The Isolated and Combined Effects of Folic Acid and Synthetic Bioactive Compounds against Aβ(25-35)-Induced Toxicity in Human Microglial Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
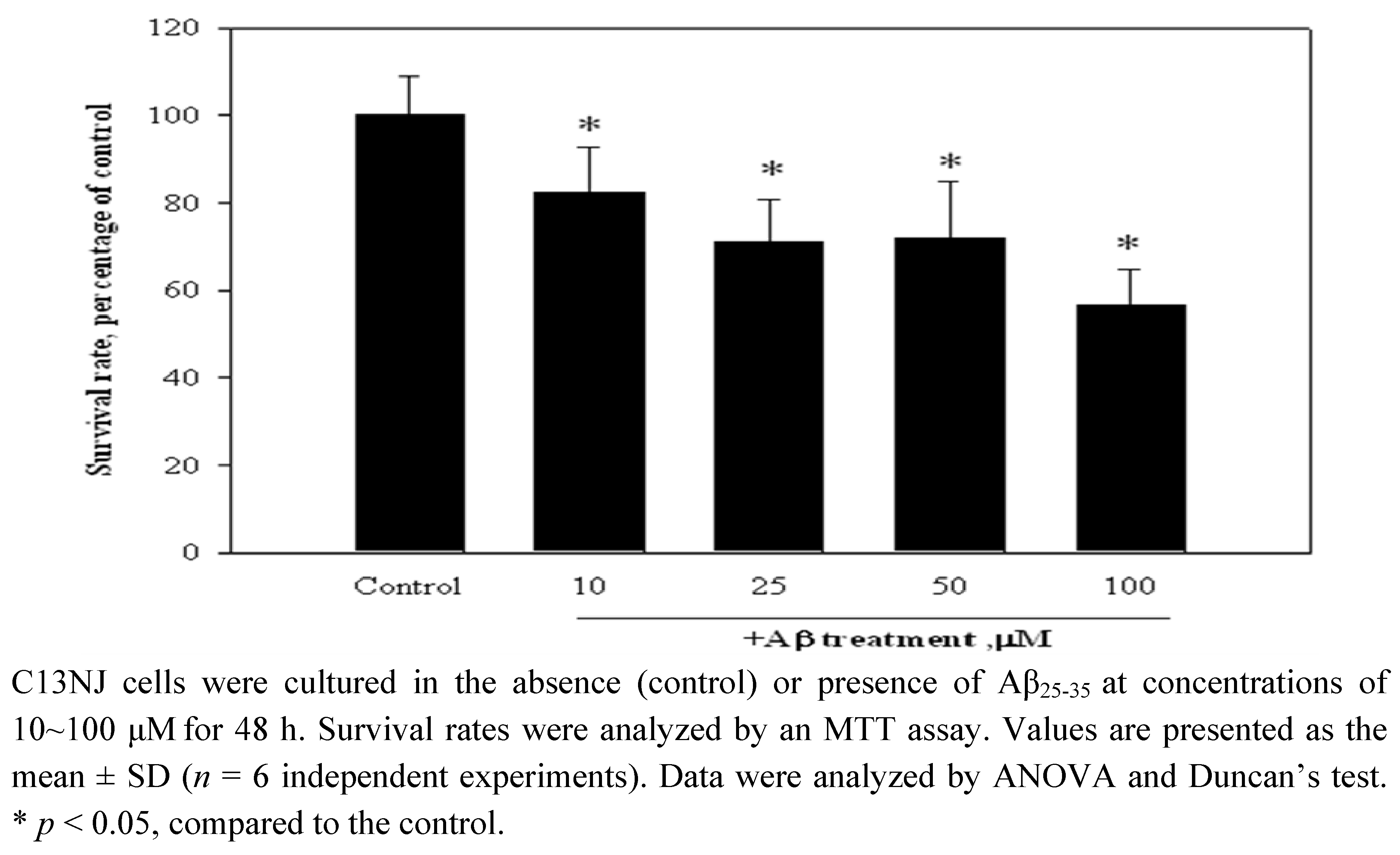
2.1. Aβ(25-35)-Induced cytotoxicity of C13NJ cells
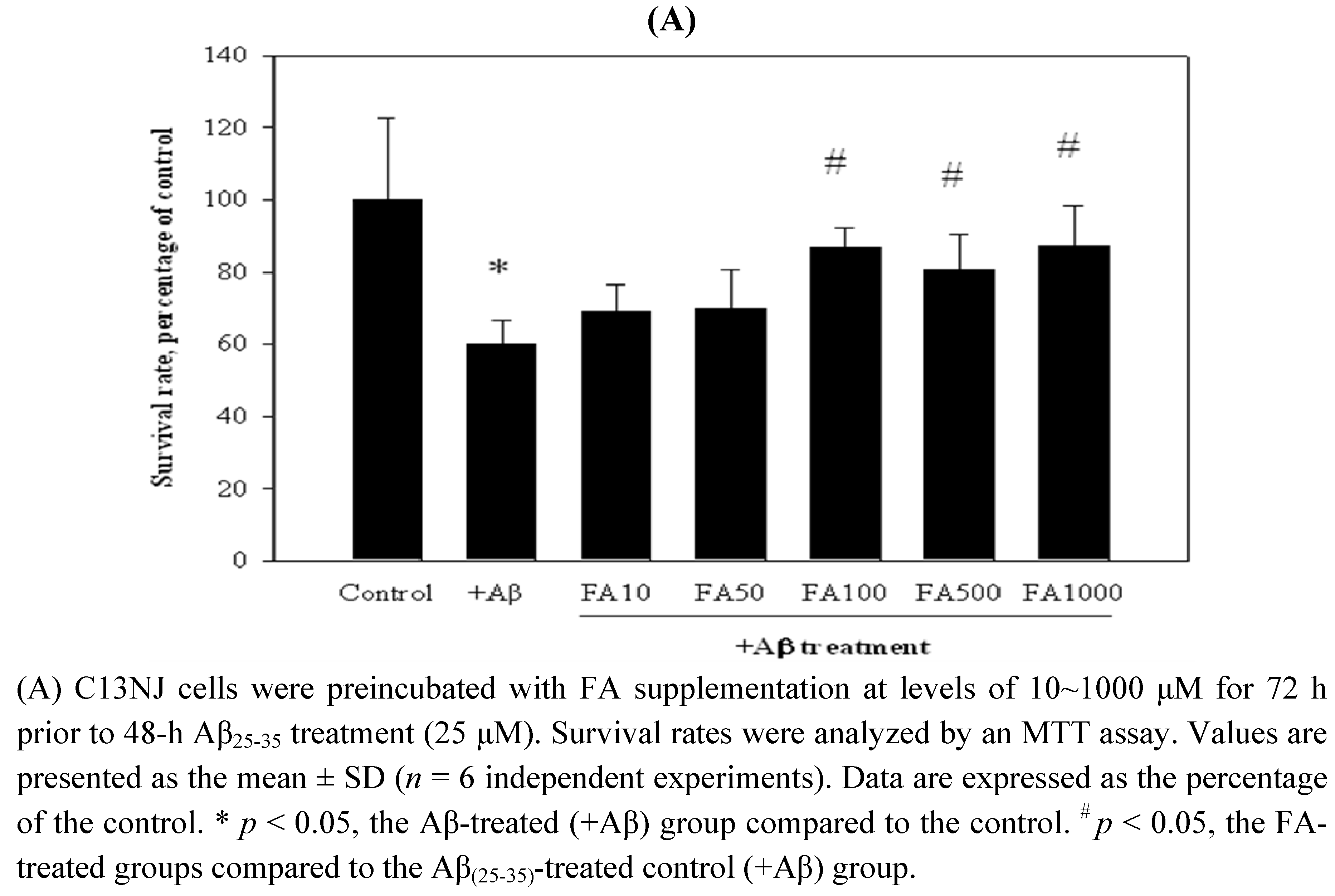
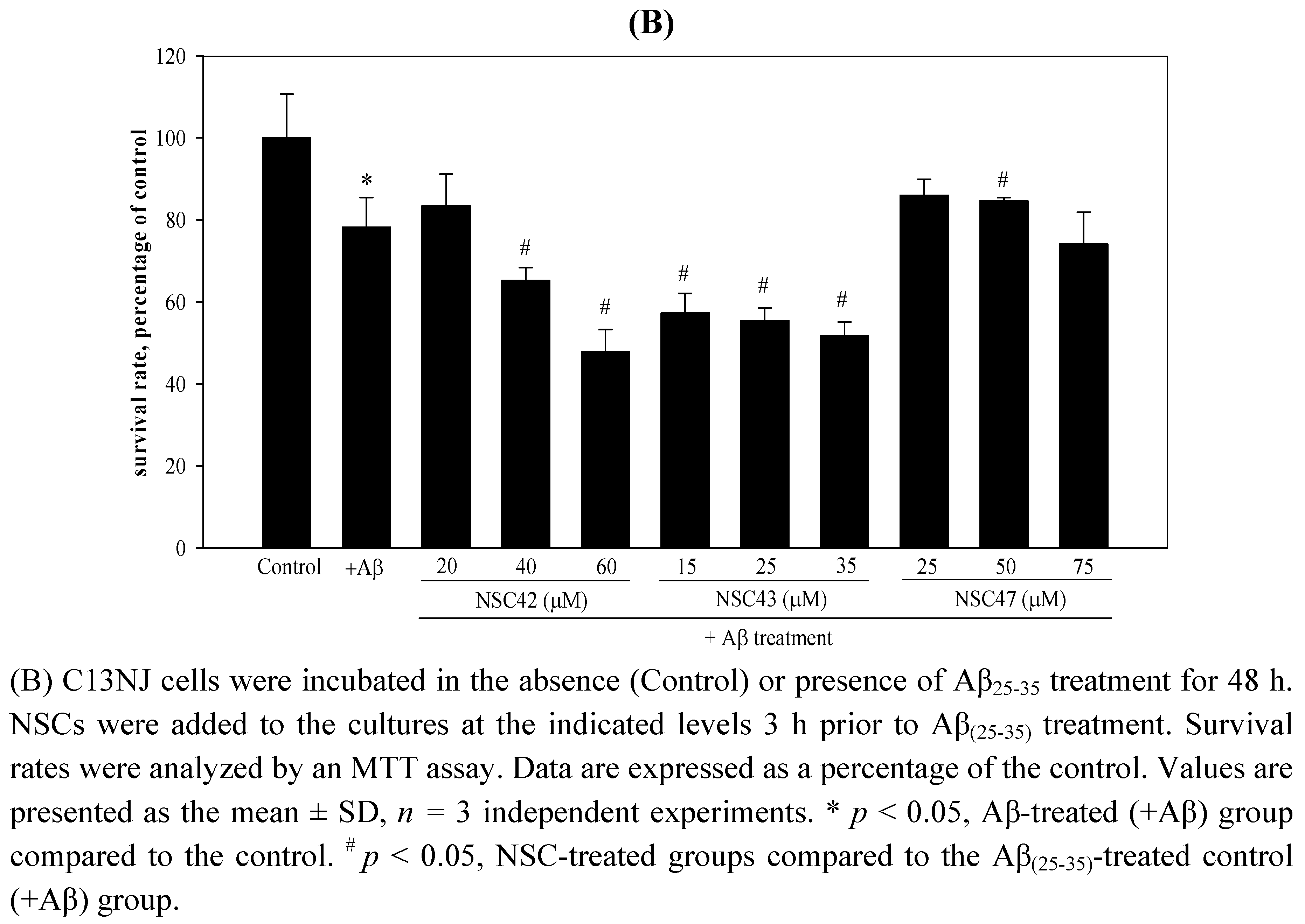
2.2. Effects of folic acid or NSCs on Aβ(25-35)-induced cytotoxicity of C13NJ cells
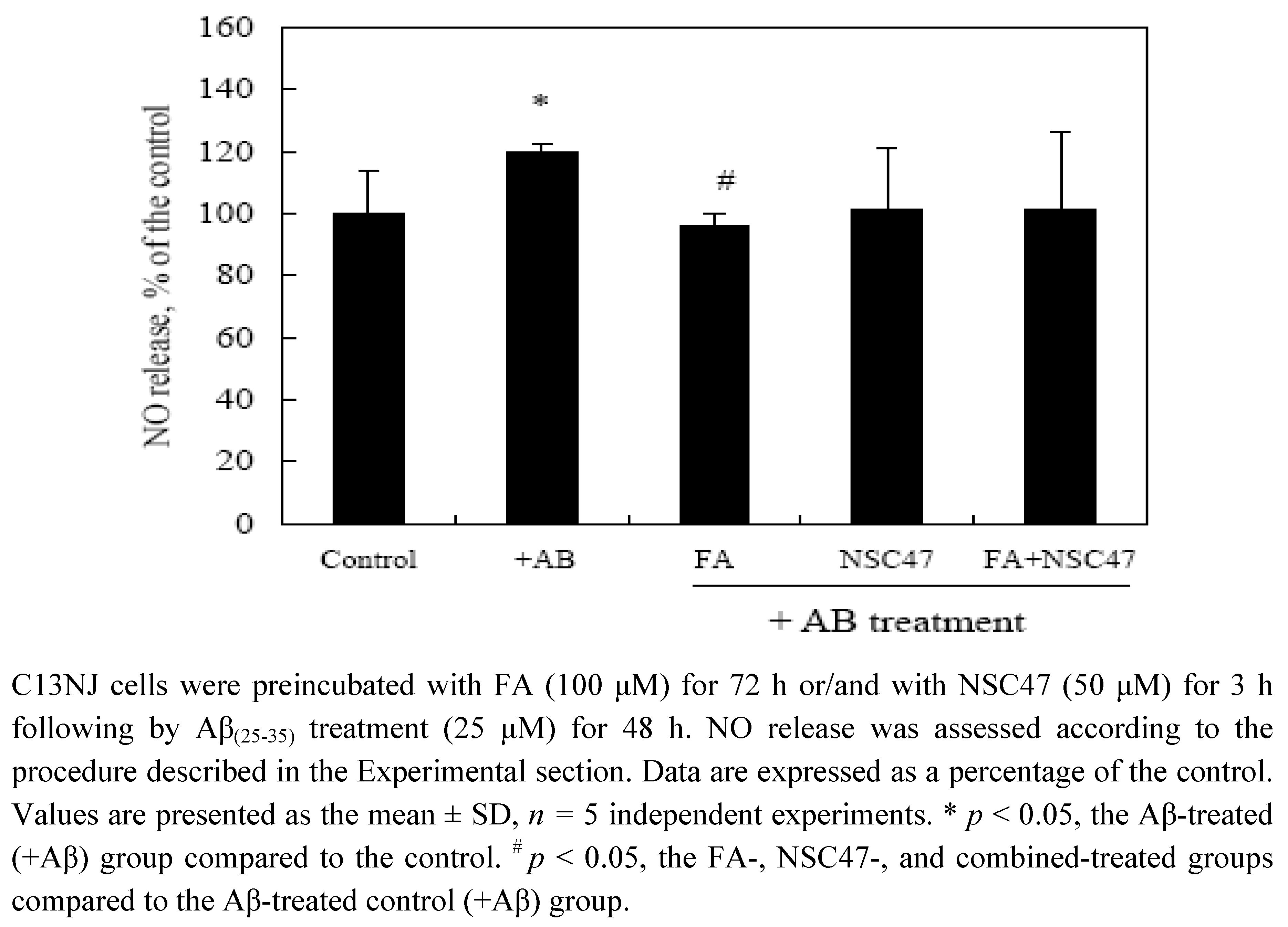
2.3. Effects of folic acid or/and NSC47 on NO production of Aβ(25-35)-treated C13NJ cells
2.4. Effects of folic acid or/and NSC47 on superoxide production of Aβ(25-35)-treated C13NJ cells
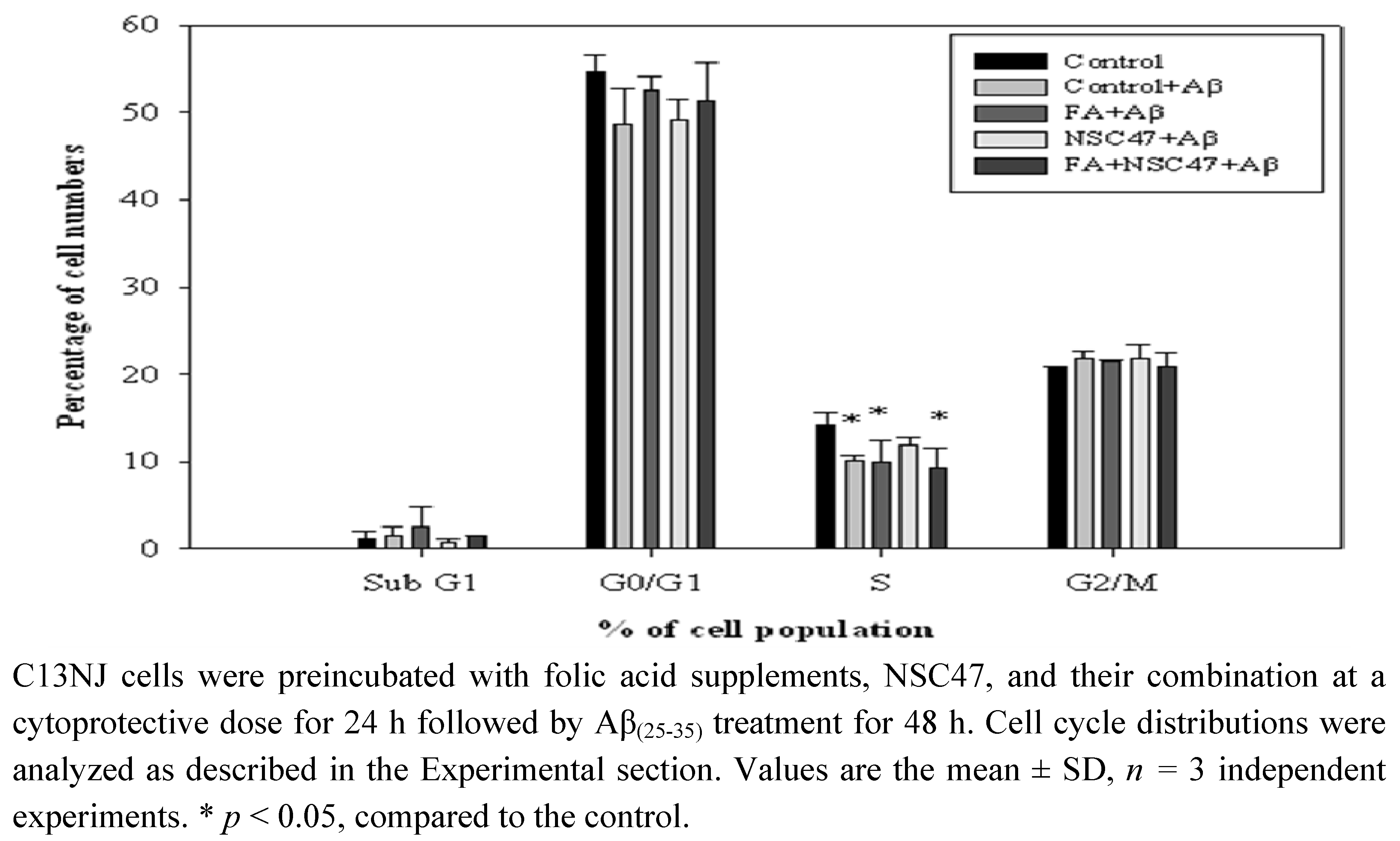
2.5. Effects of folic acid or/and NSC47 on the cell cycle of Aβ(25-35)-treated C13NJ cells
3. Discussion
4. Experimental
4.1. Preparation of NSC, folate, and the Aβ(25-35) solution
4.2. Culture of human microglial cells and experimental treatments
4.3. MTT test
4.4. NO release assay
4.5. Intracellular reactive oxygen species (ROS) levels
4.6. Cell cycle analysis
4.7. Statistical analysis
5. Conclusions
Acknowledgements
References
- Thal, D.R.; Ru, U.; Orantes, M.; Braak, H. Phases of A beta-deposition in the human brain and its relevance for the development of AD. Neurology 2002, 58, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Holscher, C.; Gengler, S.; Gault, V.A.; Harriott, P.; Mallot, H.A. Soluble betaamyloid [25–35] reversibly impairs hippocampal synaptic plasticity and spatial learning. Eur. J. Pharmacol. 2006, 561, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Rottkamp, C.A.; Nunomura, A.; Raina, A.K.; Perry, G. Oxidative stress in Alzheimer’s disease. Biochim. Biophys. Acta 2000, 1502, 139–144. [Google Scholar] [CrossRef]
- Gonzalez-Scarano, F.; Baltuch, G. Microglia as mediators of inflammatory and degenerative diseases. Annu. Rev. Neurosci. 1999, 22, 219–240. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Yamaguchi, H.; Ogawa, A.; Sugihara, S.; Nakazato, Y. Microglial activation in early stages of amyloid beta protein deposition. Acta Neuropathol. 1997, 94, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Sunamoto, M.; Ohnishi, K.; Ichimori, Y. Beta-amyloid protein-dependent nitric oxide production from microglial cells and neurotoxicity. Brain Res. 1996, 720, 93–100. [Google Scholar]
- Ard, M.D.; Cole, G.M.; Wei, J.; Mehrle, A.P.; Fratkin, J.D. Scavenging of Alzheimer’s amyloid beta-protein by microglia in culture. J. Neurosci. Res. 1996, 43, 190–202. [Google Scholar] [CrossRef]
- Reynolds, E. Vitamin B12, folic acid, and the nervous system. Lancet Neurol. 2006, 5, 949–960. [Google Scholar] [CrossRef]
- Ho, P.I.; Ashline, D.; Dhitavat, S.; Ortiz, D.; Collins, S.C.; Shea, T.B.; Rogers, E. Folate deprivation induces neurodegeneration: roles of oxidative stress and increased homocysteine. Neurobiol. Dis. 2003, 14, 32–42. [Google Scholar] [CrossRef]
- Kruman, I.I.; Kumaravel, T.S.; Lohani, A.; Pedersen, W.A.; Cutler, R.G.; Kruman, Y.; Haughey, N.; Lee, J.; Evans, M.; Mattson, M.P. Folic acid deficiency and homocysteine impair DNA repair in hippocampal neurons and sensitize them to amyloid toxicity in experimental models of Alzheimer’s disease. J. Neurosci. 2002, 22, 1752–1762. [Google Scholar] [CrossRef] [PubMed]
- Bryan, J.; Calvaresi, E.; Hughes, D. Short-term folate, vitamin B12 or vitamin B6 supplementation slightly affects memory performance but not mood in women of various ages. J. Nutr. 2002, 132, 1345–1356. [Google Scholar] [CrossRef] [PubMed]
- Luchsinger, J.A.; Tang, M.X.; Miller, J.; Green, R.; Mayeux, R. Relation of higher folate intake to lower risk of Alzheimer disease in the elderly. Arch. Neurol. 2007, 64, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.; Adhikari, S.; Patro, B.S.; Chattopadhyay, S.; Mukherjee, T. Free radical scavenging behavior of folic acid: evidence for possible antioxidant activity. Free Radic. Biol. Med. 2001, 30, 1390–1399. [Google Scholar] [CrossRef]
- Mayer, O., Jr.; Scaronimon, J.; Rosolová, H.; Hromádka, M.; Scaronubrt, I.; Vobrubová, I. The effects of folate supplementation on some coagulation parameters and oxidative status surrogates. Eur. J. Clin. Pharmacol. 2002, 58, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Doshi, S.N.; McDowell, I.F.W.; Moat, S.J.; Lang, D.; Newcombe, R.G.; Kredan, M.B.; Lewis, M.J.; Goodfellow, J. Folate improves endothelial function in coronary artery disease. An effect mediated by reduction of intracellular superoxide? Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1196–1202. [Google Scholar] [CrossRef] [PubMed]
- Chou, S.S.P.; Chen, P.W. Cycloaddition reactions of 4-sulfur-substituted dihydro-2-pyridones and 2-pyridones with conjugated dienes. Tetrahedron 2008, 64, 1879–1887. [Google Scholar] [CrossRef]
- Numata, A.; Ibuka, I. Alkaloids from Ants and Other Insects. In The Alkaloids; Brossi, A., Ed.; Academic Press: New York, NY, USA, 1987; p. 193. [Google Scholar]
- Chou, S.H.; Chou, S.S.P.; Liew, Y.F.; Leu, J.Y.; Wang, S.J.; Huang, R.F.S.; Tzeng, W.F.; Kuo, Y.C. A new synthetic compound, 2-OH, enhances interleukin-2 and interferon-γ gene expression in human peripheral blood mononuclear cells. Molecules 2009, 14, 2345–2355. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.J.; Chou, S.H.; Kuo, Y.C.; Chou, S.S.P.; Tzeng, W.F.; Leu, C.Y.; Huang, R.F.S.; Liew, Y.F. HDT-1, a new synthetic compound, inhibits glutamate release in rat cerebral cortex nerve terminals (synaptosomes). Acta Pharmacol. Sin. 2008, 29, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Michael, J.P. Indolizidine and quinolizidine alkaloids. Nat. Prod. Rep. 2008, 25, 139–165. [Google Scholar] [CrossRef] [PubMed]
- Kaltenegger, E.; Brem, B.; Mereiter, K.; Kalchhauser, H.; Kahlig, H.; Hofer, O.; Vajrodaya, S.; Greger, H. Insecticidal pyrido[1,2-a]azepine alkaloids and related derivatives from Stemona species. Phytochemistry 2003, 63, 803–816. [Google Scholar] [CrossRef]
- Hussenether, T.; Troschuetz, R. Synthesis of 5-phenyl-5,6,7,8-tetrahydro-1,6-naphthyridines and 5-phenyl-6,7,8,9-tetrahydro-5H-pyrido[3,2-c]azepines as potential D1 receptor ligands. J. Heterocycl. Chem. 2004, 41, 857–865. [Google Scholar] [CrossRef]
- Cheng, Y.-X.; Dukat, M.; Dowd, M.; Fiedler, W.; Martin, B.; Damaj, M.I.; Glennon, R.A. Synthesis and binding of 6,7,8,9-tetrahydro-5H-pyrido[3,4-d]azepine and related ring-opened analogs at central nicotinic receptors. Eur. J. Med. Chem. 1999, 34, 177–190. [Google Scholar] [CrossRef]
- Chou, S.S.P.; Ho, C.W. Synthesis and transformations of sulfur-substituted indolizidines and quinolizidines. Tetrahedron Lett. 2005, 46, 8551–8554. [Google Scholar] [CrossRef]
- Chou, S.S.P.; Liang, C.F.; Lee, T.M.; Liu, C.F. Synthesis of sulfur-substituted quinolizidines and pyrido[1,2-a]azepines by ring-closing metathesis. Tetrahedron 2007, 63, 8267–8273. [Google Scholar] [CrossRef]
- Stepanichev, M.Y.; Onufriev, M.V.; Mitrokhina, O.S.; Moiseeva, Yu.V.; Lazareva, N.A.; Victorov, I.V.; Gulyaeva, N.V. Neurochemical, behavioral, and neuromorphological effects of central administration of beta-amyloid peptide (25–35) in rat. Neurochemistry 2000, 17, 278–293. [Google Scholar]
- Barger, S.W.; Harmon, A.D. Microglial activation by Alzheimer amyloid precursor protein and modulation by apolipoprotein E. Nature 1997, 388, 878–881. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.Q.; Ye, Z.; Kholodenko, D.; Seubert, P.; Selkoe, D.J. Degradation of amyloid beta-protein by a metalloprotease secreted by microglia and other neural and non-neural cells. J. Biol. Chem. 1997, 272, 6641–6646. [Google Scholar] [CrossRef] [PubMed]
- Giulian, D.; Haverkamp, L.J.; Li, J.; Karshin, W.L.; Yu, J.; Tom, D.; Li, X.; Kirkpatrick, J.B. Senile plaques stimulate microglia to release a neurotoxin found in Alzheimer brain. Neurochem. Int. 1995, 27, 119–137. [Google Scholar] [CrossRef]
- Dorheim, M.A.; Tracey, W.R.; Pollock, J.S.; Grammas, P. Nitric oxide synthase activity is elevated in brain microvessels in Alzheimer’s disease. Biochem. Biophys. Res. Commun. 1994, 205, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, S.; Li, L.; Wang, Q.; Le, W. Folic acid protects motor neurons against the increased homocysteine, inflammation and apoptosis in SOD1G93A transgenic mice. Neuropharmacology 2008, 54, 1112–1119. [Google Scholar] [CrossRef] [PubMed]
- Ohara, Y.; Peterson, T.E.; Sayegh, H.S.; Subramanian, R.R.; Wilcox, J.N.; Harrison, D.G. Dietary correction of hypercholesterolemia in the rabbit normalizes endothelial superoxide anion production. Circulation 1995, 92, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Harrison, D.G.; Ohara, Y. Physiologic consequences of increased vascular oxidant stresses in hypercholesterolemia and atherosclerosis: Implications for impaired vasomotion. Am. J. Cardiol. 1995, 75, 75B–81B. [Google Scholar] [CrossRef]
- Vignais, P.V. The superoxide-generating NADPH oxidase: Structural aspects and activation mechanism. Cell Mol. Life Sci. 2002, 59, 1428–1459. [Google Scholar] [CrossRef] [PubMed]
- Lambeth, J.D. NOX enzymes and the biology of reactive oxygen. Nat. Rev. Immunol. 2004, 4, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Hirai, K.; Aliev, G.; Nunomura, A.; Fujioka, H.; Russell, R.L.; Atwood, C.S.; Johnson, A.B.; Kress, Y.; Vinters, H.V.; Tabaton, M.; Shimohama, S.; Cash, A.D.; Siedlak, S.L.; Harris, P.L.R.; Jones, P.K.; Petersen, R.B.; Perry, G.; Smith, M.A. Mitochondrial abnormalities in Alzheimer’s disease. J. Neurosci. 2001, 21, 3017–3023. [Google Scholar] [CrossRef] [PubMed]
- Zamzami, N.; Kroemer, G. The mitochondrion in apoptosis: How Pandora’s box opens. Nat. Rev. Mol. Cell. Biol. 2001, 2, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.F.S.; Yaong, S.C.; Chen, S.C.; Lu, Y.F. In vitro folate supplementation alleviates oxidative stress, mitochondria-associated death signalling and apoptosis induced by 7-ketocholesterol. Br. J. Nutr. 2004, 92, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.M.; Yu, C.C.; Lu, H.T.; Chou, Y.F.; Huang, R.F.S. Folate deprivation promotes mitochondrial oxidative decay: DNA large deletions, cytochrome c oxidase dysfunction, membrane depolarization and superoxide overproduction in rat liver. Br. J. Nutr. 2007, 97, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.P.; Carter, W.O.; Narayanan, P.K. Oxidative product formation analysis by flow cytometry. Meth. Cell. Biol. 1994, 41, 437–447. [Google Scholar]
Sample Availability: Samples of the compounds (NSC42 and 43) are available from the authors. |






© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liew, Y.-F.; Huang, C.-T.; Chou, S.-S.P.; Kuo, Y.-C.; Chou, S.-H.; Leu, J.-Y.; Tzeng, W.-F.; Wang, S.-J.; Tang, M.-C.; Huang, R.-F.S. The Isolated and Combined Effects of Folic Acid and Synthetic Bioactive Compounds against Aβ(25-35)-Induced Toxicity in Human Microglial Cells. Molecules 2010, 15, 1632-1644. https://doi.org/10.3390/molecules15031632
Liew Y-F, Huang C-T, Chou S-SP, Kuo Y-C, Chou S-H, Leu J-Y, Tzeng W-F, Wang S-J, Tang M-C, Huang R-FS. The Isolated and Combined Effects of Folic Acid and Synthetic Bioactive Compounds against Aβ(25-35)-Induced Toxicity in Human Microglial Cells. Molecules. 2010; 15(3):1632-1644. https://doi.org/10.3390/molecules15031632
Chicago/Turabian StyleLiew, Yih-Fong, Chao-Tzu Huang, Shang-Shing P. Chou, Yuh-Chi Kuo, Shiu-Huey Chou, Jyh-Yih Leu, Woan-Fang Tzeng, Su-Jane Wang, Ming-Chi Tang, and Rwei-Fen Syu Huang. 2010. "The Isolated and Combined Effects of Folic Acid and Synthetic Bioactive Compounds against Aβ(25-35)-Induced Toxicity in Human Microglial Cells" Molecules 15, no. 3: 1632-1644. https://doi.org/10.3390/molecules15031632