Fucoidan: Structure and Bioactivity

Abstract
:Introduction
Structure
Fucoidans mainly composed of fucose and sulfate

Fucoidans from other brown seaweeds

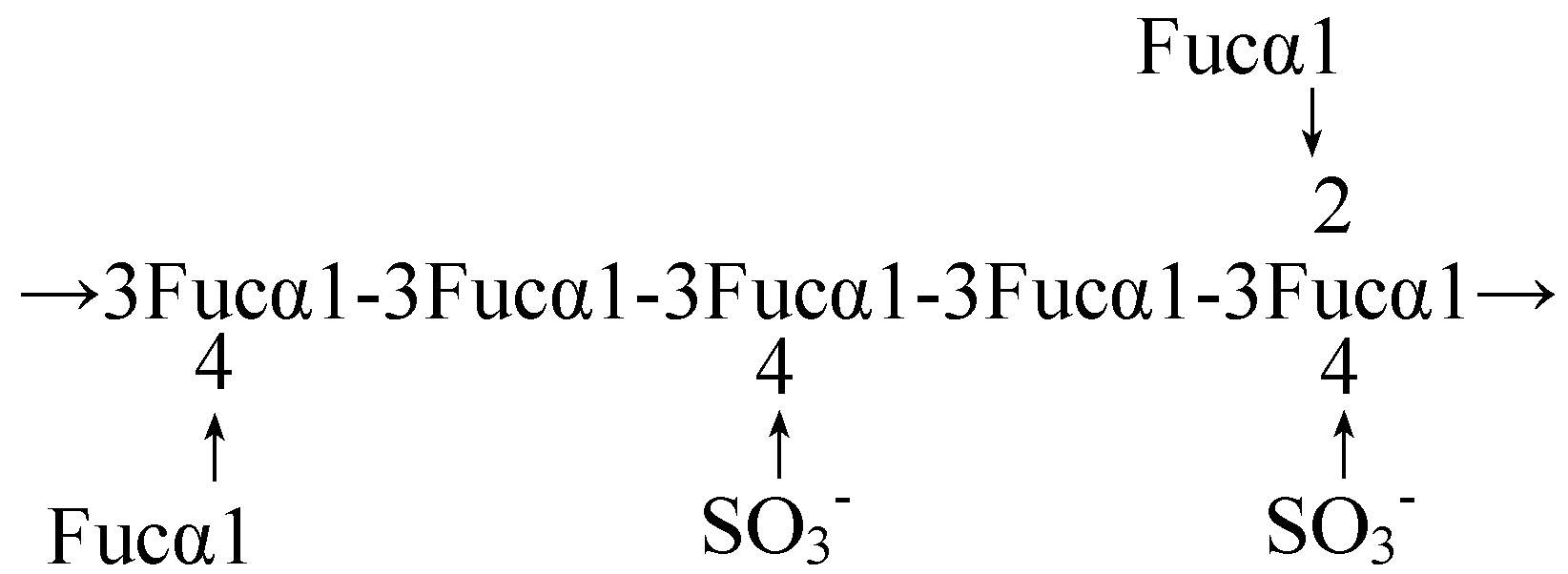{kind=link}
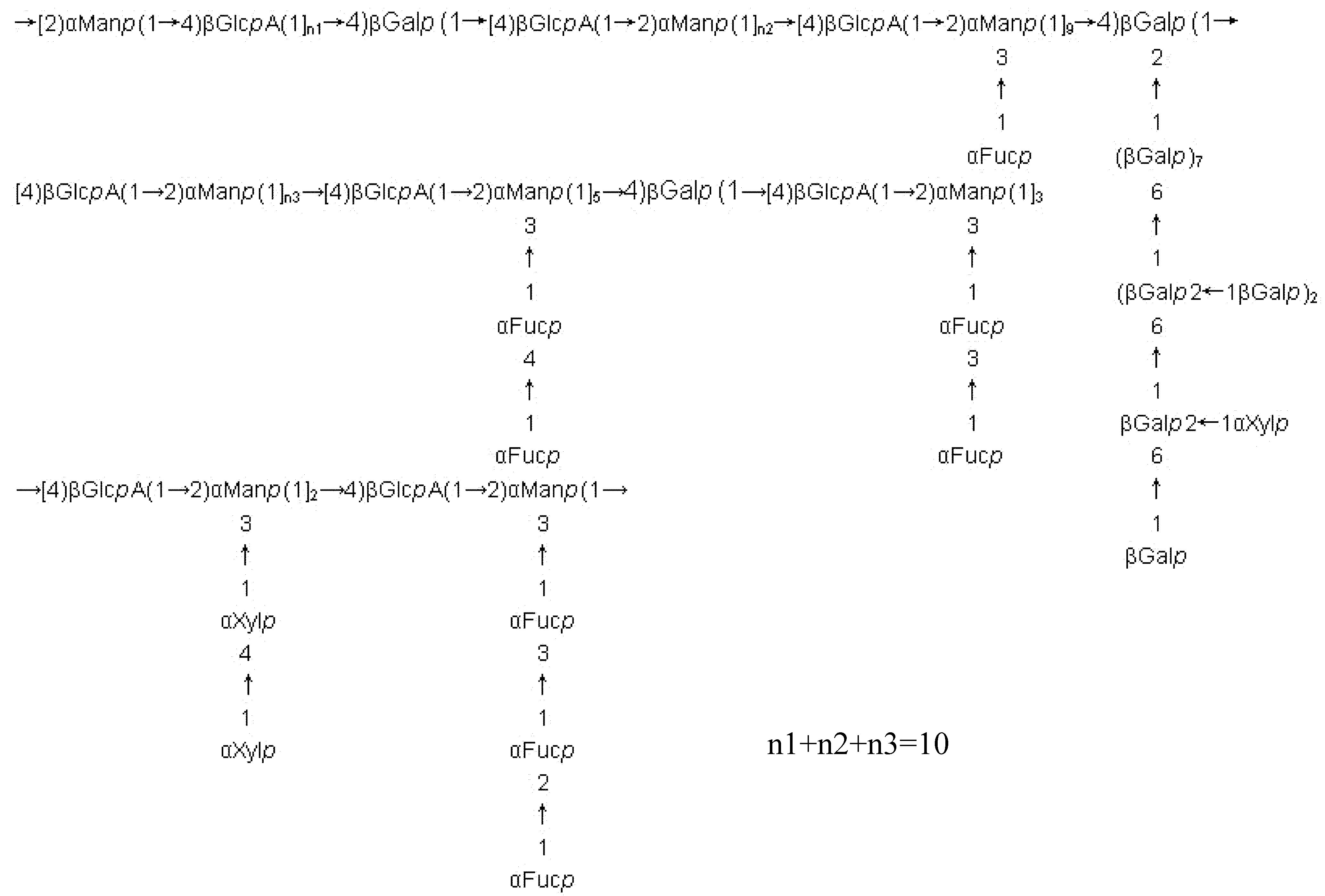{kind=link}
| Brown seaweed | Chemical composition |
|---|---|
| F. vesiculosus [3,4] | fucose, sulfate |
| F. evanescens C.Ag. [2] | fucose/sulfate/acetate (1/1.23/0.36) |
| F. distichus [9] | fucose/sulfate/acetate (1/1.21/0.08) |
| F. serratus L. [10] | fucose/sulfate/acetate (1/1/0.1) |
| Lessonia vadosa [20] | fucose/sulfate (1/1.12) |
| Macrocytis pyrifera [3] | fucose/galactose (18/1), sulfate |
| Pelvetia wrightii [21] | fucose/galactose (10/1), sulfate |
| Undaria pinnatifida (Mekabu) [22] | fucose/galactose (1/1.1), sulfate |
| Ascophyllum nodosum [23] | fucose(49%), xylose(10%), GlcA(11%), sulfate |
| Himanthalia lorea and Bifurcaria bifurcate [15] | fucose, xylose, GlcA, sulfate |
| Padina pavonia [16,17] | fucose, xylose, mannose, glucose, galactose, sulfate |
| Laminaria angustata [24] | fucose/galactose/sulfate (9/1/9) |
| Ecklonia kurome [25] | fucose, galactose, mannose, xylose, GlcA, sulfate |
| Sargassum stenophyllum [26] | fucose, galactose, mannose, GlcA, glucose, xylose, sulfate |
| Adenocytis utricularis [27] | fucose, galactose, mannose, sulfate |
| Hizikia fusiforme [28] | fucose, galactose, mannose, xylose, GlcA, sulfate |
| Dictyota menstrualis [29] | fucose/xylose/uronic acid/galactose/sulfate (1/0.8/0.7/0.8/0.4) and (1/0.3/0.4/1.5/1.3) |
| Spatoglossum schroederi [30] | fucose/xylose/galactose/sulfate (1/0.5/2/2) |
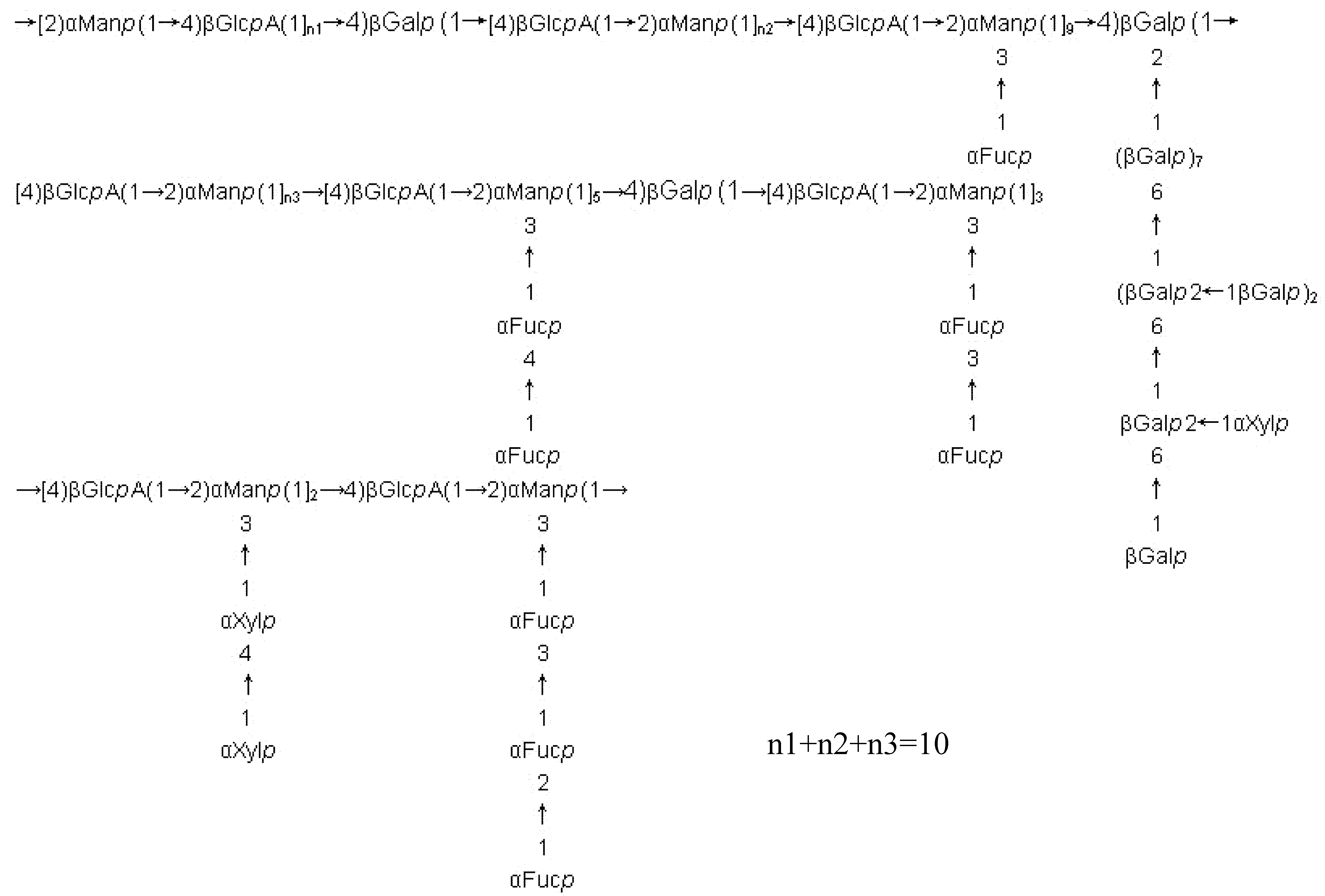
Position of sulfate groups
Fucosidase
Complexity of fucoidan structures
Bioactivity
Anticoagulant and antithrombotic activity
Antivirus activity
Antitumor and immunomodulatory activity
Antioxidant activity
Reducing blood lipids
Anticomplementary activity
Therapeutic potential in surgery
Anti-inflammatory
Gastric protection
Against hepatopathy
Against uropathy and renalpathy
Conclusions
Acknowledgements
References
- Chizhov, A.O.; Dell, A; Morris, H.R. A study of fucoidan from the brown seaweed Chorda filum. Carbohydr. Res. 1999, 320, 108–119. [Google Scholar] [CrossRef]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E. Structure of a fucoidan from the brown seaweed Fucus evanescens C.Ag. Carbohydr. Res. 2002, 337, 719–730. [Google Scholar] [CrossRef]
- Black, W.A.P.; Dewar, E.T.; Woodward, F.N. Manufacture of algal chemicals. IV. -Laboratory-scale isolation of fucoidin from brown marine algae. J. Sci. Food Agric. 1952, 3, 122–129. [Google Scholar] [CrossRef]
- Nishino, T.; Nishioka, C.; Ura, H. Isolation and partial characterization of a novel amino sugar-containing fucan sulfate from commercial Fucus vesiculosus fucoidan. Carbohydr. Res. 1994, 255, 213–224. [Google Scholar] [CrossRef]
- Conchie, J.; Percival, E.G.V. Fucoidin part II. The hydrolysis of a methylated fucoidin prepared from Fucus vesiculosus. J. Chem. Soc. 1950, 827–833. [Google Scholar] [CrossRef]
- O'Neill, A.N. Degradative studies on fucoidan. J. Amer. Chem. Soc. 1954, 76, 5074–5076. [Google Scholar] [CrossRef]
- Anno, K.; Seno, N.; Ota, M. Isolation of L-fucose 4-sulfate from fucoidan. Carbohydr. Res. 1970, 13, 167–169. [Google Scholar] [CrossRef]
- Patankar, M.S.; Oehninger, S.; Barnett, T. A revised structure for fucoidan may explain some of its biological activities. J. Biol. Chem. 1993, 268, 21770–21776. [Google Scholar]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. A highly regular fraction of a fucoidan from the brown seaweed Fucus distichus L. Carbohydr. Res. 2004, 339, 511–517. [Google Scholar]
- Bilan, M.I.; Grachev, A.A.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure of a fucoidan from the brown seaweed Fucus serratus L. Carbohydr. Res. 2006, 341, 238–245. [Google Scholar] [CrossRef]
- Adhikaria, U.; Mateub, C.G.; Chattopadhyaya, K.C.; Pujolb, A.; Damonteb, E.B.; Ray, B. Structure and antiviral activity of sulfated fucans from Stoechospermum marginatum. Phytochemistry. 2006, 67, 2474–2482. [Google Scholar] [CrossRef]
- Andrade, L.R.; Salgado, L.T.; Farina, M.; Pereira, M.S.; Mourão, P.A.S.; Amado, F.G.M. J. Structural Biology. 2004, 145, 216–225. [CrossRef]
- Ji, M.H. Algae Chemistry.; Science Publishing House: Beijing, 1997. [Google Scholar]
- Nishino, T.; Nagumo, T.; Kiyohara, H. Structure characterization of a new anticoagulant fucan sulfate from the brown seaweed Ecklonia kurome. Carbohydr. Res. 1991, 211, 77–90. [Google Scholar] [CrossRef]
- Mian, J.; Percival, E. Carbohydrates of the brown seaweeds Himanthalia lorea and Bifurcaria bifurcata Part II. structural studies of the “fucans”. Carbohydr. Res. 1973, 26, 147–161. [Google Scholar] [CrossRef]
- Hussein, M.M.; Abdel, A.; Salem, H.M. Sulfated heteropolysaccharides from Padina pavoia. Phytochemistry 1980, 19, 2131–2132. [Google Scholar] [CrossRef]
- Hussein, M.M.; Abdel, A.; Salem, H.M. Some structural features of a new sulfated heteropolyssaride from Padina pavoia. Phytochemistry 1980, 19, 2133–2135. [Google Scholar] [CrossRef]
- Chevolot, L.; Mulloy, B.; Racqueline, J. A disaccharide repeat unit is the structure structure in fucoidans from two species of brown algae. Carbohydr. Res. 2001, 330, 529–535. [Google Scholar] [CrossRef]
- Marais, M.; Joseleau, J. A fucoidan fraction from Ascophyllum nodosum. Carbohydr. Res. 2001, 336, 155–159. [Google Scholar] [CrossRef]
- Chandía, N.P.; Matsuhiro, B. Characterization of a fucoidan from Lessonia vadosa (Phaeophyta) and its anticoagulant and elicitor properties. Int. J. Biol. Macromol. 2008, 42, 235–240. [Google Scholar] [CrossRef]
- Anno, K.; Terahata, H.; Hayashi, Y. Isolation and purification of fucoidin from brown seaweed Pelvetia wrightii. Agri. Biol. Chem. 1966, 30, 495–499. [Google Scholar] [CrossRef]
- Lee, J.B.; Hayashi, K.; Hashimoto, M.; Nakano, T.; Hayashi, T. Novel antiviral fucoidan from sporophyll of Undaria pinnatifida (Mekabu). Chem. Pharm. Bull. 2004, 52, 1091–1094. [Google Scholar] [CrossRef]
- Percival, E. Glucoroxylofucan, a cell-wall component of Ascophyllum nodosum. Carbohydr. Res. 1968, 7, 272–283. [Google Scholar] [CrossRef]
- Kitamura, K.; Matsuo, M.; Yasui, T. Fucoidan from brown seaweed Laminaria angustata var. longissima. Agric. Biol. Chem. 1991, 55(2), 615–616. [Google Scholar] [CrossRef]
- Nishino, T.; Yokoyama, G.; Dobahi, K. Isolation, purification and characterization of fucose-containing sulfated polysaccharides from the brown seaweed Ecklonia kurome and their blood-anticoagulant activities. Carbohydr. Res. 1989, 186, 119–129. [Google Scholar]
- Duarate, M.; Cardoso, M.; Noseda, M. Structural studies on fucoidans from the brown seaweed Sargassum stenophyllum. Carbohydr. Res. 2001, 333, 281–293. [Google Scholar] [CrossRef]
- Ponce, N.M.A.; Pujol, C.A.; Damonte, E.B. Fucoidans from the brown seaweed Adenocystis utricularis: extraction methods, antiviral activity and structural studies. Carbohydr. Res. 2003, 338, 153–165. [Google Scholar] [CrossRef]
- Li, B.; Xin, J.W.; Sun, J.L.; Xu, S.Y. Structural investigation of a fucoidan containing a fucose-free core from the brown seaweed Hizikia fusiforme. Carbohydr. Res. 2006, 341, 1135–1146. [Google Scholar] [CrossRef]
- Albuquerque, I.R.L.; Queiroz, K.C.S.; Alves, L.G.; Santos, E.A.; Leite, E.L.; Rocha, H.A.O. Heterofucans from Dictyota menstrualis have anticoagulant activity. Braz. J. Med. Biol. Res. 2004, 37, 167–171. [Google Scholar] [CrossRef]
- Rocha, H.A.O.; Moraes, F.A.; Trindade, E.S.; Franco, C.R.C.; Torquato, R.J.S.; Veiga, S.S.; Valente, A.P.; Mourão, P.A.S.; Leite, E.L.; Nader, H.B.; Dietrich, C.P. Structural and hemostatic activities of a sulfated galactofucan from the brown alga Spatoglossum schroederi. J. Biol. Chem. 2005, 280, 1278–41288. [Google Scholar]
- Daniel, R.; Chevolot, L.; Carrascal, M.; Tissot, B.; Mourão, P.A.S.; Abian, J. Electrospray ionization mass spectrometry of oligosaccharides derived from fucoidan of Ascophyllum nodosum. Carbohydr. Res. 2007, 342, 826–834. [Google Scholar] [CrossRef]
- Li, B.; Xu, S.Y. Structural investigation of oligosaccharides in partial acid hydrolyzed products of fucoidan isolated from Hizikia fusiforme. Nat. Prod. Res. Dev. 2007, 19, 550–553. [Google Scholar]
- Zvyagintseva, T.N.; Shevchenko, N.M.; Popivnich, I.B. A new procedure for the separation of water-soluble polysaccharides from brown seaweeds. Carbohydr. Res. 1999, 322, 32–39. [Google Scholar] [CrossRef]
- Li, B.; Gu, X.H.; Xu, S.Y. Determination of the position of sulphate group in sulphated polysaccharides. J. Anal. Sci. 2004, 20, 498–500. [Google Scholar]
- Tissot, B.; Salpin, J.; Martinez, M.; Gaigeot, M.; Daniel, R. Differentiation of the fucoidan sulfated L-fucose isomers constituents by CE-ESIMS and molecular modeling. Carbohydr. Res. 2006, 341, 598–609. [Google Scholar] [CrossRef]
- Grachev, A.A.; Gerbst, A.G.; Ustyuzhanina, N.E.; Shashkov, A.S.; Usov, A.I.; Nifantiev, N.E. NMR investigation of the influence of sulfate groups at C-2 and C-4 on the conformational behavior of fucoidan fragments with homo-(1→3)-linked backbone. J. Carbohydr. Chem. 2006, 25, 315–330. [Google Scholar] [CrossRef]
- Daniel, R.; Berteau, O.; Jozefonvicz, J.; Goasdoue, N. Degradation of algal (Ascophyllum nodosum) fucoidan by an enzymatic activity contained in digestive glands of the marine mollusk Pecten maximus. Carbohydr. Res. 1999, 322, 291–297. [Google Scholar] [CrossRef]
- Berteau, O.; McCort, I.; Goasdoué, N.; Tissot, B.; Daniel, R. Characterization of a new α-L-fucosidase isolated from the marine mollusk Pecten maximus that catalyzes the hydrolysis of α-L-fucose from algal fucoidan (Ascophyllum nodosum). Glycobiology 2002, 12, 273–282. [Google Scholar] [CrossRef]
- Kusaykin, M.I.; Chizhov, A.O.; Grachev, A.A.; Alekseeva, S.A.; Bakunina, I.Y.; Nedashkovskaya, O.I.; Sova, V.V.; Zvyagintseva, T.N. Comparative study of specificity of fucoidanases from marine microorganisms and invertebrates. J. Appl. Phycol. 2006, 18, 369–373. [Google Scholar] [CrossRef]
- Silva, T.M.A.; Alves, L.G.; Queiroz, K.C.S.; Santos, M.G.L.; Marques, C.T.; Chavante, S.F.; Rocha, H.A.O.; Leite, E.L. Partial characterization and anticoagulant activity of a heterofucan from the brown seaweed Padina gymnospora. Braz. J. Med. Biol. Res. 2005, 38, 523–533. [Google Scholar]
- Ribeiro, A.; Vieira, R.P.; Mourão, P.A.S. A sulfated α-L-fucan from sea cucumber. Carbohydr. Res. 1994, 255, 225–240. [Google Scholar] [CrossRef]
- Pavao, M.S.G.; Mourão, P.A.S.; Mulloy, B. Structure of a unique sulfated α-L-galactofucan from the tunicate Clavelina. Carbohydr. Res. 1990, 208, 153–161. [Google Scholar] [CrossRef]
- Mulloy, B.; Ribeino, A.; Alves, A. Sulfated fucans from echinoderms have a regular tetrasaccharide repeating unit defined by specific patterns of sulfation at the O-2 and O-4 position. J. Biol. Chem. 1994, 269, 22113–22123. [Google Scholar]
- Vieira, R.P.; Mourão, P.A.S. Occurrence of a unique fucose-branced chondroitin sulfate in the body wall of a sea cucumber. J. Biol. Chem. 1988, 263, 18176–18183. [Google Scholar]
- Pereira, M.S.; Molloy, B.; Mourão, P.A.S. Structure and anticoagulant activity of sulfated fucans. J. Biol. Chem. 1999, 274, 7656–7667. [Google Scholar] [CrossRef]
- Nishino, T.; Nagumo, T. Sugar constituents and blood-anticoagulant activities of fucose-containing sulfated polysaccharides in nine brown seaweed species. Nippon Nogeikagaku Kaishi 1987, 61, 361–363. [Google Scholar] [CrossRef]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D'Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; Usov, A.I.; Nadezhda, E.; Grachev, A.A.; Sanderson, C.J.; Kelly, M.; Rabinovich, G.A.; Iacobelli, S. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar]
- Dobashi, K.; Nishino, T.; Fujihara, M. Isolation and preliminary characterization of fucose-containing sulfated polysaccharides with blood-anticoagulant activity from seaweed Hizikia fusiforme. Carbohydr. Res. 1989, 194, 315–320. [Google Scholar] [CrossRef]
- Nishino, T.; Nagumo, T. Anticoagulant and antithrombin activities of oversulfated fucans. Carbohydr. Res. 1992, 229, 355–362. [Google Scholar] [CrossRef]
- Qiu, X.D.; Amarasekara, A.; Doctor, V. Effect of oversulfation on the chemical and biological properties of fucoidan. Carbohydrate Polymers. 2006, 63, 224–228. [Google Scholar]
- Chevolot, L.; Foucault, A.; Chauber, F. Further data on the structure of brown seaweed fucans: relationships with anticoagulant activitity. Carbohydr. Res. 1999, 319, 154–165. [Google Scholar] [CrossRef]
- Yoon, S.J.; Pyun, Y.R.; Hwang, J.K.; Mourão, P.A.S. A sulfated fucan from the brown alga Laminaria cichorioides has mainly heparin cofactor II-dependent anticoagulant activity. Carbohydr. Res. 2007, 342, 2326–2330. [Google Scholar] [CrossRef]
- Pomin, V.H.; Pereira, M.S.; Valente, A.; Tollefsen, D.M.; Pavão, M.S.G.; Mourão, P.A.S. Selective cleavage and anticoagulant activity of a sulfated fucan: stereospecific removal of a 2-sulfate ester from the polysaccharide by mild acid hydrolysis, preparation of oligosaccharides, and heparin cofactor II–dependent anticoagulant activity. Glycobiology 2005, 15, 369–381. [Google Scholar]
- Pereira, M.S.; Vilela-Silva, A.E.S.; Valente, A.; Mourão, P.A.S. A 2-sulfated, 3-linked α-L-galactan is an anticoagulant polysaccharide. Carbohydr. Res. 2002, 337, 2231–2238. [Google Scholar] [CrossRef]
- Pereira, M.S.; Melo, F.R.; Mourão, P.A.S. Is there a correlation between structure and anticoagulant action of sulfated galactans and sulfated fucans? Glycobiology 2002, 12, 573–580. [Google Scholar] [CrossRef]
- Li, B.; Rui, X.Z.; Xin, J.W. Anticoagulant activity of fucoidan from Hizikia fusiforme. Agro Food Ind. Hi-tech. 2008, 19, 22–24. [Google Scholar]
- Zheng, J.; Wang, Y.; Qian, J.J. Isolation, purification and the anticoagulant activities of fucoidan. J. Mol. Sci. 2002, 18, 109–112. [Google Scholar]
- Zoysa, M.; Nikapitiya, C.; Jeon, Y.J.; Jee, Y.; Lee, J. Anticoagulant activity of sulfated polysaccharide isolated from fermented brown seaweed Sargassum fulvellum. J. Appl. Phycol. 2008, 20, 67–74. [Google Scholar] [CrossRef]
- Frank, C.; James, B.M.; Rita, E.T. Antithrombin activity of fucoidan. J. Biol. Chem. 1989, 264, 3618–3623. [Google Scholar]
- Chen, X.; Chen, W.Z.; Zheng, G.Y. Cardiovascular Pharmacology.; People’s Medical Publishing House: Beijing, 1998. [Google Scholar]
- Kuznetsova, T.A.; Besednova, N.N.; Mamaev, A.N.; Momot, A.P.; Shevchenko, N.M.; Zvyagintseva, T.N. Anticoagulant activity of fucoidan from brown algae Fucus evanescens of the Okhotsk sea. Bull. Exp. Biol. Med. 2003, 136, 471–473. [Google Scholar] [CrossRef]
- Mourão, P.A.S. Use of sulfated fucans as anticoagulant and antithrombotic agents: future perspectives. Curr. Pharmaceut. Des. 2004, 10, 967–981. [Google Scholar] [CrossRef]
- Melo, F.R.; Pereira, M.S.; Foguel, D.; Mourão, P.A.S. Antithrombin-mediated Anticoagulant Activity of Sulfated Polysaccharides. J. Biol. Chem. 2004, 279, 20824–20835. [Google Scholar] [CrossRef]
- Cheng, Z.L.; Wang, S. Study on anticoagulant activities in vitro of fucoidan and fucoidan/collagen blends. J. Funct. Polym. 2003, 16, 557–560. [Google Scholar]
- Li, F.; Tian, T.C.; Shi, Y.C. Study on anti virus effect of fucoidan in vitro. J. N. Bethune Univ. Med. Sci. 1995, 21, 255–257. [Google Scholar]
- Hemmingson, J.A.; Falshaw, R.; Furneaux, R.H.; Thompson, K. Structure and antiviral activity of the galactofucan sulfates extracted from Undaria pinnatifida (Phaeophyta). J. Appl. Phycol. 2006, 18, 185–193. [Google Scholar] [CrossRef]
- Mandal, P.; Mateu, C.G.; Chattopadhyay, K.; Pujol, C.A.; Damonte, E.B.; Ray, B. Structural features and antiviral activity of sulphated fucans from the brown seaweed Cystoseira indica. Antivir. Chem. Chemother. 2007, 18, 153–162. [Google Scholar]
- Hayashi, K.; Nakano, T.; Hashimoto, M.; Kanekiyo, K.; Hayashi, T. Defensive effects of a fucoidan from brown alga Undaria pinnatifida against herpes simplex virus infection. Int. Immunopharmacol. 2008, 8, 109–116. [Google Scholar] [CrossRef]
- Lapshina, L.; Reunov, A.; Nagorskaya, V.; Zvyagintseva, T.; Shevchenko, N. Inhibitory effect of fucoidan from brown alga Fucus evanescens on the spread of infection induced by tobacco mosaic virus in tobacco leaves of two cultivars. Russ. J. Plant Physiol. 2006, 53, 246–251. [Google Scholar] [CrossRef]
- OoI, E.V.; Ang, P. O., Jr. Antiviral polysaccharides isolated from Hong Kong brown seaweed Hydroclathrus clathratus. Sci. China Ser. C-Life Sci. 2007, 50, 611–618. [Google Scholar] [CrossRef]
- Doh-ura, K.; Kuge, T.; Uomoto, M.; Nishizawa, K.; Kawasaki, Y.; Iha, M. Prophylactic effect of dietary seaweed fucoidan against enteral prion infection. Antimicrob. Agents Chemother. 2007, 51, 2274–2277. [Google Scholar] [CrossRef]
- Usui, T. Isolation of highly purified fucoidan from Eisenia bicyclics and its anticoagulant and antitumor activities. Agric. Biol. Chem. 1980, 44, 1965–1966. [Google Scholar] [CrossRef]
- Song, J.Q.; Xu, Y.T.; Zhang, H.K. Immunomodulation action of sulfate polysaccharide of Laminaria japonica on peritoneal macrophages of mice. Chin. J. Immunol. 2000, 16, 70–70. [Google Scholar]
- Shi, Z.Y.; Guo, Y.Z.; Wang, Z. Pharmacological activity of fucoidan from Laminaria japonic. J. Shanghai Fish. Univ. 2000, 9, 268–271. [Google Scholar]
- Aisa, Y.; Miyakawa, Y.; Nakazato, T.; Shibata, H.; Saito, K.; Ikeda, Y.; Kizaki, M. Fucoidan induces apoptosis of human HS-Sultan cells accompanied by activation of caspase-3 and down-regulation of ERK pathways. Am. J. Hematol. 2004, 78, 7–14. [Google Scholar]
- Alekseyenko, T.V.; Zhanayeva, S.Y.; Venediktova, A.A.; Zvyagintseva, T.N.; Kuznetsova, T.A.; Besednova, N.N.; Korolenko, T.A. Antitumor and antimetastatic activity of fucoidan, a sulfated polysaccharide isolated from the Okhotsk sea Fucus evanescens brown alga. Bull. Exp. Biol. Med. 2007, 143, 730–732. [Google Scholar] [CrossRef]
- Liu, J.M.; Bignon, J.; Haroun-Bouhedja, F.; Bittoun, P.; Vassy, J.; Fermandjian, S.; Wdzieczak-Bakala, J.; Boisson-Vidal, C. Inhibitory effect of fucoidan on the adhesion of adenocarcinoma cells to fibronectin. Anticancer Res. 2005, 25, 2129–2133. [Google Scholar]
- Haneji, K.; Matsuda, T.; Tomita, M.; Kawakami, H.; Ohshiro, K.; Uchihara, J.; Masuda, M.; Takasu, N.; Tanaka, Y.; Ohta, T.; Mori, N. Fucoidan extracted from Cladosiphon okamuranus Tokida induces apoptosis of human T-Cell leukemia virus type 1-infected T-Cell lines and primary adult T-Cell leukemia cells. Nutrit. Cancer 2005, 52, 189–201. [Google Scholar] [CrossRef]
- Maruyamaa, H.; Tamauchib, H.; Iizuka, M.; Nakano, T. The role of NK cells in antitumor activity of dietary fucoidan from Undaria pinnatifida Sporophylls (Mekabu). Planta Med. 2006, 72, 1415–1417. [Google Scholar] [CrossRef]
- Wang, W.T.; Zhou, J.H.; Xing, S.T.; Guan, H.S. Immunomodulating action of marine algae sulfated polysaccharides on normal and immunosuppressed mice. Chin. J. Pharm Toxicol. 1994, 8, 199–202. [Google Scholar]
- Wu, X.W.; Yang, M.L.; Huang, X.L.; Yan, J.; Luo, Q. Effect of fucoidan on splenic lymphocyte apoptosis induced by radiation. Chin. J. Radiol. Med. Prot. 2003, 23, 430–432. [Google Scholar]
- Wu, X.W.; Yang, M.L.; Huang, X.L.; Yan, J.; Luo, Q. Effect of Laminaria japonica polysaccharides on radioprotection and splenic lymphocyte apoptosis. Med. J. Wuhan Univ. 2004, 25, 239–241. [Google Scholar]
- Yang, X.L.; Sun, J.Y.; Xu, H.N. An experimental study on immunoregulatory effect of fucoidan. Chin. J. Marine Drugs 1995, 9–13. [Google Scholar]
- Shimizu, J.; Wada-Funada, U.; Mano, H.; Matahira, Y.; Kawaguchi, M.; Wada, M. Proportion of murine cytotoxic T cells is increased by high molecular-weight fucoidan extracted from Okinawa mozuku (Cladosiphon okamuranus). J. Health Sci. 2005, 51, 394–397. [Google Scholar] [CrossRef]
- Kima, M.H.; Joo, H.G. Immunostimulatory effects of fucoidan on bone marrow-derived dendritic cells. Immunol. Lett. 2008, 115, 138–143. [Google Scholar] [CrossRef]
- Choi, E.M.; Kim, A.J.; Kim, Y.; Hwang, J.K. Immunomodulating activity of arabinogalactan and fucoidan in vitro. J. Med. Food 2005, 8, 446–453. [Google Scholar] [CrossRef]
- Li, D.Y.; Xu, R.Y.; Zhou, W.Z.; Sheng, X.B.; Yang, A.Y.; Cheng, J.L. Effects of fucoidan extracted from brown seaweed on lipid peroxidation in mice. Acta Nutrim. Sin. 2002, 24, 389–392. [Google Scholar]
- Zhang, Q.B.; Yu, P.Z.; Zhou, G.F.; Li, Z.E.; Xu, Z.H. Studies on antioxidant activities of fucoidan from Laminaria japonica. Chin. Trad. Herbal Drugs 2003, 34, 824–826. [Google Scholar]
- Micheline, R.S.; Cybelle, M.; Celina, G.D.; Fernando, F.S.; Hugo, O.R.; Edda, L. Antioxidant activities of sulfated polysaccharides from brown and red seaweeds. J. Appl. Phycol. 2007, 19, 153–160. [Google Scholar] [CrossRef]
- Zhao, X.; Xue, C.H.; Cai, Y.P.; Wang, D.F.; Fang, Y. The study of antioxidant activities of fucoidan from Laminaria japonica. High Tech. Lett. 2005, 11, 91–94. [Google Scholar]
- Li, L.H.; Xue, C.H.; Xue, Y.; Li, Z.J.; Fu, X.Y. The effects of fucoidans from Laminaria japonica on AAPH mediated oxidation of human low-density lipoprotein. Acta Oceanol Sin. 2006, 25, 124–130. [Google Scholar]
- Wang, J.; Zhang, Q.; Zhang, Z.; Li, Z. Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2008, 42, 127–132. [Google Scholar] [CrossRef]
- Li, D.Y.; Xu, Z.; Zhang, S.H. Prevention and cure of fucoidan of L. japonica on mice with hypercholesterolemia. Food Sci. 1999, 20, 45–46. [Google Scholar]
- Li, D.Y.; Xu, Z.; Huang, L.M.; Wang, H.B.; Zhang, S.H. Effect of fucoidan of L. japonica on rats with hyperlipidaemia. Food Sci. 2001, 22, 92–95. [Google Scholar]
- Wang, S.Z.; Bi, A.F. Clinic observation of fucoidan on patients with hyperlipidaemia. Med. J. Qilu. 1994, 173–174. [Google Scholar]
- Li, Z.J.; Xue, C.H.; Lin, H. The hypolipidemic effects and antioxidative activity of sulfated fucan on the experimental hyperlipidemia in rats. Acta Nutrim. Sin. 1999, 21, 280–283. [Google Scholar]
- Fu, X.Y.; Xue, C.H.; Ning, Y.; Li, Z.J.; Xu, J.C. Acute antihypertensive effects of fucoidan oligosaccharides prepared from Laminaria japonica on renovascular hypertensive rat. J. Ocean Univ. Qingdao 2004, 34, 560–564. [Google Scholar]
- Blondin, C.; Fisher, E.; Boisson-Vidal, C.; Kazatchkine, M.; Jozefonvicz, J. Inhibition of complement activation by natural sulphated polysaccharides (fucans) from brown seaweed. Mol. Immunol. 1994, 31, 247–253. [Google Scholar] [CrossRef]
- Zvyagintseva, T.; Shevshenko, N.; Nazarova, I.; Scobun, A.; Luk'yanov, P.; Elyakova, L. Inhibition of complement activation by water-soluble polysaccharides of some far-eastern brown seaweeds. Comp. Biochem. Phys. C. 2000, 126, 209–215. [Google Scholar]
- Tissot, B.; Daniel, R. Biological properties of sulfated fucans: the potent inhibiting activity of algal fucoidan against the human complement system. Glycobiology 2003, 13, 29–30. [Google Scholar] [CrossRef]
- Fukuta, K.; Nakamura, T. Induction of hepatocyte growth factor by fucoidan and fucoidan-derived oligosaccharides. J. Pharm. Pharmacol. 2008, 60, 499–503. [Google Scholar] [CrossRef]
- Nardella, A.; Chaubet, F.; Boisson-Vidal, C.; Blondin, C.; Durand, P.; Jozefonvicz, J. Anticoagulant low molecular weight fucans produced by radical process and ion exchange chromatography of high molecular weight fucans extracted from the brown seaweed Ascophyllum nodosum. Carbohydr. Res. 1996, 289, 201–208. [Google Scholar] [CrossRef]
- Haroun-Bouhedja, F.; Ellouali, M.; Sinquin, C.; Boisson-Vidal, C. Relationship between sulfate groups and biological activities of fucans. Thromb. Res. 2000, 100, 453–459. [Google Scholar] [CrossRef]
- Luyt, C.E.; Meddahi-Pellé, A.; Ho-Tin-Noe, B.; Colliec-Jouault, S.; Guezennec, J.; Louedec, L.; Prats, H.; Jacob, M.P.; Osborne-Pellegrin, M.; Letourneur, D.; Michel, J.B. Low-molecular-weight fucoidan promotes therapeutic revascularization in a rat model of critical hindlimb ischemia. J. Pharmacol. Exp. Therapeut. 2003, 305, 24–30. [Google Scholar] [CrossRef]
- Nakamura, S.; Nambu, M.; Ishizuka, T.; Hattori, H.; Kanatani, Y.; Takase, B.; Kishimoto, S.; Amano, Y.; Aoki, H.; Kiyosawa, T.; Ishihara, M.; Maehara, T. Effect of controlled release of fibroblast growth factor-2 from chitosan/fucoidan micro complex-hydrogel on in vitro and in vivo vascularization. J. Biomed. Mater. Res. A. 2008, 85, 619–627. [Google Scholar]
- Logeart, D.; Prigent-Richard, S.; Jozefonvicz, J.; Letourneur, D. Fucans, sulfated polysaccharides extracted from brown seaweed, inhibit vascular smooth muscle cell proliferation, I: comparison with heparin for antiproliferative activity, binding and internalization. Eur. J. Cell Biol. 1997, 74, 376–384. [Google Scholar]
- Deux, J.F.; Meddahi-Pellé, A.; Blanche, A.F.L.; Feldman, L.J.; Colliec-Jouault, S.; Brée, F.; Boudghène, F.; Michel, J.B.; Letourneur, D. Low molecular weight fucoidan prevents neointimal hyperplasia in rabbit iliac artery in-stent restenosis model. Arterio. Thromb. Vasc. Biol. 2002, 22, 1604–1609. [Google Scholar] [CrossRef]
- Alkhatib, B.; Freguin-Bouilland, C.; Lallemand, F.; Henry, J.P.; Litzler, P.Y.; Marie, J.P.; Richard, V.; Thuillez, C.; Plissonnier, D. Low molecular weight fucan prevents transplant coronaropathy in rat cardiac allograft model. Transpl. Immunol. 2006, 16, 14–19. [Google Scholar] [CrossRef]
- Fréguin-Bouilland, C.; Alkhatib, B.; David, N.; Lallemand, F.; Henry, J.P.; Godin, M.; Thuillez, C.; Plissonnier, D. Low molecular weight fucoidan prevents neointimal hyperplasia after aortic allografting. Transplantation 2007, 83, 1234–1241. [Google Scholar] [CrossRef]
- Ronan, O.; Mark, R.; John, W.E. Fucoidan modulates the effect of transforming growth factor (TGF)-β1 on fibroblast proliferation and wound repopulation in in vitro models of dermalwound repair. Biol. Pharm. Bull. 2004, 27, 266–270. [Google Scholar] [CrossRef] [Green Version]
- Sezer, A.D.; Hatipolu, F.; Cevher, E.; Ogurtan, Z.; Bas, A.L.; Akbuga, J. Chitosan film containing fucoidan as a wound dressing for dermal burn healing: preparation and in vitro/in vivo evaluation. AAPS PharmSciTech 2007, 8, 39. [Google Scholar] [CrossRef]
- Sezer, A.D.; Hatipoglu, F.; Ogurtan, Z.; Bas, A.L.; Akbuga, J. Evaluation of fucoidan-chitosan hydrogels on superficial dermal burn healing in rabbit: An in vivo Study. J. Biotechnol. 2005, 118 (suppl.), S77–S77. [Google Scholar]
- Maruyamaa, H.; Tamauchib, H.; Hashimotoc, M.; Nakano, T. Suppression of Th2 immune responses by Mekabu fucoidan from Undaria pinnatifida Sporophylls. Int. Arch. Allergy Immunol. 2005, 137, 289–294. [Google Scholar] [CrossRef]
- Yang, J.W.; Se, Y.Y.; Soo, J.O.; Sang, K.K.; Keon, W.K. Bifunctional effects of fucoidan on the expression of inducible nitric oxide synthase. Biochem. Biophys. Res. 2006, 346, 345–350. [Google Scholar] [CrossRef]
- Shibata, H.; Kimura-Takagi, I.; Nagaoka, M.; Hashimoto, S.; Aiyama, R.; Iha, M.; Ueyama, S.; Yokokura, T. Properties of fucoidan from Cladosiphon okamuranus tokida in gastric mucosal protection. BioFactors. 2000, 11, 235–245. [Google Scholar] [CrossRef]
- Itsuko, K. Antiulcer agent and adhesion inhibitor for Helicobacter pylori. Eur. Pat. EP0645143.
- Kawamoto, H.; Miki, Y.; Kimura, T.; Tanaka, K.; Nakagawa, T.; Kawamukai, M.; Matsuda, H. Effects of fucoidan from Mozuku on human stomach cell lines. Food Sci. Technol. Res. 2006, 12, 218–222. [Google Scholar] [CrossRef]
- Saito, A.; Yoneda, M.; Yokohama, S.; Okada, M.; Haneda, M.; Nakamura, K. Fucoidan prevents concanavalin A-induced liver injury through induction of endogenous IL-10 in mice. Hepatol. Res. 2006, 35(3), 190–198. [Google Scholar]
- Kawano, N.; Egashira, Y.; Sanada, H. Effect of dietary fiber in edible seaweeds on the development of D-galactosamine-induced hepatopathy in rats. J. Nutr. Sci. Vitaminol. (Tokyo). 2007, 53, 446–450. [Google Scholar] [CrossRef]
- Kawano, N.; Egashira, Y.; Sanada, H. Effects of various kinds of edible seaweeds in diets on the development of D-galactosamine-induced hepatopathy in rats. J. Nutr. Sci. Vitaminol. (Tokyo). 2007, 53, 315–323. [Google Scholar] [CrossRef]
- Hayashi, S.; Itoh, A; Isoda, K.; Kondoh, M.; Kawase, M.; Yagi, K. Fucoidan partly prevents CCl4-induced liver fibrosis. Eur. J. Pharmacol. 2008, 580, 380–384. [Google Scholar] [CrossRef]
- Veena, C.K.; Josephine, A.; Preetha, S.P.; Varalakshmi, P.; Sundarapandiyan, R. Renal peroxidative changes mediated by oxalate: the protective role of fucoidan. Life Sci. 2006, 79, 1789–1795. [Google Scholar] [CrossRef]
- Veena, C.K.; Josephine, A.; Preetha, S.P.; Rajesh, N.G.; Varalakshmi, P. Mitochondrial dysfunction in an animal model of hyperoxaluria: a prophylactic approach with fucoidan. Eur. J. Pharmacol. 2008, 579, 330–336. [Google Scholar] [CrossRef]
- Veena, C.K.; Josephine, A.; Preetha, S.P.; Varalakshmi, P. Effect of sulphated polysaccharides on erythrocyte changes due to oxidative and nitrosative stress in experimental hyperoxaluria. Human Exp. Toxicol. 2007, 26, 923–932. [Google Scholar] [CrossRef]
- Veena, C.K.; Josephine, A.; Preetha, S.P.; Varalakshmi, P. Physico-chemical alterations of urine in experimental hyperoxaluria: a biochemical approach with fucoidan. J. Pharm. Pharmacol. 2007, 59, 419–527. [Google Scholar] [CrossRef]
- Veena, C.K.; Josephine, A.; Preetha, S.P.; Varalakshmi, P. Beneficial role of sulfated polysaccharides from edible seaweed Fucus vesiculosus in experimental hyperoxaluria. Food Chem. 2007, 100, 1552–1559. [Google Scholar] [CrossRef]
- Liu, J.C.; Zheng, F.L.; Liu, Y.P. Effect of fucoidan on renal interstitial fibrosis in adenine-induced chronic renal failure in rats. Nephrology 2008, 13 (suppl.), 158. [Google Scholar]
- Zhang, Q.B.; Li, N.; Zhao, T.T.; Qi, H.M.; Xu, Z.H.; Li, Z.E. Fucoidan inhibits the development of proteinuria in active Heymann nephritis. Phytother. Res. 2005, 19, 50–53. [Google Scholar]
© 2008 by the authors. Licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and Bioactivity. Molecules 2008, 13, 1671-1695. https://doi.org/10.3390/molecules13081671
Li B, Lu F, Wei X, Zhao R. Fucoidan: Structure and Bioactivity. Molecules. 2008; 13(8):1671-1695. https://doi.org/10.3390/molecules13081671
Chicago/Turabian StyleLi, Bo, Fei Lu, Xinjun Wei, and Ruixiang Zhao. 2008. "Fucoidan: Structure and Bioactivity" Molecules 13, no. 8: 1671-1695. https://doi.org/10.3390/molecules13081671