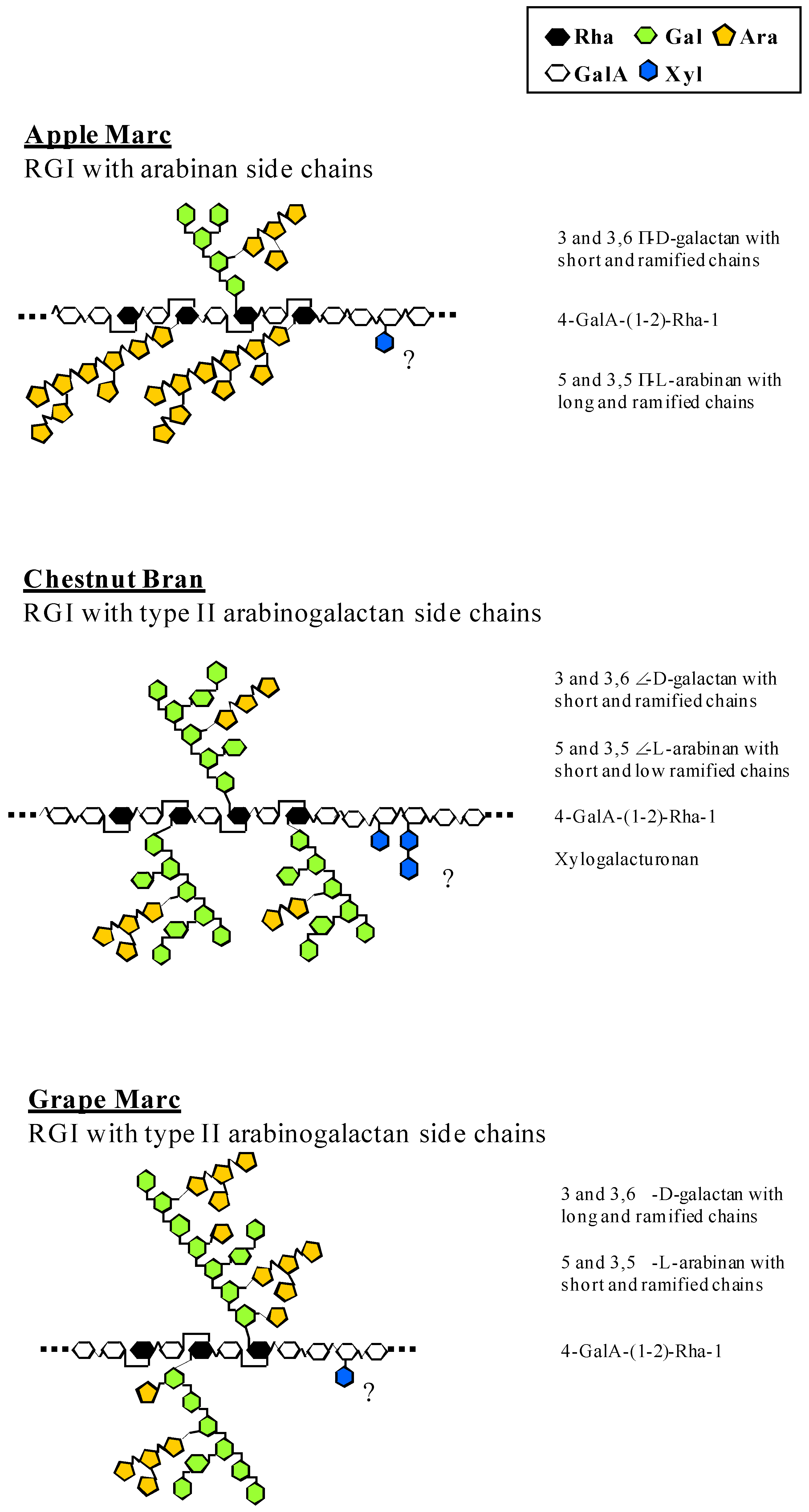{kind=link}
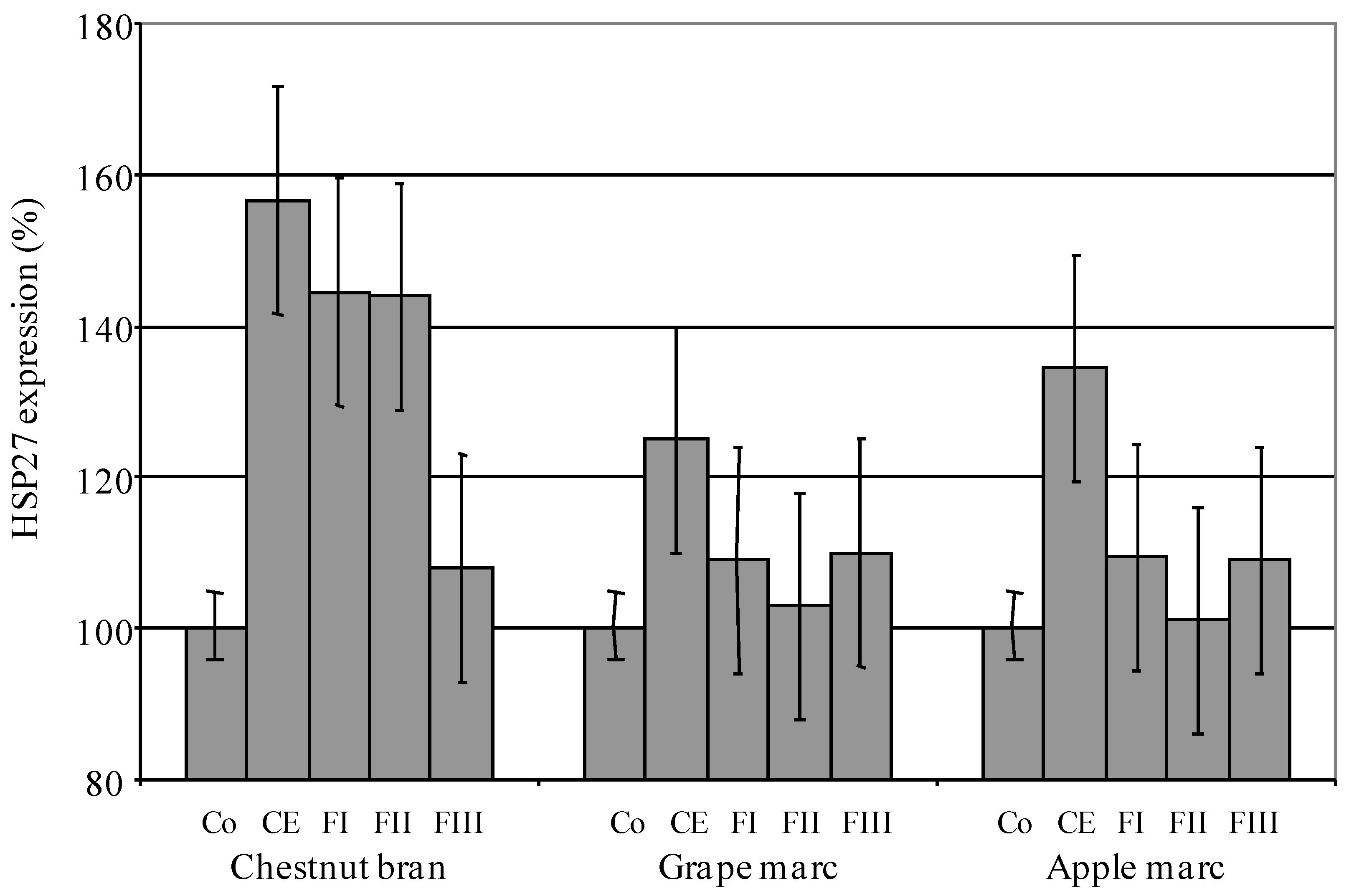{kind=link}
| Plant materials and fractions | Yield of extraction (W/W Total Sugar / Raw Material) | %[UA] |
|---|---|---|
| Chestnut bran | ||
| Crude extract | 46.4 | 8.4 |
| FI | 0.6 | 50 |
| FII | 1.3 | 80 |
| FIII | 44.5 | 0 |
| Apple marc | ||
| Crude extract | 6.8 | 69.5 |
| FI | 3.3 | 47.1 |
| FII | 2.7 | 86.8 |
| FIII | 0.8 | 8.5 |
| Grape marc | ||
| Crude extract | 24.0 | 36.9 |
| FI | 8.0 | 42.9 |
| FII | 6.9 | 54.6 |
| FIII | 9.1 | 34.0 |
| Plant materials and fractions | Ara | Rha | Xyl | Man | Gal | Gal A | Glc | Glc A |
|---|---|---|---|---|---|---|---|---|
| Chestnut bran | ||||||||
| Crude extract | t | t | 0 | 0 | 4.0 | t | 96.0 | 0 |
| FI | 14.6 | 16.6 | 9.4 | 1.4 | 15.0 | 38.2 | 2.2 | 2.5 |
| FII | 6.4 | 3.6 | 10.2 | 0.8 | 1.7 | 65.9 | 11.3 | 0 |
| FIII | 0 | 0 | 0 | 0 | 0 | 0 | 100 | 0 |
| Apple marc | ||||||||
| Crude extract | 36.4 | 6.3 | 0 | 0 | 6.8 | 50.5 | 0 | 0 |
| FI | 29.1 | 6.0 | 2.5 | 2.3 | 3.9 | 53 | 1.4 | 1.7 |
| FII | 10.0 | 1.9 | 1.2 | 0 | 0.5 | 81.6 | 0 | 4.8 |
| FIII | 34.8 | 5.9 | 9.7 | 0.7 | 20.7 | 3.5 | 24.7 | 0 |
| Grape marc | ||||||||
| Crude extract | 26.2 | 9.1 | 5.7 | 20.2 | 13.0 | 21.2 | 4.5 | 0 |
| FI | 24.5 | 11.2 | 5.5 | 9.4 | 15.6 | 29.0 | 1.3 | 3.4 |
| FII | 18.9 | 7.8 | 7.1 | 7.0 | 8.9 | 41.9 | 0 | 8.4 |
| FIII | 12.9 | 3.9 | 4.1 | 44.2 | 14.1 | t | 20.8 | 0 |
| Carbohydrates | Chestnut bran | Apple marc | Grape marc | |
|---|---|---|---|---|
| FI | FII | FI | FI | |
| t-Ara | 14.5 | 16.9 | 35.0 | 28.0 |
| 5-Ara | 14.5 | 15.2 | 23.2 | 12.7 |
| 3,5-Ara | 2.7 | 11.1 | 14.6 | 8.5 |
| Galactose | ||||
| t-Gal | 13.5 | 10.7 | 5.8 | 6.7 |
| 3-Gal | 9.3 | 2.3 | 3.1 | 11.6 |
| 4-Gal | 13.4 | 10.8 | 10.9 | 4.3 |
| 6-Gal | 5.7 | 0 | 1.1 | 3.7 |
| 3,6-Gal | 12.9 | 3.3 | 3.7 | 20.5 |
| 4,6-Gal | 0 | 4.7 | 0 | 0 |
| Xylose | ||||
| t-Xyl | 8.6 | 18.7 | 2.6 | 4.0 |
| 4-Xyl | 4.9 | 6.3 | 0 | 0 |

| Concentrations (%W/V) | Co | 0.004 | 0.01 | 0.02 | 0.04 | 0.1 | 0.2 | 0.4 |
|---|---|---|---|---|---|---|---|---|
| Chestnut bran | 102 | 99 | 98 | 100 | 117 | 92 | 78 | 60 |
| Apple marc | 100 | 123 | 130 | 121 | 84 | 13 | 10 | 10 |
| Grape marc | 103 | 103 | 99 | 100 | 50 | 35 | 20 | 15 |

Conclusions
Experimental
Biological material
Chemicals and enzymes
Isolation, enzymatic hydrolysis and fractionation of rhamnogalacturonans
Composition of polysaccharides
Monosaccharide linkage analysis
Submerged keratinocyte cultures
Characterization of keratinocyte physiology
Keratinocyte viability
Keratinocyte differentiation, quantification of the differentiation-specific protein HSP-27
Acknowledgements
References
- Gloaguen, V.; Krausz, P. Plant polysaccharides: a biologically active class of molecules. SOWF J. 2004, 130, 20–26. [Google Scholar]
- Paulsen, BS. Biologically active polysaccharides as possible lead compounds. Phytochem. Rev. 2002, 1, 379–387. [Google Scholar] [CrossRef]
- Mansell, M.A. Polysaccharides in skin care. Cosmet. Toiletries 1994, 109, 67–72. [Google Scholar]
- Pauly, M.; Freis, O.; Pauly, G. Galactomannan and xyloglucan: bio-active polysaccharides. Cosmet. Toiletries 1999, 114, 65–78. [Google Scholar]
- Deters, A.; Dauer, A.; Schnetz, E.; Fartasch, M.; Hensel, A. High molecular compounds (polysaccharides and proanthocyanidins) from Hamamelis virginiana bark: influence on human skin keratinocyte proliferation and differentiation and influence on irritated skin. Phytochemistry 2001, 58, 949–958. [Google Scholar] [CrossRef]
- Deters, A.; Schröder, K.R.; Smiatek, T.; Hensel, A. Ispaghula (Plantago ovata) seed husk polysaccharides promote proliferation of human epithelial cells (skin, keratinocytes and fibroblasts) via enhanced growth factor receptors and energy production. Planta Med. 2005, 71, 33–39. [Google Scholar] [CrossRef]
- Paulsen, B.S.; Barsett, H. Bioactive pectic polysaccharides. Adv. Polym. Sci. 2005, 186, 69–101. [Google Scholar] [CrossRef]
- Nergard, C.S.; Kiyohara, H.; Reynolds, C.; Thomas-Oates, J.E.; Matsumoto, T.; Yamada, H.; Patel, T.; Petersen, D.; Michaelsen, T.E.; Diallo, D.; Paulsen, B.S. Structures and structure-activity relationship of three mitogenic and complement fixing pectic arabinogalactans from the Malian antiulcer plants Cochlospermum tinctorium A. rich and Vernonia kotschyana Sch. Bip. Ex Walp. Biomacromolecules 2006, 7, 71–79. [Google Scholar] [CrossRef]
- Brunolds, C.; Deters, A.; Knoepfel-Sidler, F.; Hafner, J.; Müller, B.; Hensel, A. Polysaccharides from Hibiscus sabdariffa flowers stimulate proliferation and differentiation of human keratinocytes. Planta Med. 2004, 70, 370–3. [Google Scholar]
- Deters, A.; Schröder, K.R.; Hensel, A. Kiwi fruit (Actinidia chinensis L.) polysaccharides exert stimulating effects on cell proliferation via enhanced growth factor receptors, energy production, and collagen synthesis of human keratinocytes, fibroblasts, and skin equivalents. J. Cell Physiol. 2005, 202, 717–722. [Google Scholar] [CrossRef]
- Deters, A.; Lengsfeld, C.; Hensel, A. Oligo- and polysaccharides exhibit a structure-dependent bioactivity on human keratinocytes in vitro. J. Ethnopharmacol. 2005, 102, 391–399. [Google Scholar] [CrossRef]
- Paufique, J. Active ingredient for skin treatments, method for the production thereof and use of the same. Patent WO 03/05925, 2004. [Google Scholar]
- Jouandeaud, M.; Moreau, L.; Chauprade, C.; Guichard, N.; Closs, B.; Gloaguen, V. Barrier function and dry skin: use of rhamnogalacturonans as a new therapeutic strategy. SOFW J. 2004, 130, 2–7. [Google Scholar]
- Schols, H.A.; Vierhuis, E.; Bakx, E.J.; Voragen, A.G.J. Different populations of pectic hairy region occur in apple cell walls. Carbohydr. Res. 1995, 275, 343–360. [Google Scholar] [CrossRef]
- Brillouet, J.M. A study of pectic polysaccharides in musts from various mature grapes grown in the Pech Rouge experimental vineyards. Biochimie 1987, 69, 713–721. [Google Scholar] [CrossRef]
- Kindas, M.I.; Trautinger, F. Increased expression of the Mr 27000 heat shock protein (HSP 27) in vitro differentiated normal human keratinocytes. Cell Growth Differ. 1994, 5, 777–781. [Google Scholar]
- Jonack, C.; Klosner, G.; Kokesch, C.; Fodinger, D.; Honigsmann, R.; Trautinger, F. Subcorneal colocalization of the small heat shock protein, HSP 27, with keratins and proteins of the cornified cell envelope. Br. J. Dermatol. 2002, 147, 13–9. [Google Scholar] [CrossRef]
- Ebringerova, A.; Kardosova, A.; Hromadkova, Z.; Malovikova, A.; Hribalova, V. Immuno-modulatory activity of acidic xylans in relation to their structural and molecular properties. Int. J. Biol. Macromol. 2002, 30, 1–6. [Google Scholar] [CrossRef]
- Moine, C.; Krausz, P.; Chaleix, V.; Kraemer, M.; Gloaguen, V. Structural characterization and cytotoxic properties of a 4−O−Methylglucuronoxylan from Castanea sativa. J. Nat. Prod. 2007, 70, 60–6. [Google Scholar] [CrossRef]
- Gloaguen, V.; Morvan, H.; Hoffmann, L.; Sainte Catherine, O.; Kraemer, M.; Krausz, P. Bioactive Capsular Polysaccharide from the Thermophilic Cyanophyte/Cyanobacterium Mastigocladus laminosus - Cytotoxic Properties. Planta Med. 2007, 73, 1402–1407. [Google Scholar] [CrossRef]
- Schols, H.A.; Bakx, E.J.; Schipper, D.; Voragen, A.G.J. A xylogalacturonan subunit present in the modified hairy region of apple pectin. Carbohydr. Res. 1995, 279, 265–279. [Google Scholar] [CrossRef]
- Vincken, J.P.; Schols, H.A.; Oomen, R.J.F.J.; McCann, M.C.; Ulvskov, P.; Voragen, A.G.J.; Visser, R.G.F. If homogalacturonan were a side chain of rhamnoglacturonan I. Implications for cell wall architecture. Plant Physiol. 2003, 132, 1781–1789. [Google Scholar] [CrossRef]
- Oechslin, R.; Lutz, M.V.; Amado, R. Pectic substances isolated from apple cellulosic residue: structural characterization of a new type of rhamnogalacturonan. Carbohydr. Pol. 2003, 51, 301–310. [Google Scholar] [CrossRef]
- Inngjerdingen, K.T.; Debes, S.C.; Inngjerdingen, M.; Hokputsa, S.; Harding, S.E.; Rolstad, B.; Michaelsen, T.E.; Diallo, D.; Paulsen, B.S. Bioactive pectic polysaccharide from Glinus oppositifolius (L.) Aug. DC., a Malian medicinal plant, isolation and partial characterization. J. Ethnopharmacol. 2005, 101, 204–214. [Google Scholar]
- Inngjerdingen, K.T.; Coulibaly, A.; Diallo, D.; Michaelsen, T.E.; Paulsen, B.S. A complement fixing polysaccharide from Biophytum petersianum Klotzsc, a medicinal plant from Mali, West Africa. Biomacromolecules 2006, 7, 48–53. [Google Scholar]
- Duan, J.; Zheng, Y.; Dong, Q.; Fang, J. Structural analysis of a pectic polysaccharide from the leaves of Diospyros kaki. Phytochemistry 2004, 65, 609–615. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, KA.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–256. [Google Scholar]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Kamerling, J.P.; Gerwig, G.J.; Vliegenhart, J.F.G.; Clamp, J.R. Characterization by gas liquid chromatography mass spectrometry of pertrimethylsilyl glycosides obtained in the methanolysis of glycoproteins and glycolipids. Biochem. J. 1975, 151, 491–495. [Google Scholar]
- Montreuil, J.; Bouquelet, S.; Debray, H.; Fournet, B.; Spik, G.; Strecker, G. Glycoproteins. In Carbohydrate Analysis: a Practical Approach; Chaplin, M. F., Kennedy, J. F., Eds.; IRL Press: Oxford, 1986; pp. 143–204. [Google Scholar]
- Parente, J.P.; Cardon, P.; Leroy, Y.; Montreuil, J.; Fournet, B.; Ricart, G. A convenient method for methylation of glycoprotein glycans in small amounts by using lithium methylsulfinyl carbanion. Carbohydr. Res. 1985, 141, 41–47. [Google Scholar] [CrossRef]
- Kim, J.B.; Carpita, N. Changes in esterification of the uronic acid groups of cell wall polysaccharides during elongation of maize coleoptile. Plant Physiol. 1992, 98, 646–653. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J. Immunol. Meth. 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Sample availability: Active chestnut bran rhamnogalacturonan is commercially available from the SILAB Company under the tradename Recoverine®.
© 2008 by the authors. Licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gloaguen, V.; Krausz, P.; Brudieux, V.; Closs, B.; Leroy, Y.; Guerardel, Y. Structural Patterns of Rhamnogalacturonans Modulating Hsp-27 Expression in Cultured Human Keratinocytes. Molecules 2008, 13, 1207-1218. https://doi.org/10.3390/molecules13051207
Gloaguen V, Krausz P, Brudieux V, Closs B, Leroy Y, Guerardel Y. Structural Patterns of Rhamnogalacturonans Modulating Hsp-27 Expression in Cultured Human Keratinocytes. Molecules. 2008; 13(5):1207-1218. https://doi.org/10.3390/molecules13051207
Chicago/Turabian StyleGloaguen, Vincent, Pierre Krausz, Véronique Brudieux, Brigitte Closs, Yves Leroy, and Yann Guerardel. 2008. "Structural Patterns of Rhamnogalacturonans Modulating Hsp-27 Expression in Cultured Human Keratinocytes" Molecules 13, no. 5: 1207-1218. https://doi.org/10.3390/molecules13051207