Down-Regulation of Treg Cells and Up-Regulation of Th1/Th2 Cytokine Ratio Were Induced by Polysaccharide from Radix Glycyrrhizae in H22 Hepatocarcinoma Bearing Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of GP on Tumor Growth
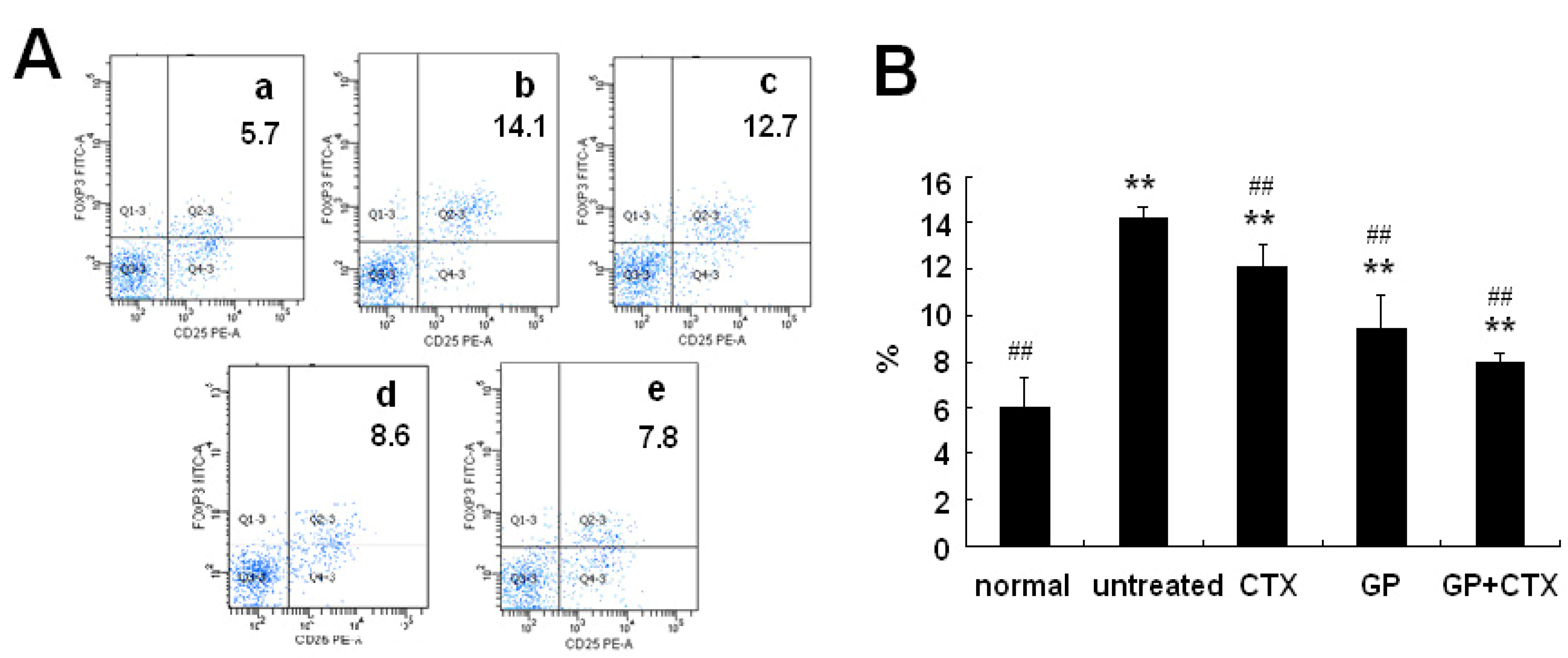
2.2. Effect of GP on Treg Cells in the Spleen
2.3. Effect of GP on Treg Cells in the Lymph Node


2.4. Effects of GP on mRNA Expression of Foxp3 and IL-10 in the Lymph Nodes

2.5. Effects of GP on the Production of Th1/Th2 Cytokines in Serum
3. Discussion

4. Experimental
4.1. Animals and Cell Lines
4.2. Drugs
4.3. Animal Treatment
4.4. Flow Cytometric Analysis
4.5. Real-Time PCR Analysis
4.6. Cytokine Assays
4.7. Statistical Analysis
5. Conclusions
Acknowledgements
Conflict of Interest
- Samples Availability: Samples of the compounds are available from the authors.
References
- Kwon, H.J.; Kim, H.H.; Ryu, Y.B.; Kim, J.H.; Jeong, H.J.; Lee, S.W.; Chang, J.S.; Cho, K.O.; Rho, M.C.; Park, S.J.; Lee, W.S. In vitro anti-rotavirus activity of polyphenol compounds isolated from the roots of Glycyrrhiza uralensis. Bioorg. Med. Chem. 2010, 18, 7668–7674. [Google Scholar]
- Li, J.; Tu, Y.; Tong, L.; Zhang, W.; Zheng, J.; Wei, Q. Immunosuppressive activity on the murine immune responses of glycyrol from Glycyrrhiza uralensis via inhibition of calcineurin activity. Pharm. Biol. 2010, 48, 1177–1184. [Google Scholar]
- Cheng, A.; Wan, F.; Wang, J.; Jin, Z.; Xu, X. Macrophage immunomodulatory activity of polysaccharides isolated from Glycyrrhiza uralensis Fish. Int. Immunopharmacol. 2008, 8, 43–50. [Google Scholar] [CrossRef]
- Cheng, A.; Wan, F.; Jin, Z.; Wang, J.; Xu, X. Nitrite oxide and inducible nitric oxide synthase were regulated by polysaccharides isolated from Glycyrrhiza uralensis Fisch. J. Ethnopharmacol. 2008, 118, 59–64. [Google Scholar] [CrossRef]
- Hong, Y.K.; Wu, H.T.; Ma, T.; Liu, W.J.; He, X.J. Effects of Glycyrrhiza glabra polysaccharides on immune and antioxidant activities in high-fat mice. Int. J. Biol. Macromol. 2009, 45, 61–64. [Google Scholar] [CrossRef]
- Beyer, M.; Schultze, J.L. Regulatory T cells in cancer. Blood 2006, 108, 804–811. [Google Scholar] [CrossRef]
- Ménétrier-Caux, C.; Gobert, M.; Caux, C. Differences in tumor regulatory T-cell localization and activation status impact patient outcome. Cancer Res. 2009, 69, 7895–7898. [Google Scholar] [CrossRef]
- Petersen, R.P.; Campa, M.J.; Sperlazza, J.; Conlon, D.; Joshi, M.B.; Harpole, D.H.; Patz, E.F. Tumor infiltrating Foxp3+ regulatory T-cells are associated with recurrence in pathologic stage I NSCLC patients. Cancer 2006, 107, 2866–2872. [Google Scholar]
- Miller, A.M.; Lundberg, K.; Ozenci, V.; Banham, A.H.; Hellström, M.; Egevad, L.; Pisa, P. CD4+CD25high T cells are enriched in the tumor and peripheral blood of prostate cancer patients. J. Immunol. 2006, 177, 7398–7405. [Google Scholar]
- Zou, W. Regulatory T cells, tumour immunity and immunotherapy. Nat. Rev. Immunol. 2006, 6, 295–307. [Google Scholar] [CrossRef]
- Valzasina, B.; Piconese, S.; Guiducci, C.; Colombo, M.P. Tumor-induced expansion of regulatory T cells by conversion of CD4+CD25- lymphocytes is thymus and proliferation independent. Cancer Res. 2006, 66, 4488–4495. [Google Scholar]
- Wittschier, N.; Faller, G.; Hensel, A. Aqueous extracts and polysaccharides from liquorice roots (Glycyrrhiza glabra L.) inhibit adhesion of Helicobacter pylori to human gastric mucosa. J. Ethnopharmacol. 2009, 125, 218–223. [Google Scholar] [CrossRef]
- Curiel, T.J.; Coukos, G.; Zou, L.; Alvarez, X.; Cheng, P.; Mottram, P.; Evdemon-Hogan, M.; Conejo-Garcia, J.R.; Zhang, L.; Burow, M.; Zhu, Y.; Wei, S.; Kryczek, I.; Daniel, B.; Gordon, A.; Myers, L.; Lackner, A.; Disis, M.L.; Knutson, K.L.; Chen, L.; Zou, W. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat. Med. 2004, 10, 942–949. [Google Scholar]
- Kryczek, I.; Zou, L.; Rodriguez, P.; Zhu, G.; Wei, S.; Mottram, P.; Brumlik, M.; Cheng, P.; Curiel, T.; Myers, L.; Lackner, A.; Alvarez, X.; Ochoa, A.; Chen, L.; Zou, W. B7-H4 expression identifies a novel suppressive macrophage population in human ovarian carcinoma. J. Exp. Med. 2006, 203, 871–881. [Google Scholar]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar]
- Torgerson, T.R.; Ochs, H.D. Immune dysregulation, polyendocrinopathy, enteropathy, X-linked: Forkhead box protein 3 mutations and lack of regulatory T cells. J. Allergy Clin. Immunol. 2007, 120, 744–750. [Google Scholar] [CrossRef]
- Gavin, M.A.; Rasmussen, J.P.; Fontenot, J.D.; Vasta, V.; Manganiello, V.C.; Beavo, J.A.; Rudensky, A.Y. Foxp3-dependent programme of regulatory T-cell differentiation. Nature 2007, 445, 771–775. [Google Scholar] [CrossRef]
- Campbell, D.J.; Ziegler, S.F. FOXP3 modifies the phenotypic and functional properties of regulatory T cells. Nat. Rev. Immunol. 2007, 7, 305–310. [Google Scholar] [CrossRef]
- Thornton, A.M.; Shevach, E.M. CD4+CD25+ immunoregulatory T cells suppress polyclonal T cell activation in vitro by inhibiting interleukin 2 production. J. Exp. Med. 1998, 188, 287–296. [Google Scholar] [CrossRef]
- Miyara, M.; Sakaguchi, S. Natural regulatory T cells: Mechanisms of suppression. Trends Mol. Med. 2007, 13, 108–116. [Google Scholar] [CrossRef]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef]
- Patel, P.; Varghese, E.; Ding, G.; Fan, S.; Kapasi, A.; Reddy, K.; Franki, N.; Nahar, N.; Singhal, P. Transforming growth factor beta induces mesangial cell apoptosis through NO- and p53-dependent and -independent pathways. J. Invest. Med. 2000, 48, 403–410. [Google Scholar]
- Hanahan, D. Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Mukamoto, M.; Kodama, H. Regulation of early chicken thymocyte proliferation by transforming growth factor-beta from thymic stromal cells and thymocytes. Vet. Immunol. Immunopathol. 2000, 77, 121–132. [Google Scholar] [CrossRef]
- Nakamura, K.; Kitani, A.; Strober, W. Cell contact-dependent immunosuppression by CD4(+)CD25(+) regulatory T cells is mediated by cell surface-bound transforming growth factor beta. J. Exp. Med. 2001, 194, 629–644. [Google Scholar] [CrossRef]
- Nakamura, K.; Kitani, A.; Fuss, I.; Pedersen, A.; Harada, N.; Nawata, H.; Strober, W. TGF-beta 1 plays an important role in the mechanism of CD4+CD25+ regulatory T cell activity in both humans and mice. J. Immunol. 2004, 172, 834–842. [Google Scholar]
- Chen, W.; Wahl, S.M. TGF-beta: the missing link in CD4+CD25+ regulatory T cell-mediated immunosuppression. Cytokine Growth Factor Rev. 2003, 14, 85–89. [Google Scholar] [CrossRef]
- Annunziato, F.; Cosmi, L.; Liotta, F.; Lazzeri, E.; Manetti, R.; Vanini, V.; Romagnani, P.; Maggi, E.; Romagnani, S. Phenotype, localization, and mechanism of suppression of CD4(+)CD25(+) human thymocytes. J. Exp. Med. 2002, 196, 379–387. [Google Scholar] [CrossRef]
- Zhang, X.; Izikson, L.; Liu, L.; Weiner, H.L. Activation of CD25(+)CD4(+) regulatory T cells by oral antigen administration. J. Immunol. 2001, 167, 4245–4253. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
He, X.; Li, X.; Liu, B.; Xu, L.; Zhao, H.; Lu, A. Down-Regulation of Treg Cells and Up-Regulation of Th1/Th2 Cytokine Ratio Were Induced by Polysaccharide from Radix Glycyrrhizae in H22 Hepatocarcinoma Bearing Mice. Molecules 2011, 16, 8343-8352. https://doi.org/10.3390/molecules16108343
He X, Li X, Liu B, Xu L, Zhao H, Lu A. Down-Regulation of Treg Cells and Up-Regulation of Th1/Th2 Cytokine Ratio Were Induced by Polysaccharide from Radix Glycyrrhizae in H22 Hepatocarcinoma Bearing Mice. Molecules. 2011; 16(10):8343-8352. https://doi.org/10.3390/molecules16108343
Chicago/Turabian StyleHe, Xiaojuan, Xiaobing Li, Biao Liu, Li Xu, Hongyan Zhao, and Aiping Lu. 2011. "Down-Regulation of Treg Cells and Up-Regulation of Th1/Th2 Cytokine Ratio Were Induced by Polysaccharide from Radix Glycyrrhizae in H22 Hepatocarcinoma Bearing Mice" Molecules 16, no. 10: 8343-8352. https://doi.org/10.3390/molecules16108343