Pentamethylquercetin Improves Adiponectin Expression in Differentiated 3T3-L1 Cells via a Mechanism that Implicates PPARγ together with TNF-α and IL-6

Abstract
:1. Introduction
2. Results and Discussion
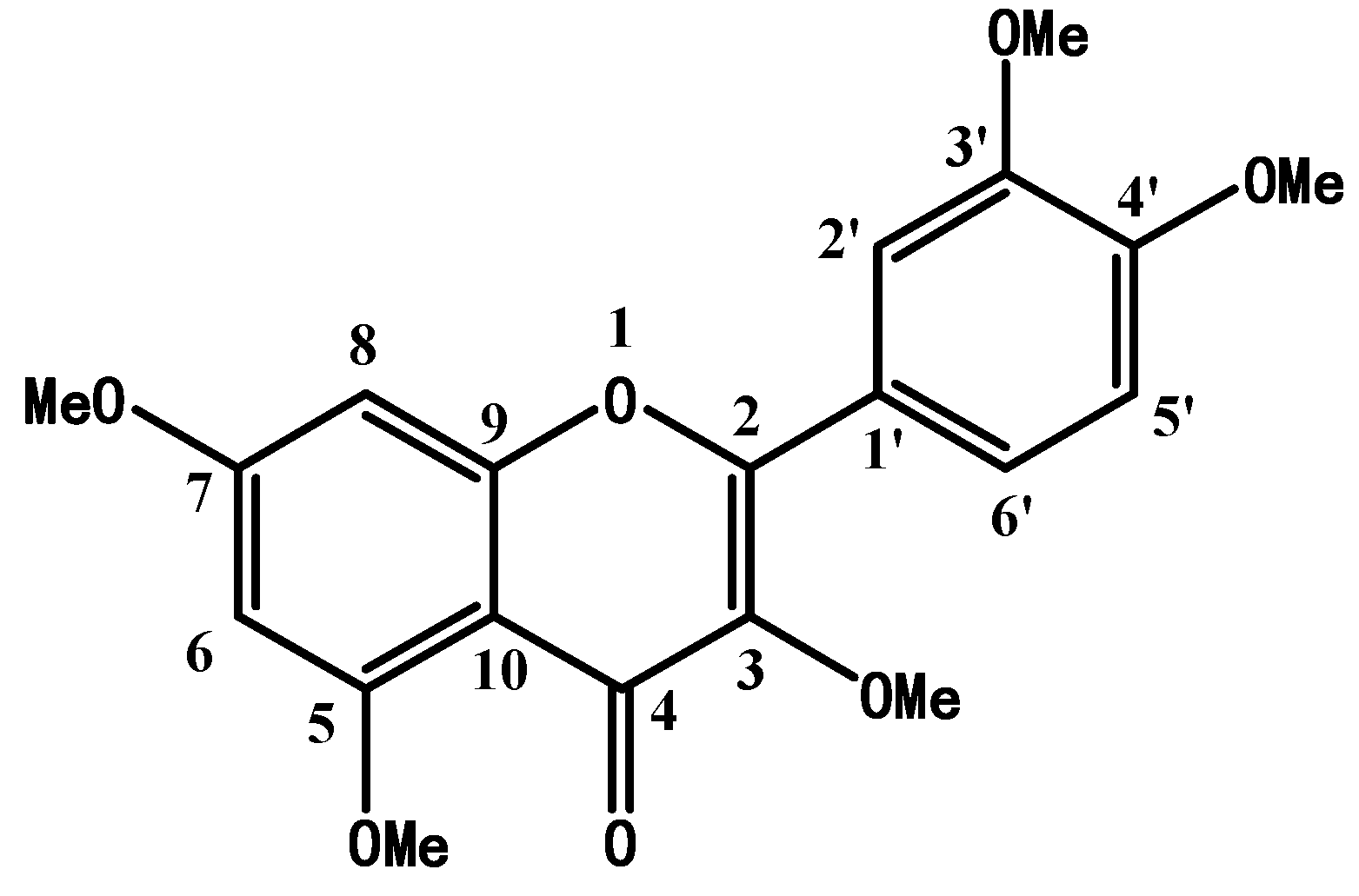
2.1. Structure Confirmation and Identification of Synthetic PMQ


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 13C-NMR | 1H-NMR |
|---|---|---|
| δC | δH | |
| 1 | - | - |
| 2 | 152.5 sp2 | - |
| 3 | 141.1 sp2 | - |
| 4 | 173.9 sp2 | - |
| 5 | 158.7 sp2 | - |
| 6 | 95.7 sp2 | 6.343–6.348 d |
| 7 | 163.8 sp2 | - |
| 8 | 92.3 sp2 | 6.502–6.508 d |
| 9 | 160.9 sp2 | - |
| 10 | 109.4 sp2 | - |
| 1’ | 123.3 sp2 | - |
| 2’ | 111.1 sp2 | 7.715 s |
| 3’ | 148.6 sp2 | - |
| 4’ | 150.7 sp2 | - |
| 5’ | 110.7 sp2 | 6.968–6.911 d |
| 6’ | 121.5 sp2 | 7.697–7.715 d |
| 5X-OCH3 | 55.7 sp3 | 3.847–3.989 m |
| 55.9 sp3 | ||
| 56.0 sp3 | ||
| 56.3 sp3 | ||
| 59.9 sp3 |
2.2. Adipogenesis in Mouse 3T3-L1 Preadipocytes and the Effects of PMQ on Cytotoxicity and Lipid Content in Maturate Adipocyte

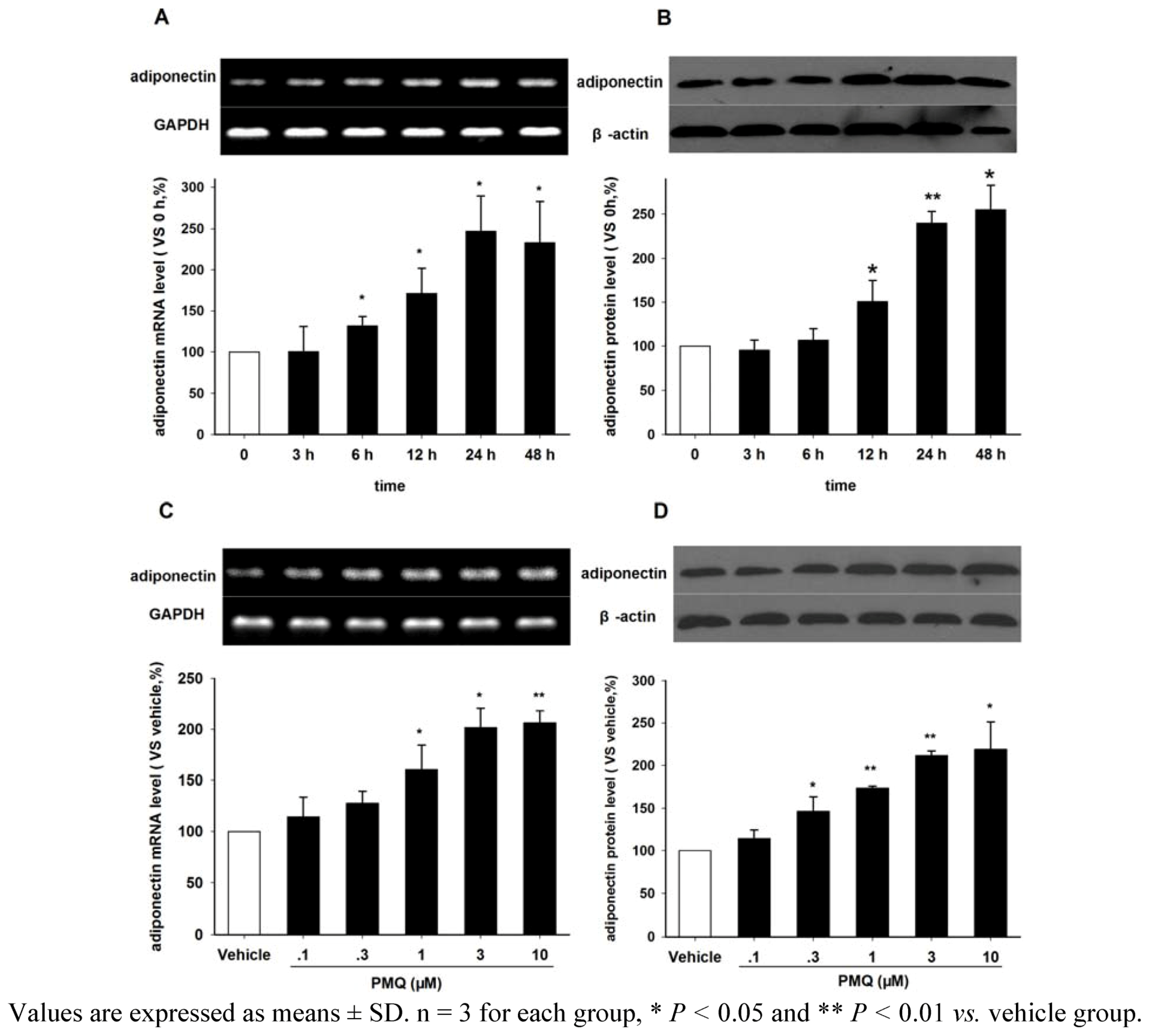
2.3. Effect of PMQ on Adiponectin Expression in 3T3-L1 Adipocytes

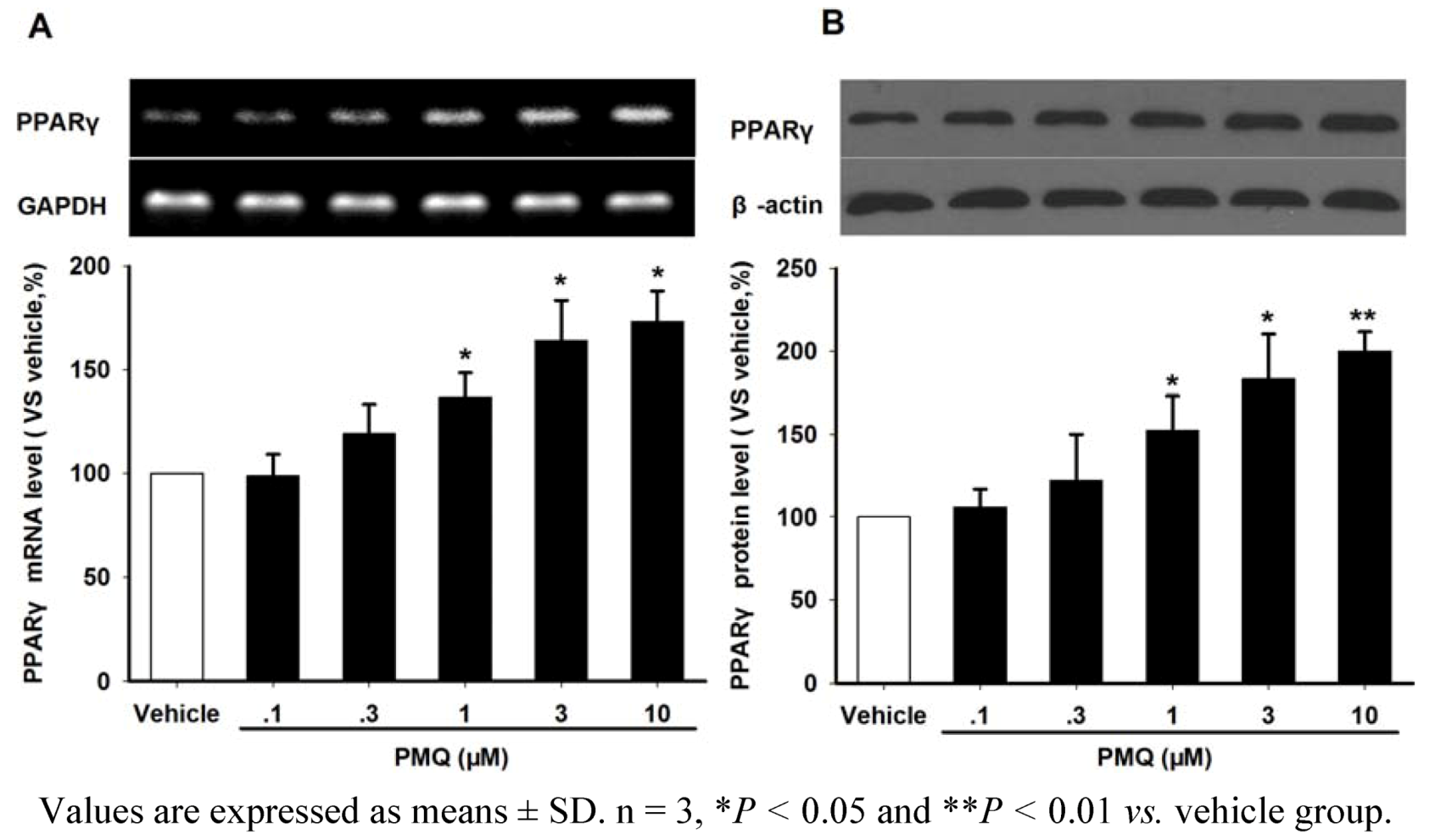
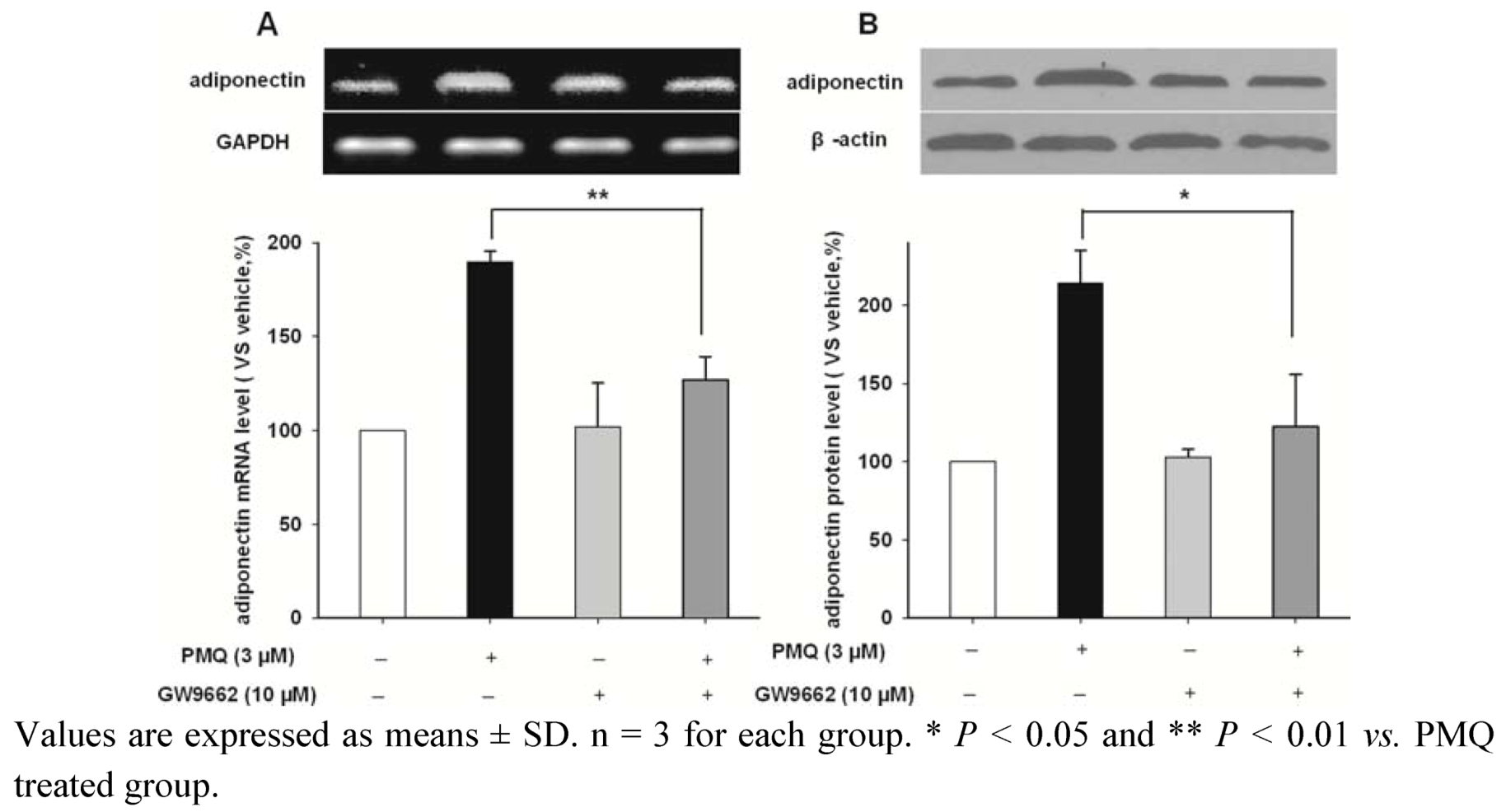
2.4. PMQ Regulates Adiponectin Expression via PPARγ Pathway


2.5. Effects of PMQ on Expression and Secretion of TNF-α and IL-6

3. Experimental
3.1. Chemicals
3.2. Preparation of Pentamethylquercetin
3.3. Cell Culture and Treatment
3.4. Oil Red O Staining
3.5. Cell Viability Assay
3.6. Reverse Transcription-Polymerase Chain Reaction
| Gene | Sequences (5’ -3’) | Accession No | Length (bp) |
|---|---|---|---|
| PPARγ | Forward: ACTGCCTATGAGCACTTCAC | NM_011146 | 448 |
| Reverse: CAATCGGATGGTTCTTCGGA | |||
| Adiponectin | Forward: GGGTGAGACAGGAGATGTTGGAATG | NM_009605 | 478 |
| Reverse: GCCAGTAAATGTAGAGTCGTTGACG | |||
| IL-6 | Forward: AGTTGCCTTCTTGGGACTGA | NM_031168 | 521 |
| Reverse: GCCACTCCTTCTGTGACTCC | |||
| TNF-α | Forward: TGGAGTCATTGCTCTGTGAAGGGA | NM_013693 | 250 |
| Reverse: AGTCCTTGATGGTGGTGCATGAGA | |||
| GAPDH | Forward: GACAAAATGGTGAAGGTCGGTG | NM_008084 | 256 |
| Reverse: TGATGTTAGTGGGGTCTCGCTC |
3.7. Western Blotting
3.8. ELISIA
3.9. Statistical Analysis
4. Conclusions
Acknowledgements
References and Notes
- Haslam, D.W.; James, W.P. Obesity. Lancet 2005, 366, 1197–1209. [Google Scholar] [CrossRef]
- Szmitko, P.E.; Teoh, H.; Stewart, D.J.; Verma, S. Adiponectin and cardiovascular disease: State of the art? Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1655–H1663. [Google Scholar]
- Scherer, P.E.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. A novel serum protein similar to C1q, produced exclusively in adipocytes. J. Biol. Chem. 1995, 270, 26746–26749. [Google Scholar]
- Hu, E.; Liang, P.; Spiegelman, B.M. AdipoQ is a novel adipose-specific gene dysregulated in obesity. J. Biol. Chem. 1996, 271, 10697–10703. [Google Scholar] [CrossRef]
- Lara-Castro, C.; Fu, Y.; Chung, B.H.; Garvey, W.T. Adiponectin and the metabolic syndrome: Mechanisms mediating risk for metabolic and cardiovascular disease. Curr. Opin. Lipidol. 2007, 18, 263–270. [Google Scholar] [CrossRef]
- Oh, D.K.; Ciaraldi, T.; Henry, R.R. Adiponectin in health and disease. Diabetes Obes. Metab. 2007, 9, 282–289. [Google Scholar] [CrossRef]
- Matsuda, M.; Shimomura, I.; Sata, M.; Arita, Y.; Nishida, M.; Maeda, N.; Kumada, M.; Okamoto, Y.; Nagaretani, H.; Nishizawa, H.; et al. Role of adiponectin in preventing vascular stenosis. The missing link of adipo-vascular axis. J. Biol. Chem. 2002, 277, 37487–37491. [Google Scholar]
- Ouchi, N.; Kihara, S.; Arita, Y.; Nishida, M.; Matsuyama, A.; Okamoto, Y.; Ishigami, M.; Kuriyama, H.; Kishida, K.; Nishizawa, H.; et al. Adipocyte-derived plasma protein, adiponectin, suppresses lipid accumulation and class A scavenger receptor expression in human monocyte-derived macrophages. Circulation 2001, 103, 1057–1063. [Google Scholar] [CrossRef]
- Ouchi, N.; Kihara, S.; Arita, Y.; Okamoto, Y.; Maeda, K.; Kuriyama, H.; Hotta, K.; Nishida, M.; Takahashi, M.; Muraguchi, M.; et al. Adiponectin, an adipocyte-derived plasma protein, inhibits endothelial NF-kappaB signaling through a cAMP-dependent pathway. Circulation 2000, 102, 1296–1301. [Google Scholar] [CrossRef]
- Ouchi, N.; Kihara, S.; Funahashi, T.; Nakamura, T.; Nishida, M.; Kumada, M.; Okamoto, Y.; Ohashi, K.; Nagaretani, H.; Kishida, K.; et al. Reciprocal association of C-reactive protein with adiponectin in blood stream and adipose tissue. Circulation 2003, 107, 671–674. [Google Scholar] [CrossRef]
- Arita, Y.; Kihara, S.; Ouchi, N.; Takahashi, M.; Maeda, K.; Miyagawa, J.; Hotta, K.; Shimomura, I.; Nakamura, T.; Miyaoka, K.; et al. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem. Biophys. Res. Commun. 1999, 257, 79–83. [Google Scholar] [CrossRef]
- Matsubara, M.; Maruoka, S.; Katayose, S. Decreased plasma adiponectin concentrations in women with dyslipidemia. J. Clin. Endocrinol. Metab. 2002, 87, 2764–2769. [Google Scholar] [CrossRef]
- Hotta, K.; Funahashi, T.; Arita, Y.; Takahashi, M.; Matsuda, M.; Okamoto, Y.; Iwahashi, H.; Kuriyama, H.; Ouchi, N.; Maeda, K.; et al. Plasma concentrations of a novel, adipose-specific protein, adiponectin, in type 2 diabetic patients. Arterioscler Thromb. Vasc. Biol. 2000, 20, 1595–1599. [Google Scholar] [CrossRef]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2006, 69, 273–278. [Google Scholar]
- Knekt, P.; Kumpulainen, J.; Jarvinen, R.; Rissanen, H.; Heliovaara, M.; Reunanen, A.; Hakulinen, T.; Aromaa, A. Flavonoid intake and risk of chronic diseases. Am. J. Clin. Nutr. 2002, 76, 560–568. [Google Scholar]
- Hertog, M.G.; Feskens, E.J.; Hollman, P.C.; Katan, M.B.; Kromhout, D. Dietary antioxidant flavonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet 1993, 342, 1007–1011. [Google Scholar]
- Mink, P.J.; Scrafford, C.G.; Barraj, L.M.; Harnack, L.; Hong, C.P.; Nettleton, J.A.; Jacobs, D.R., Jr. Flavonoid intake and cardiovascular disease mortality: A prospective study in postmenopausal women. Am. J. Clin. Nutr. 2007, 85, 895–909. [Google Scholar]
- Walle, U.K.; Walle, T. Bioavailable flavonoids: Cytochrome P450-mediated metabolism of methoxyflavones. Drug Metab. Dispos. 2007, 35, 1985–1989. [Google Scholar] [CrossRef]
- Li, R.W.; Theriault, A.G.; Au, K.; Douglas, T.D.; Casaschi, A.; Kurowska, E.M.; Mukherjee, R. Citrus polymethoxylated flavones improve lipid and glucose homeostasis and modulate adipocytokines in fructose-induced insulin resistant hamsters. Life Sci. 2006, 79, 365–373. [Google Scholar] [CrossRef]
- Liu, L.; Shan, S.; Zhang, K.; Ning, Z.Q.; Lu, X.P.; Cheng, Y.Y. Naringenin and hesperetin, two flavonoids derived from Citrus aurantium up-regulate transcription of adiponectin. Phytother. Res. 2008, 22, 1400–1403. [Google Scholar] [CrossRef]
- Kunimasa, K.; Kuranuki, S.; Matsuura, N.; Iwasaki, N.; Ikeda, M.; Ito, A.; Sashida, Y.; Mimaki, Y.; Yano, M.; Sato, M.; et al. Identification of nobiletin, a polymethoxyflavonoid, as an enhancer of adiponectin secretion. Bioorg. Med. Chem. Lett. 2009, 19, 2062–2064. [Google Scholar]
- Hibasami, H.; Mitani, A.; Katsuzaki, H.; Imai, K.; Yoshioka, K.; Komiya, T. Isolation of five types of flavonol from seabuckthorn (Hippophae rhamnoides) and induction of apoptosis by some of the flavonols in human promyelotic leukemia HL-60 cells. Int. J. Mol. Med. 2005, 15, 805–809. [Google Scholar]
- Mao, Z.; Liang, Y.; Du, X.; Sun, Z. 3,3',4',5,7-Pentamethylquercetin reduces angiotensin II-induced cardiac hypertrophy and apoptosis in rats. Can. J. Physiol. Pharmacol. 2009, 87, 720–728. [Google Scholar] [CrossRef]
- Dong, H.; Gou, Y.-L.; Cao, S.-G.; Chen, S.-X.; Sim, K.-Y.; Goh, S.-H.; Kini, R.M. Eicosenones and methylated flavonols from Amomumkoenigii. Phytochemistry 1999, 50, 899–902. [Google Scholar]
- Heijnen, C.G.M.; Haenen, G.R.M.M.; Vekemansb, J.A.J.M.; Basta, A. Peroxynitrite scavenging of flavonoids: Structure activity relationship. Environ. Toxicol. Pharmacol. 2001, 10, 199–206. [Google Scholar] [CrossRef]
- Fasshauer, M.; Kralisch, S.; Klier, M.; Lossner, U.; Bluher, M.; Klein, J.; Paschke, R. Adiponectin gene expression and secretion is inhibited by interleukin-6 in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2003, 301, 1045–1050. [Google Scholar] [CrossRef]
- Kim, K.Y.; Kim, J.K.; Jeon, J.H.; Yoon, S.R.; Choi, I.; Yang, Y. c-Jun N-terminal kinase is involved in the suppression of adiponectin expression by TNF-alpha in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2005, 327, 460–467. [Google Scholar] [CrossRef]
- Must, A.; Spadano, J.; Coakley, E.H.; Field, A.E.; Colditz, G.; Dietz, W.H. The disease burden associated with overweight and obesity. JAMA 1999, 282, 1523–1529. [Google Scholar]
- Yang, W.S.; Lee, W.J.; Funahashi, T.; Tanaka, S.; Matsuzawa, Y.; Chao, C.L.; Chen, C.L.; Tai, T.Y.; Chuang, L.M. Weight reduction increases plasma levels of an adipose-derived anti-inflammatory protein, adiponectin. J. Clin. Endocrinol. Metab. 2001, 86, 3815–3819. [Google Scholar]
- Kondo, H.; Shimomura, I.; Matsukawa, Y.; Kumada, M.; Takahashi, M.; Matsuda, M.; Ouchi, N.; Kihara, S.; Kawamoto, T.; Sumitsuji, S.; Funahashi, T.; Matsuzawa, Y. Association of adiponectin mutation with type 2 diabetes: A candidate gene for the insulin resistance syndrome. Diabetes 2002, 51, 2325–2328. [Google Scholar]
- Michalik, L.; Auwerx, J.; Berger, J.P.; Chatterjee, V.K.; Glass, C.K.; Gonzalez, F.J.; Grimaldi, P.A.; Kadowaki, T.; Lazar, M.A.; O'Rahilly, S.; et al. International Union of Pharmacology. LXI. Peroxisome proliferator-activated receptors. Pharmacol. Rev. 2006, 58, 726–741. [Google Scholar] [CrossRef]
- He, W.; Barak, Y.; Hevener, A.; Olson, P.; Liao, D.; Le, J.; Nelson, M.; Ong, E.; Olefsky, J.M.; Evans, R.M. Adipose-specific peroxisomeproliferator-activated receptor gamma knockout causes insulin resistance in fat and liver but not in muscle. Proc. Natl. Acad. Sci. USA 2003, 100, 15712–15717. [Google Scholar]
- Yu, J.G.; Javorschi, S.; Hevener, A.L.; Kruszynska, Y.T.; Norman, R.A.; Sinha, M.; Olefsky, J.M. The effect of thiazolidinediones on plasma adiponectin levels in normal, obese, and type 2 diabetic subjects. Diabetes 2002, 51, 2968–2974. [Google Scholar] [CrossRef]
- Kanatani, Y.; Usui, I.; Ishizuka, K.; Bukhari, A.; Fujisaka, S.; Urakaze, M.; Haruta, T.; Kishimoto, T.; Naka, T.; Kobayashi, M. Effects of pioglitazone on suppressor of cytokine signaling 3 expression: Potential mechanisms for its effects on insulin sensitivity and adiponectin expression. Diabetes 2007, 56, 795–803. [Google Scholar]
- Iwaki, M.; Matsuda, M.; Maeda, N.; Funahashi, T.; Matsuzawa, Y.; Makishima, M.; Shimomura, I. Induction of adiponectin, a fat-derived antidiabetic and antiatherogenic factor, by nuclear receptors. Diabetes 2003, 52, 1655–1663. [Google Scholar]
- Seo, J.B.; Moon, H.M.; Noh, M.J.; Lee, Y.S.; Jeong, H.W.; Yoo, E.J.; Kim, W.S.; Park, J.; Youn, B.S.; Kim, J.W.; et al. Adipocyte determination- and differentiation-dependent factor 1/sterol regulatory element-binding protein 1c regulates mouse adiponectin expression. J. Biol. Chem. 2004, 279, 22108–22117. [Google Scholar]
- Fernandez-Real, J.M.; Vayreda, M.; Richart, C.; Gutierrez, C.; Broch, M.; Vendrell, J.; Ricart, W. Circulating interleukin 6 levels, blood pressure, and insulin sensitivity in apparently healthy men and women. J. Clin. Endocrinol. Metab. 2001, 86, 1154–1159. [Google Scholar] [CrossRef]
- Mohamed-Ali, V.; Goodrick, S.; Rawesh, A.; Katz, D.R.; Miles, J.M.; Yudkin, J.S.; Klein, S.; Coppack, S.W. Subcutaneous adipose tissue releases interleukin-6, but not tumor necrosis factor-alpha, in vivo. J. Clin. Endocrinol. Metab. 1997, 82, 4196–4200. [Google Scholar] [CrossRef]
- Straub, R.H.; Hense, H.W.; Andus, T.; Scholmerich, J.; Riegger, G.A.; Schunkert, H. Hormone replacement therapy and interrelation between serum interleukin-6 and body mass index in postmenopausal women: A population-based study. J. Clin. Endocrinol. Metab. 2000, 85, 1340–1344. [Google Scholar]
- Muller, S.; Martin, S.; Koenig, W.; Hanifi-Moghaddam, P.; Rathmann, W.; Haastert, B.; Giani, G.; Illig, T.; Thorand, B.; Kolb, H. Impaired glucose tolerance is associated with increased serum concentrations of interleukin 6 and co-regulated acute-phase proteins but not TNF-alpha or its receptors. Diabetologia 2002, 45, 805–812. [Google Scholar] [CrossRef]
- Kado, S.; Nagase, T.; Nagata, N. Circulating levels of interleukin-6, its soluble receptor and interleukin-6/interleukin-6 receptor complexes in patients with type 2 diabetes mellitus. Acta Diabetol. 1999, 36, 67–72. [Google Scholar] [CrossRef]
- Pickup, J.C.; Mattock, M.B.; Chusney, G.D.; Burt, D. NIDDM as a disease of the innate immune system: Association of acute-phase reactants and interleukin-6 with metabolic syndrome X. Diabetologia 1997, 40, 1286–1292. [Google Scholar] [CrossRef]
- Zhang, B.; Berger, J.; Hu, E.; Szalkowski, D.; White-Carrington, S.; Spiegelman, B.M.; Moller, D.E. Negative regulation of peroxisomeproliferator-activated receptor-gamma gene expression contributes to the antiadipogenic effects of tumor necrosis factor-alpha. Mol. Endocrinol. 1996, 10, 1457–1466. [Google Scholar] [CrossRef]
- Kita, A.; Yamasaki, H.; Kuwahara, H.; Moriuchi, A.; Fukushima, K.; Kobayashi, M.; Fukushima, T.; Takahashi, R.; Abiru, N.; Uotani, S.; Kawasaki, E.; Eguchi, K. Identification of the promoter region required for human adiponectin gene transcription: Association with CCAAT/enhancer binding protein-beta and tumor necrosis factor-alpha. Biochem. Biophys. Res. Commun. 2005, 331, 484–490. [Google Scholar] [CrossRef]
- Ron, D.; Brasier, A.R.; McGehee, R.E., Jr.; Habener, J.F. Tumor necrosis factor-induced reversal of adipocytic phenotype of 3T3-L1 cells is preceded by a loss of nuclear CCAAT/enhancer binding protein (C/EBP). J. Clin. Invest. 1992, 89, 223–233. [Google Scholar] [CrossRef]
- Lim, J.Y.; Kim, W.H.; Park, S.I. GO6976 prevents TNF-alpha-induced suppression of adiponectin expression in 3T3-L1 adipocytes: Putative involvement of protein kinase C. FEBS Lett. 2008, 582, 3473–3478. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compound PMQ is available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, L.; He, T.; Han, Y.; Sheng, J.-Z.; Jin, S.; Jin, M.-W. Pentamethylquercetin Improves Adiponectin Expression in Differentiated 3T3-L1 Cells via a Mechanism that Implicates PPARγ together with TNF-α and IL-6. Molecules 2011, 16, 5754-5768. https://doi.org/10.3390/molecules16075754
Chen L, He T, Han Y, Sheng J-Z, Jin S, Jin M-W. Pentamethylquercetin Improves Adiponectin Expression in Differentiated 3T3-L1 Cells via a Mechanism that Implicates PPARγ together with TNF-α and IL-6. Molecules. 2011; 16(7):5754-5768. https://doi.org/10.3390/molecules16075754
Chicago/Turabian StyleChen, Lei, Ting He, Yi Han, Ji-Zhong Sheng, Si Jin, and Man-Wen Jin. 2011. "Pentamethylquercetin Improves Adiponectin Expression in Differentiated 3T3-L1 Cells via a Mechanism that Implicates PPARγ together with TNF-α and IL-6" Molecules 16, no. 7: 5754-5768. https://doi.org/10.3390/molecules16075754