Inhibitory Effect of Indigo Naturalis on Tumor Necrosis Factor-α-Induced Vascular Cell Adhesion Molecule-1 Expression in Human Umbilical Vein Endothelial Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
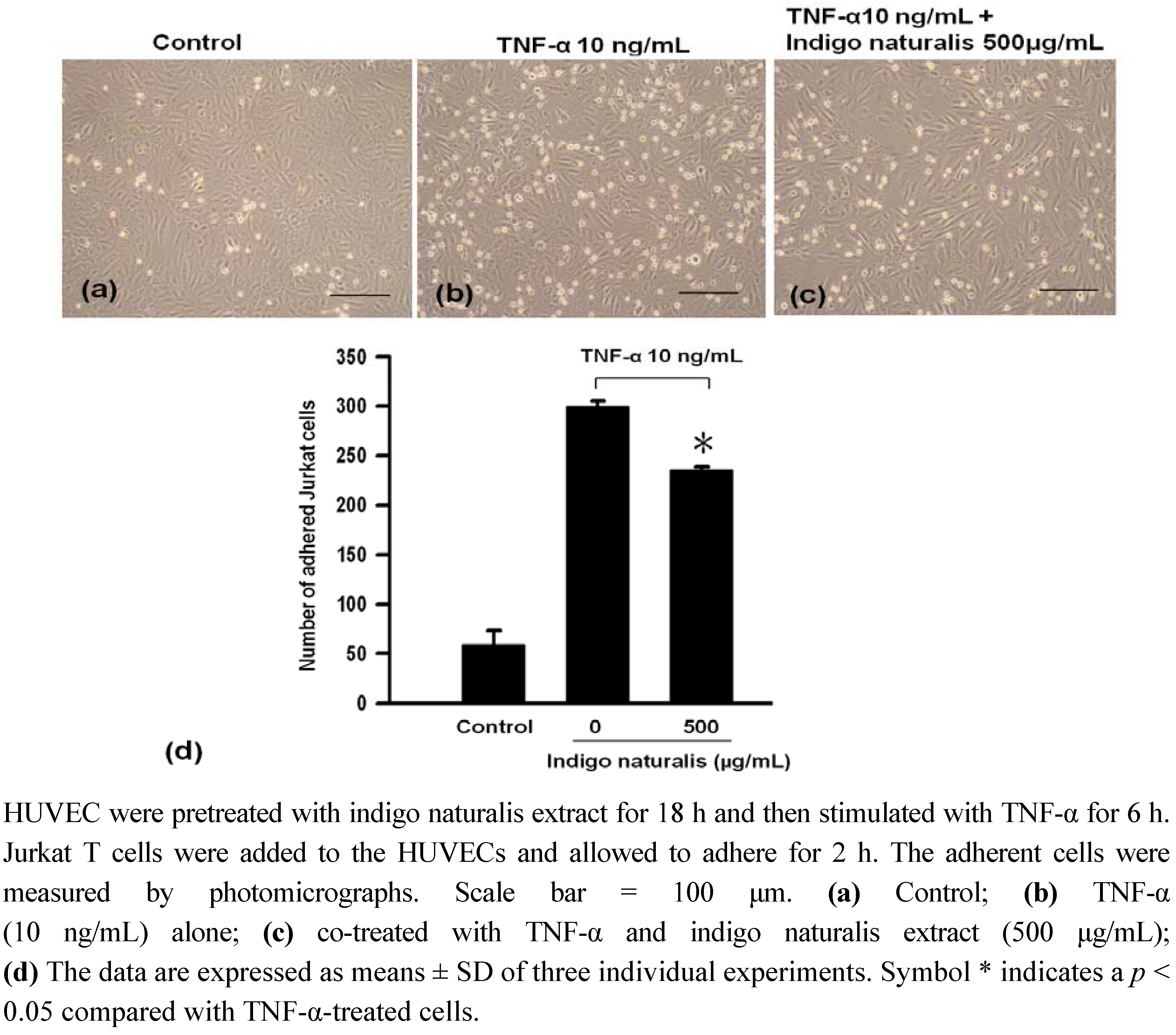
2.1. Indigo Naturalis Extract Inhibited TNF-α-induced Adhesion of Jurkat T cells to HUVECs

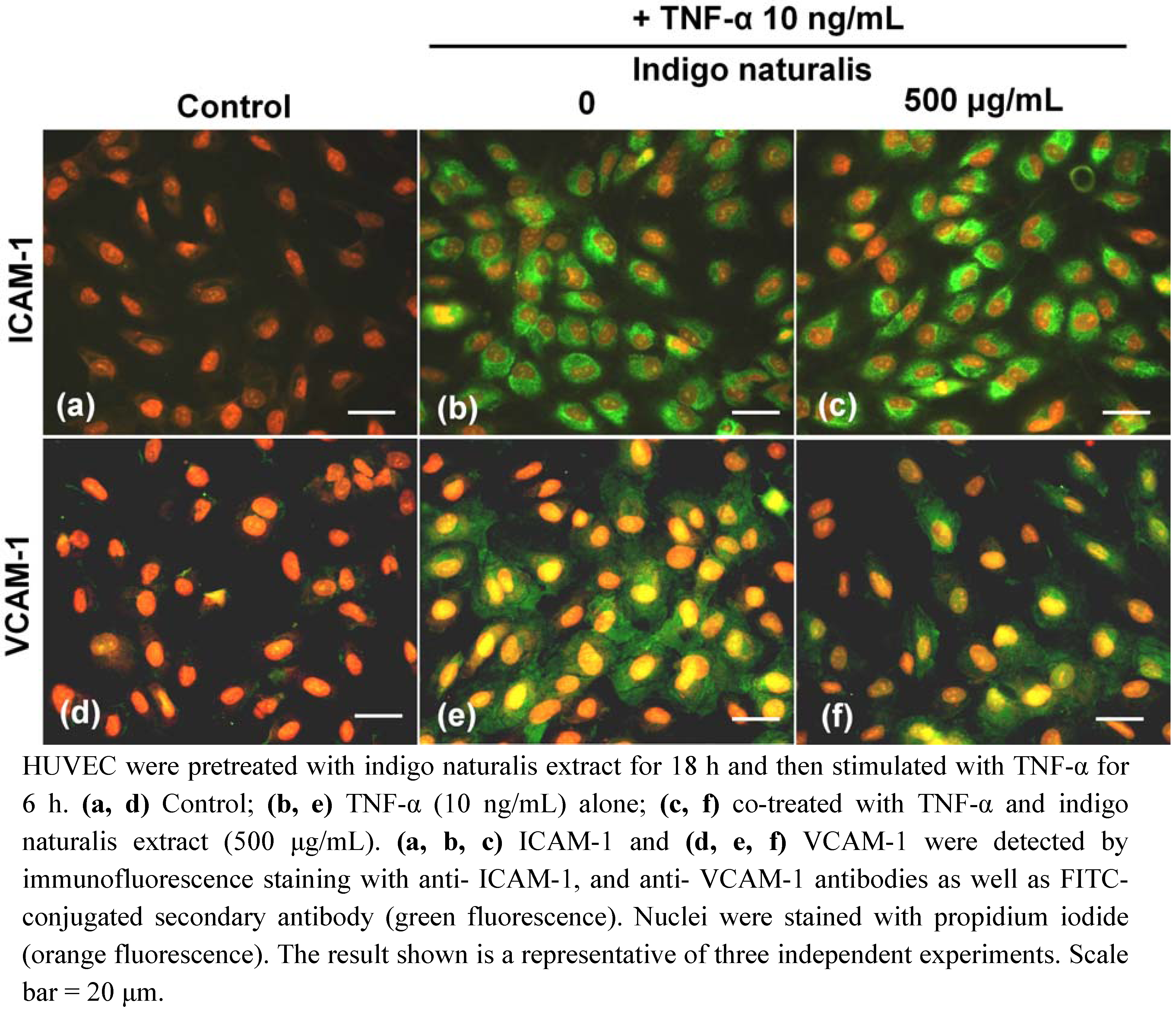
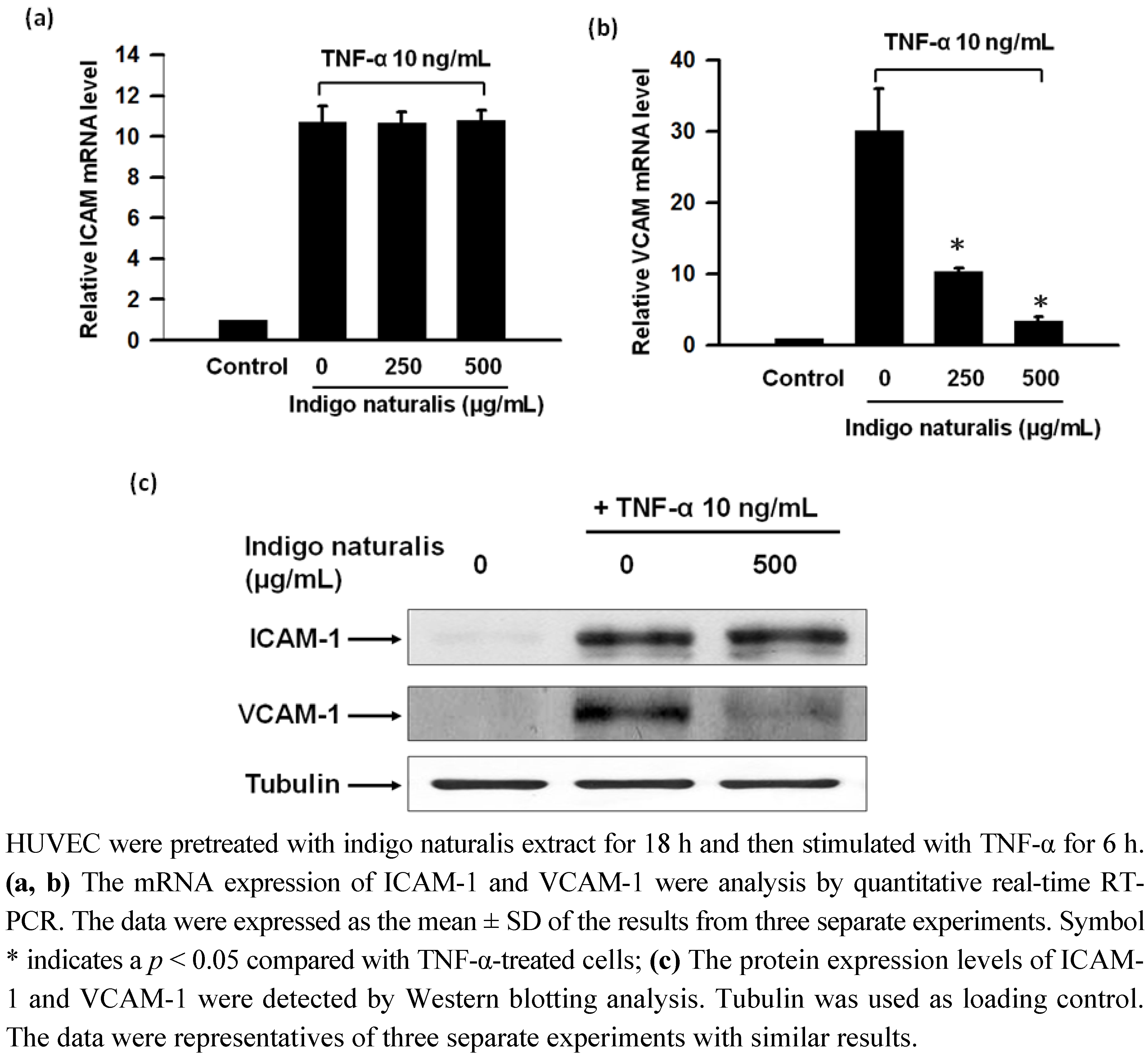
2.2. Indigo Naturalis Extract Inhibits TNF-α- induced VCAM-1 Gene Expression in HUVECs


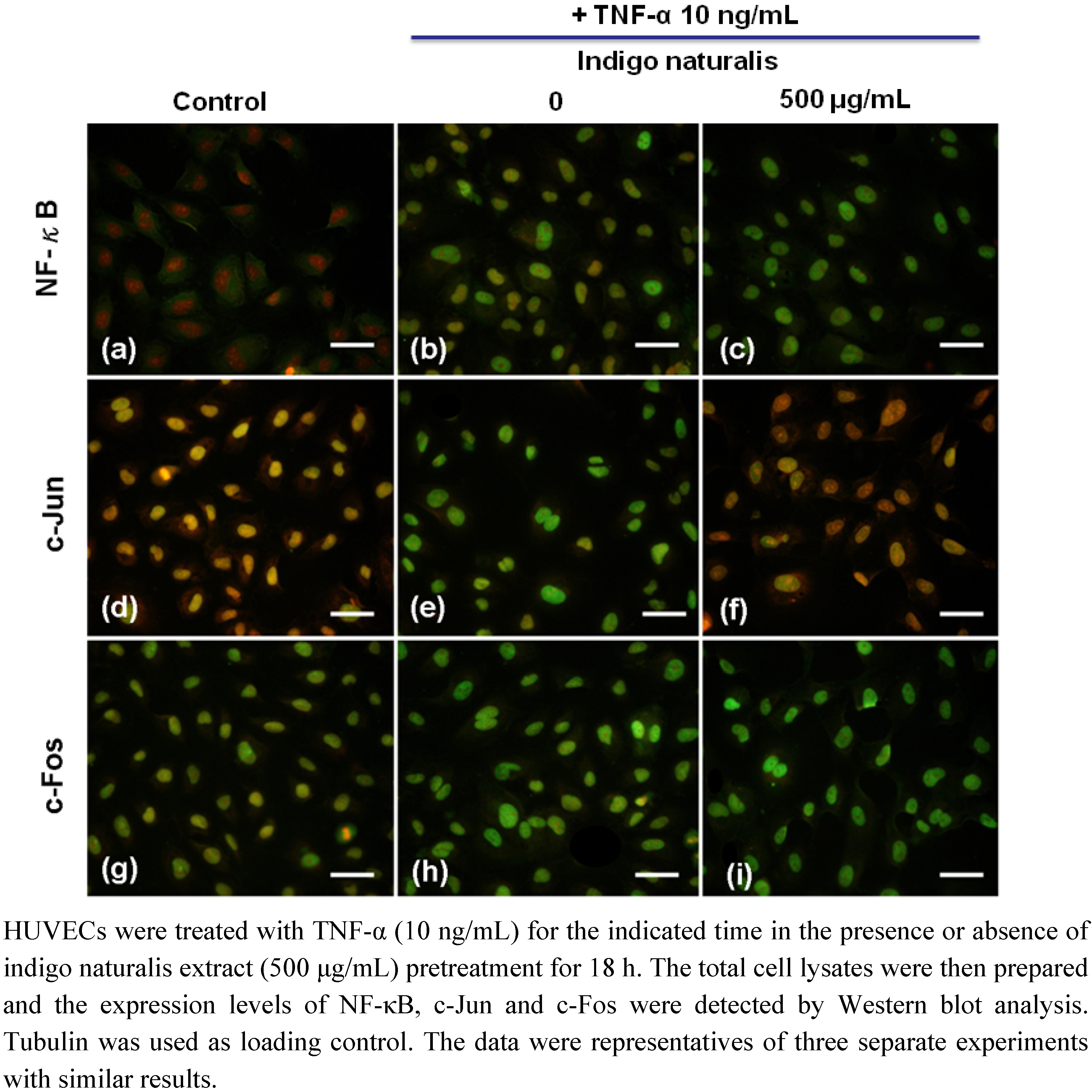
2.4. Indigo Naturalis Extract Inhibits TNF-α-induced c-Jun Expression in HUVECs


3. Experimental
3.1. Materials
3.2. Cell Culture
3.3. Cell Cytotoxicity Test
3.4. Adhesion of Jurkat T cell to HUVEC
3.5. Immunofluorescence Staining
3.6. Quantitative Real-time RT-PCR Analysis
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Acknowledgements
References and Notes
- Hedrick, M.N.; Lonsdorf, A.S.; Shirakawa, A.K.; Lee, C.C.R.; Liao, F.; Singh, S.P.; Zhang, H.H.; Grinberg, A.; Love, P.E.; Hwang, S.T.; Farber, J.M. CCR6 is required for IL-23-induced psoriasis-like inflammation in mice. J. Clin. Invest. 2009, 119, 2317–2329. [Google Scholar] [CrossRef]
- Schön, M.P.; Zollner, T.M.; Boehncke, W.H. The molecular basis of lymphocyte recruitment to the skin: clues for pathogenesis and selective therapies of inflammatory disorders. J. Invest. Dermatol. 2003, 121, 951–962. [Google Scholar] [CrossRef]
- de Boer, O.J.; Wakelkamp, I.M.; Pals, S.T.; Claessen, N.; Bos, J.D.; Das, P.K. Increased expression of adhesion receptors in both lesional and non-lesional psoriatic skin. Arch. Dermatol. Res. 1994, 286, 304–311. [Google Scholar] [CrossRef]
- Wakita, H.; Takigawa, M. E-selectin and vascular cell adhesion molecule-1 are critical for initial trafficking of helper-inducer/memory T cells in psoriatic plaques. Arch. Dermatol. 1994, 130, 457–463. [Google Scholar] [CrossRef]
- Terajima, S.; Higaki, M.; Igarashi, Y.; Nogita, T.; Kawashima, M. An important role of tumor necrosis factor-alpha in the induction of adhesion molecules in psoriasis. Arch. Dermatol. Res. 1998, 290, 246–452. [Google Scholar] [CrossRef]
- Aggarwal, B.B. Signalling pathways of the TNF superfamily: a double-edged sword. Nature Rev. Immunol. 2003, 3, 745–756. [Google Scholar] [CrossRef]
- Zhou, Z.; Gengaro, P.; Wang, W.; Wang, X.; Li, C.; Faubel, S.; Rivard, C.; Schrier, R.W. Role of NF-kappaB and PI 3-kinase/Akt in TNF-alpha-induced cytotoxicity in microvascular endothelial cells. Am. J. Physiol. Renal. Physiol. 2008, 295, F932–F941. [Google Scholar] [CrossRef]
- Lowes, M.A.; Bowcock, A.M.; Krueger, J.G. Pathogenesis and therapy of psoriasis. Nature 2007, 445, 866–873. [Google Scholar] [CrossRef]
- Koo, J.; Arain, S. Traditional Chinese medicine for the treatment of dermatologic disorders. Arch. Dermatol. 1998, 134, 1388–1393. [Google Scholar] [CrossRef]
- Lin, Y.K.; Yen, H.R.; Wong, W.R.; Yang, S.H.; Pang, J.H. Successful treatment of pediatric psoriasis with Indigo naturalis composite ointment. Pediatr. Dermatol. 2006, 23, 507–510. [Google Scholar] [CrossRef]
- Lin, Y.K.; Wong, W.R.; Su Pang, J.H. Successful treatment of recalcitrant psoriasis with Indigo naturalis ointment. Clin. Exp. Dermatol. 2007, 32, 99–100. [Google Scholar]
- Xiao, Z.; Hao, Y.; Liu, B.; Qian, L. Indirubin and meisoindigo in the treatment of chronic myelogenous leukemia in China. Leuk. Lymphoma. 2002, 43, 1763–1768. [Google Scholar] [CrossRef]
- Yuan, Z.Z.; Yuan, X.; Xu, Z.X. Studies on tabellae indigo naturalis in treatment of psoriasis. J. Tradit. Chin. Med. 1982, 2, 306. [Google Scholar]
- Lin, Y.K.; Wong, W.R.; Chang, Y.C.; Chang, C.J.; Tsay, P.K.; Chang, S.C.; Pang, J.H.S. The efficacy and safety of topically applied indigo naturalis ointment in patients with plaque-type psoriasis. Dermatology 2007, 214, 155–161. [Google Scholar]
- Lin, Y.K.; Leu, Y.L.; Yang, S.H.; Chen, H.W.; Wang, C.T.; Pang, J.H.S. Anti-psoriatic effects of indigo naturalis on the proliferation and differentiation of keratinocytes with indirubin as the active component. J. Dermatol. Sci. 2009, 54, 168–174. [Google Scholar] [CrossRef]
- Lin, Y.K.; Chang, C.J.; Chang, Y.C.; Wong, W.R.; Chang, S.C.; Pang, J.H.S. Clinical assessment of patients with recalcitrant psoriasis in a randomized, observer-blind, vehicle-controlled trial using indigo naturalis. Arch. Dermatol. 2008, 144, 1457–1464. [Google Scholar] [CrossRef]
- Lin, Y.K.; Leu, Y.L.; Huang, T.H.; Wu, Y.H.; Chung, P.J.; Su Pang, J.H.; Hwang, T.L. Anti-inflammatory effects of the extract of indigo naturalis in human neutrophils. J. Ethnopharmacol. 2009, 125, 51–58. [Google Scholar] [CrossRef]
- Torres-Alvarez, B.; Castanedo-Cazares, J.P.; Fuentes-Ahumada, C.; Moncada, B. The effect of methotrexate on the expression of cell adhesion molecules and activation molecule CD69 in psoriasis. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 334–339. [Google Scholar] [CrossRef]
- Cabrijan, L.; Lipozencić, J.; Batinac, T.; Lenković, M.; Stanić Zgombić, Z. Influence of PUVA and UVB radiation on expression of ICAM-1 and VCAM-1 molecules in psoriasis vulgaris. Coll. Antropol. 2008, 32 Suppl. 2, 53–56. [Google Scholar]
- Yamasaki, E.; Soma, Y.; Kawa, Y.; Mizoguchi, M. Methotrexate inhibits proliferation and regulation of the expression of intercellular adhesion molecule-1 and vascular cell adhesion molecule-1 by cultured human umbilical vein endothelial cells. Br. J. Dermatol. 2003, 149, 30–38. [Google Scholar]
- Gille, J.; Paxton, L.L.; Lawley, T.J.; Caughman, S.W.; Swerlick, R.A. Retinoic acid inhibits the regulated expression of vascular cell adhesion molecule-1 by cultured dermal microvascular endothelial cells. J. Clin. Invest. 1997, 99, 492–500. [Google Scholar] [CrossRef]
- Sen, C.K.; Bagchi, D. Regulation of inducible adhesion molecule expression in human endothelial cells by grape seed proanthocyanidin extract. Mol. Cell. Biochem. 2001, 216, 1–7. [Google Scholar] [CrossRef]
- Arbiser, J.L.; Govindarajan, B.; Battle, T.E.; Lynch, R.; Frank, D.A.; Ushio-Fukai, M.; Perry, B.N.; Stern, D.F.; Bowden, G.T.; Liu, A.; Klein, E.; Kolodziejski, P.J.; Eissa, N.T.; Hossain, C.F.; Nagle, D.G. Carbazole is a naturally occurring inhibitor of angiogenesis and inflammation isolated from antipsoriatic coal tar. J. Invest. Dermatol. 2006, 126, 1396–1402. [Google Scholar]
- Ahmad, M.; Theofanidis, P.; Medford, R.M. Role of activating protein-1 in the regulation of the vascular cell adhesion molecule-1 gene expression by tumor necrosis factor-alpha. J. Biol. Chem. 1998, 273, 4616–4621. [Google Scholar]
- Zenz, R.; Eferl, R.; Scheinecker, C.; Redlich, K.; Smolen, J.; Schonthaler, H.B.; Kenner, L.; Tschachler, E.; Wagner, E.F. Activator protein 1 (Fos/Jun) functions in inflammatory bone and skin disease. Arthritis Res. Ther. 2008, 10, 201. [Google Scholar]
- Mehic, D.; Bakiri, L.; Ghannadan, M.; Wagner, E.F.; Tschachler, E. Fos and jun proteins are specifically expressed during differentiation of human keratinocytes. J. Inves.t Dermatol. 2005, 124, 212–220. [Google Scholar] [CrossRef]
- Zenz, R.; Eferl, R.; Kenner, L.; Florin, L.; Hummerich, L.; Mehic, D.; Scheuch, H.; Angel, P.; Tschachler, E.; Wagner, E. F. Psoriasis-like skin disease and arthritis caused by inducible epidermal deletion of Jun proteins. Nature 2005, 437, 369–375. [Google Scholar]
- Bedi, M. K.; Shenefelt, P. D. Herbal therapy in dermatology. Arch. Dermatol. 2002, 138, 232–242. [Google Scholar] [CrossRef]
- Keith, C.T.; Borisy, A.A.; Stockwell, B.R. Multicomponent therapeutics for networked systems. Nat. Rev. Drug Discov. 2005, 4(1), 71–78. [Google Scholar] [CrossRef]
- Kong, D.X.; Li, X.J.; Zhang, H.Y. Where is the hope for drug discovery? Let history tell the future. Drug Discov. Today 2009, 14, 115–119. [Google Scholar] [CrossRef]
- Sample Availability: Indigo naturalis is available from the authors.
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chang, H.-N.; Pang, J.-H.S.; Yang, S.-H.; Hung, C.-F.; Chiang, C.-H.; Lin, T.-Y.; Lin, Y.-K. Inhibitory Effect of Indigo Naturalis on Tumor Necrosis Factor-α-Induced Vascular Cell Adhesion Molecule-1 Expression in Human Umbilical Vein Endothelial Cells. Molecules 2010, 15, 6423-6435. https://doi.org/10.3390/molecules15096423
Chang H-N, Pang J-HS, Yang S-H, Hung C-F, Chiang C-H, Lin T-Y, Lin Y-K. Inhibitory Effect of Indigo Naturalis on Tumor Necrosis Factor-α-Induced Vascular Cell Adhesion Molecule-1 Expression in Human Umbilical Vein Endothelial Cells. Molecules. 2010; 15(9):6423-6435. https://doi.org/10.3390/molecules15096423
Chicago/Turabian StyleChang, Hsin-Ning, Jong-Hwei Su Pang, Sien-Hung Yang, Chi-Feng Hung, Chi-Hsin Chiang, Tung-Yi Lin, and Yin-Ku Lin. 2010. "Inhibitory Effect of Indigo Naturalis on Tumor Necrosis Factor-α-Induced Vascular Cell Adhesion Molecule-1 Expression in Human Umbilical Vein Endothelial Cells" Molecules 15, no. 9: 6423-6435. https://doi.org/10.3390/molecules15096423