Parthenolide Induces Apoptosis and Cell Cycle Arrest of Human 5637 Bladder Cancer Cells In Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussions
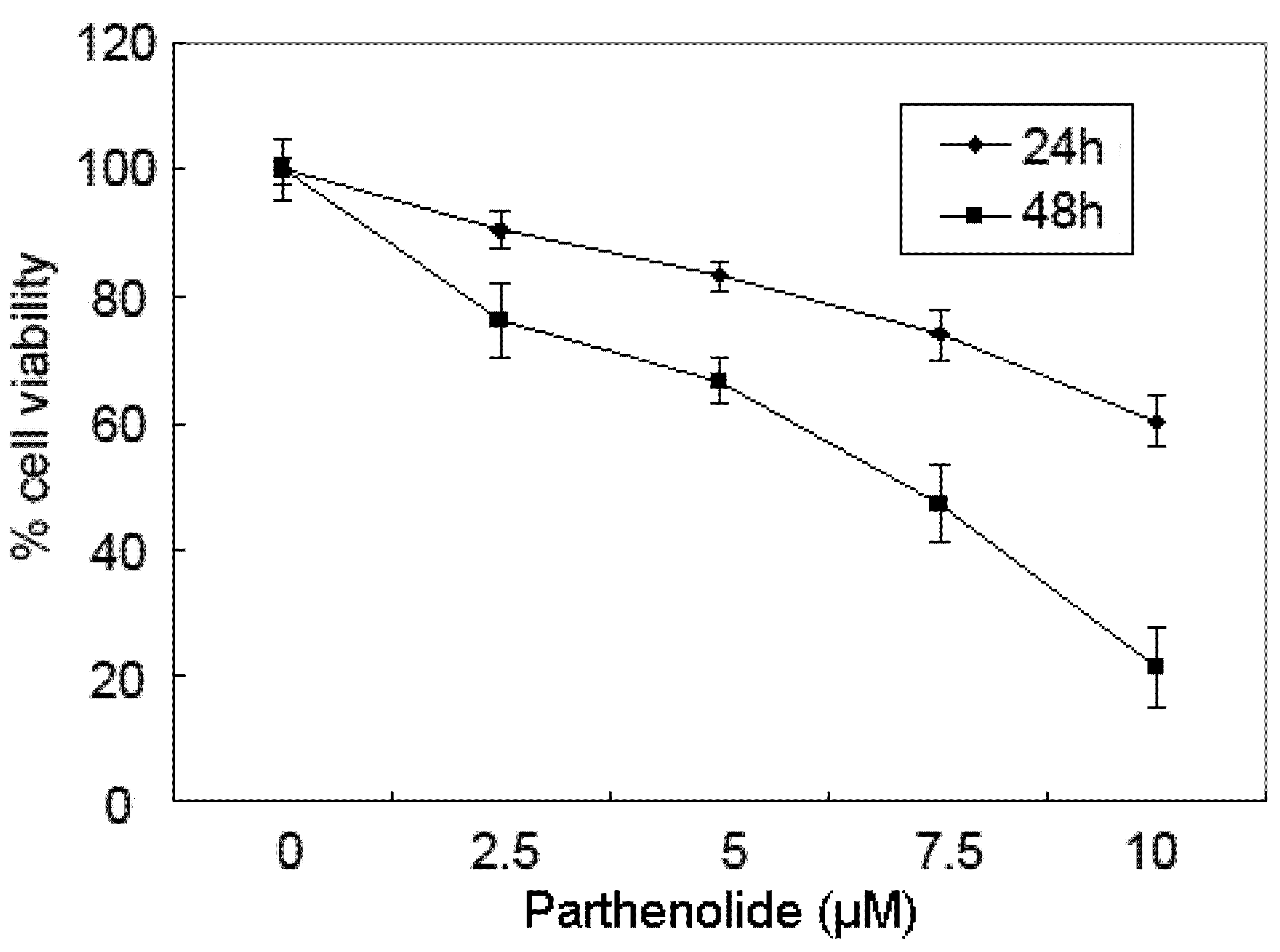
2.1. Cytotoxicity Effects of PTL on 5637 Bladder Cancer Cells

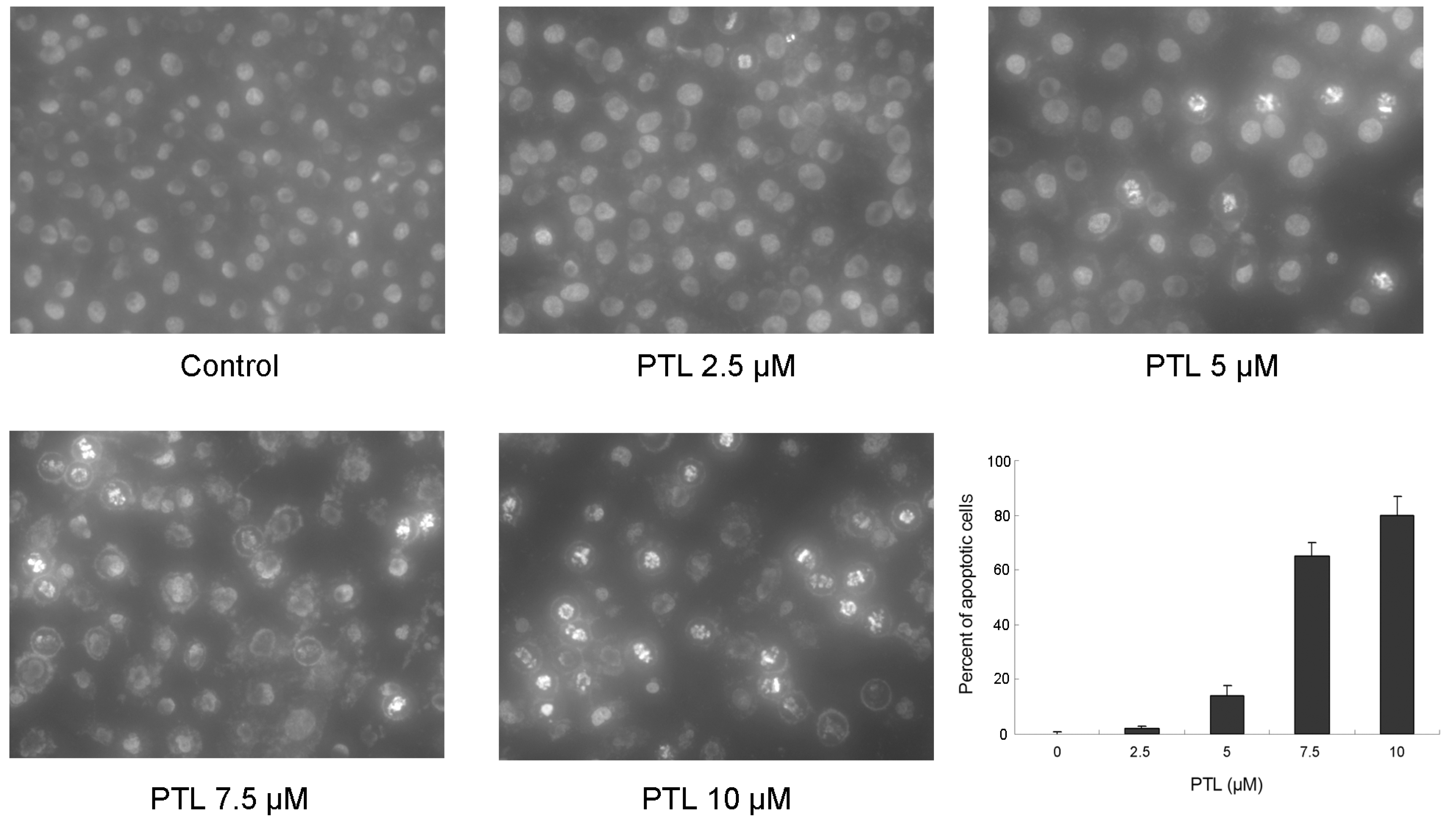
2.2. Induction of Apoptosis of 5637 Cells by PTL

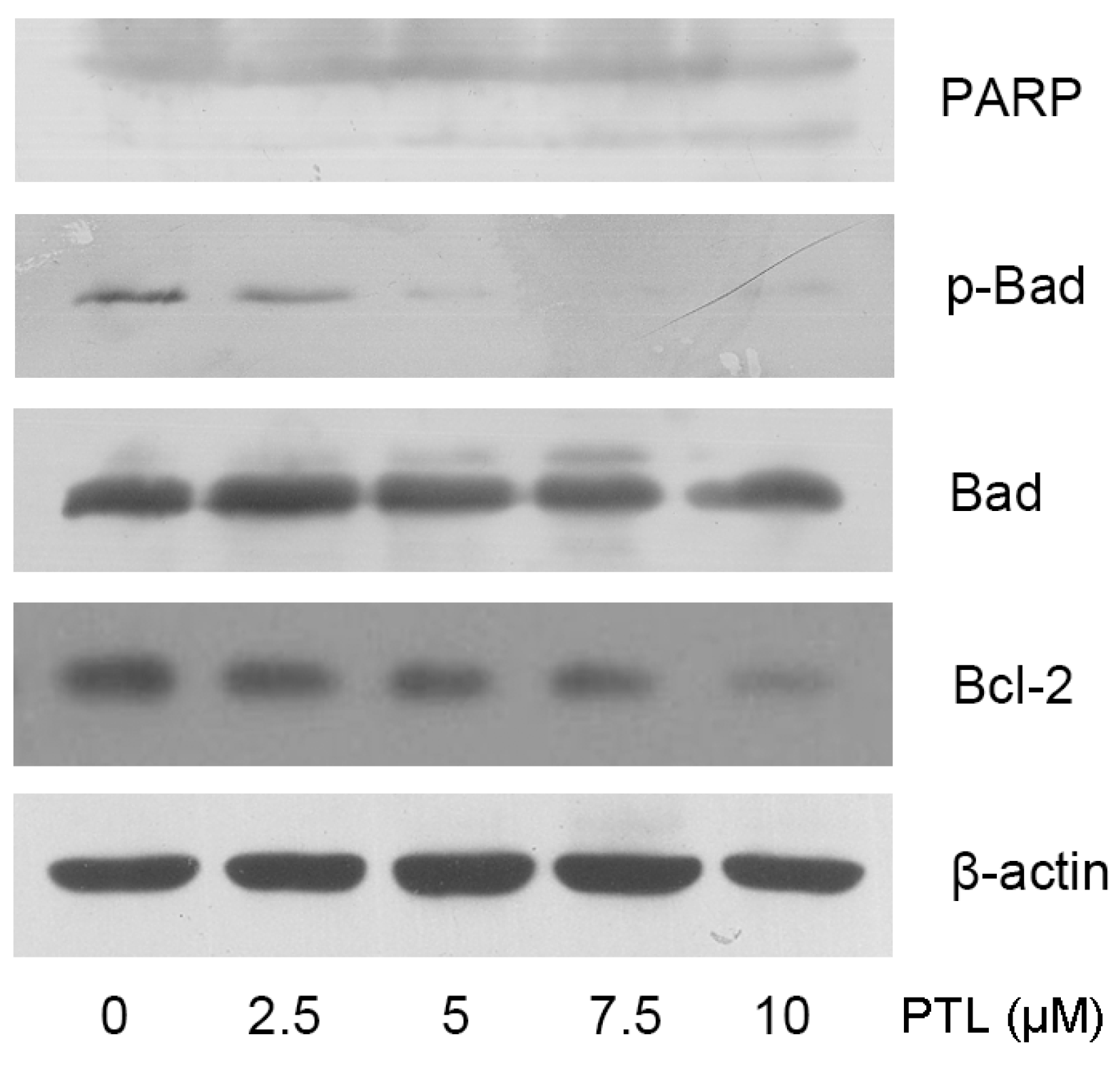
2.3. PTL-Induced Apoptosis Mediated by Bcl-2 Family Modulation and PARP Cleavage

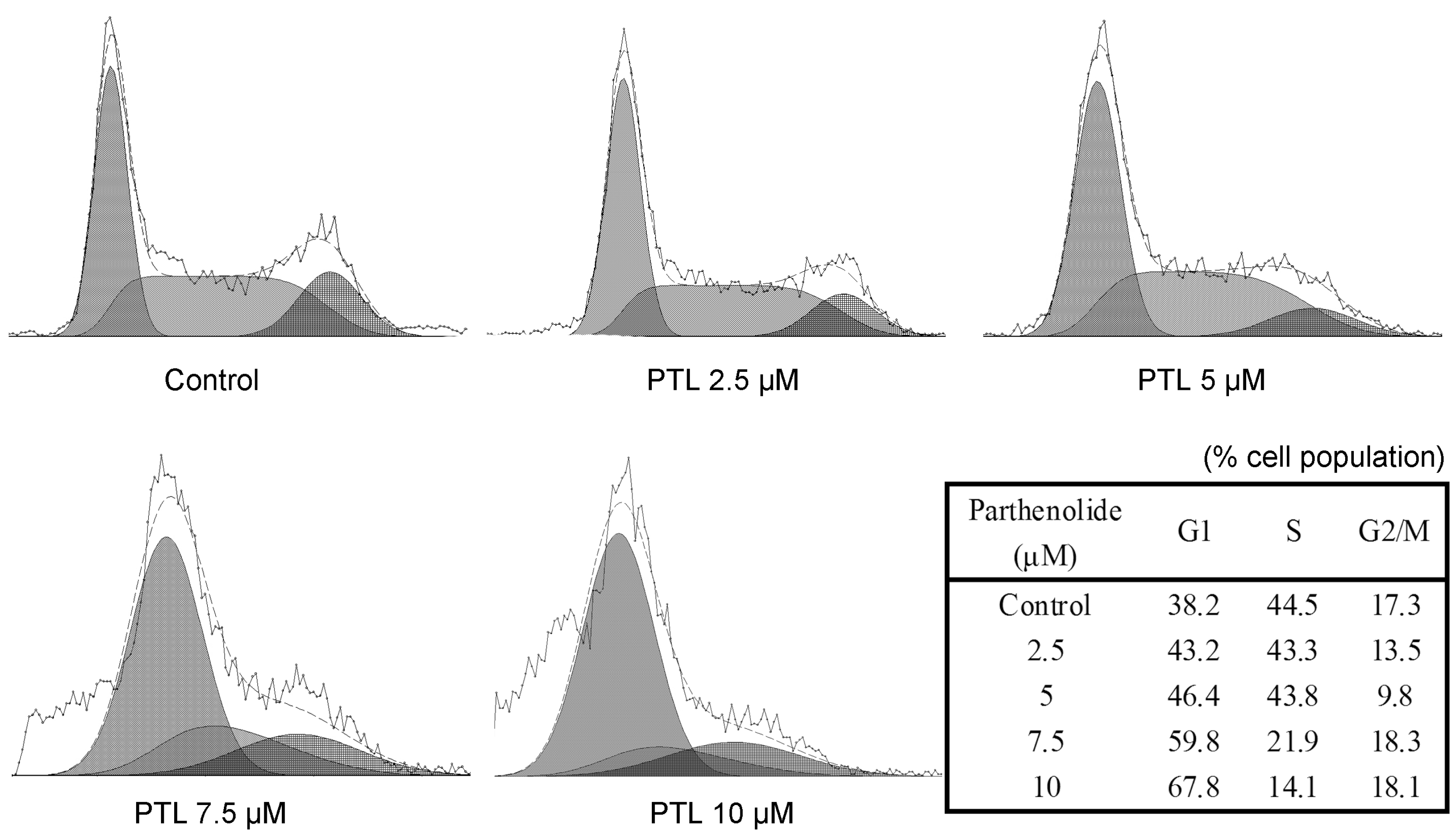
2.4. Cell Cycle Accumulation at G1 Phase after Treatment with PTL

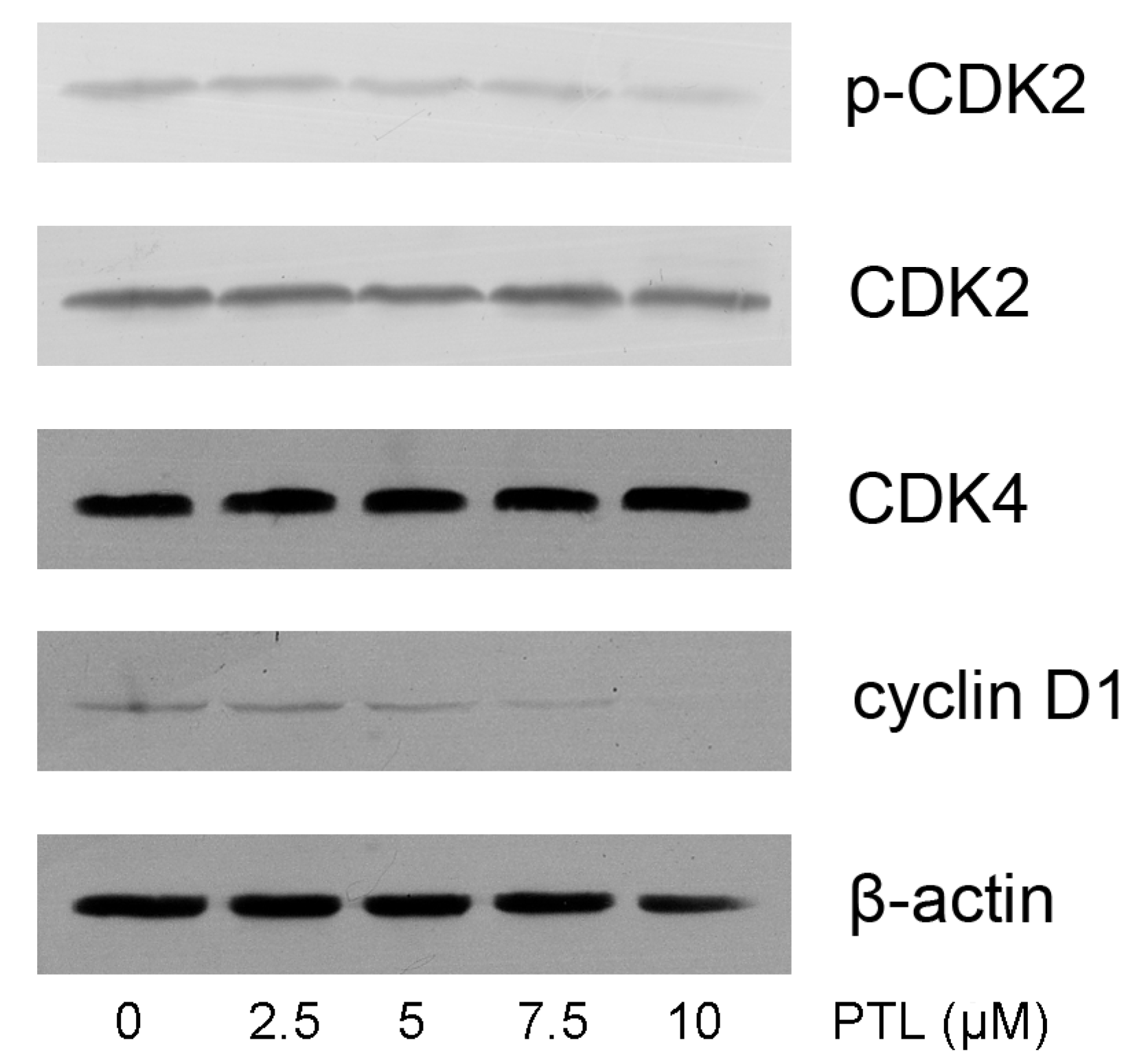
2.5. Modulation of Cell Cycle related Proteins by PTL

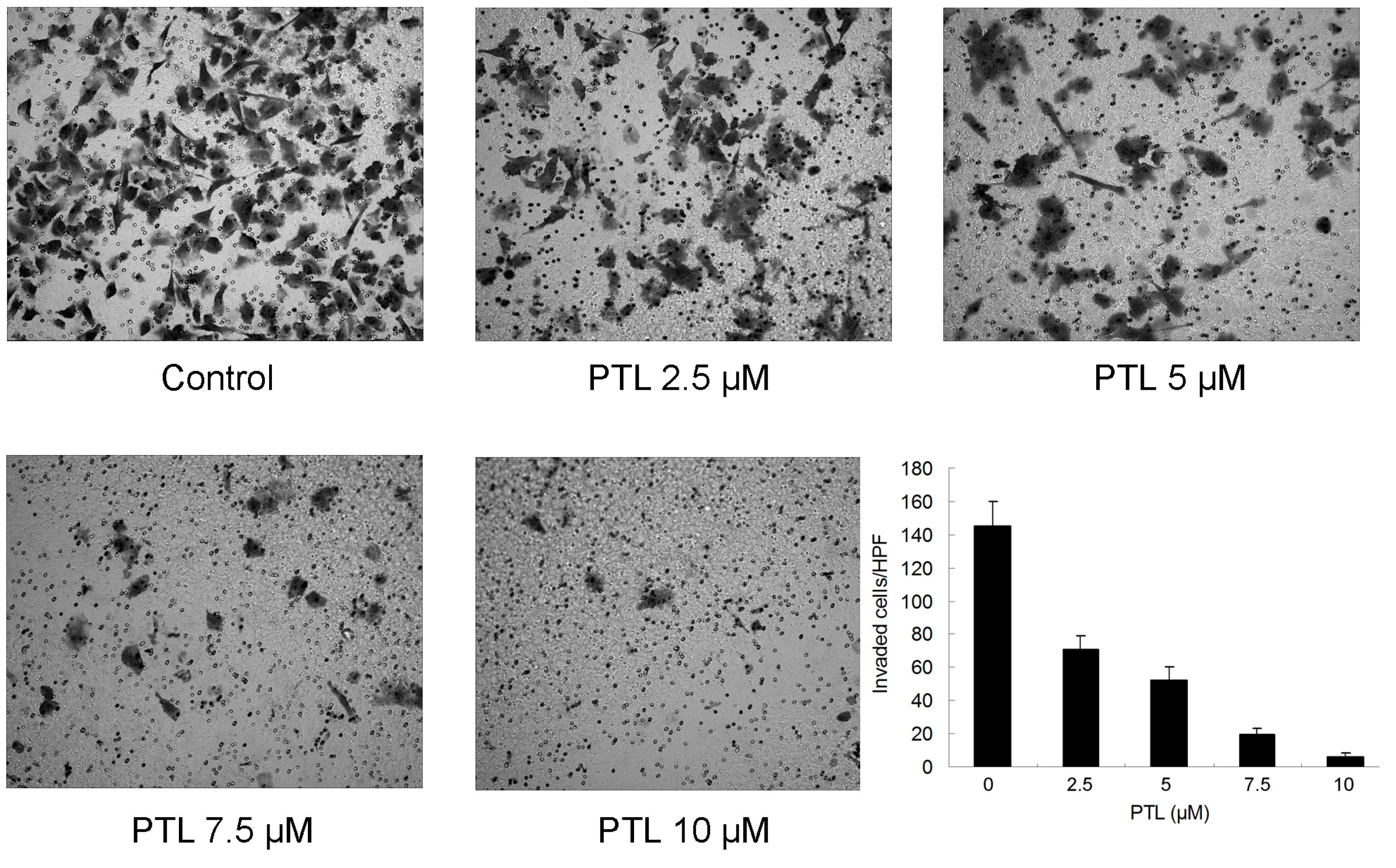
2.6. Inhibition of Cell Invasion by PTL

3. Experimental
3.1. Cell Culture and Reagents
3.2. Cell Viability Assay
3.3. Nuclear Morphology
3.4. Cell Cycle Distribution Analysis
3.5. In Vitro Cell Invasion Assay
3.6. Western Blot Analysis
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
References and Notes
- Jemal, A.; Siegel, R.; Xu, J.; Ward, E. Cancer statistics, 2010. CA Cancer J. Clin. 2010, 60, 277–300. [Google Scholar] [CrossRef]
- Rosenberg, J.E.; Carroll, P.R.; Small, E.J. Update on chemotherapy for advanced bladder cancer. J. Urol. 2005, 174, 14–20. [Google Scholar] [CrossRef]
- Biggs, M.J.; Johnson, E.S.; Persaud, N.P.; Ratcliffe, D.M. Platelet aggregation in patients using feverfew for migraine. Lancet 1982, 2, 776. [Google Scholar]
- Hall, I.H.; Lee, K.H.; Starnes, C.O.; Sumida, Y.; Wu, R.Y.; Waddell, T.G.; Cochran, J.W.; Gerhart, K.G. Anti-inflammatory activity of sesquiterpene lactones and related compounds. J. Pharm. Sci. 1979, 68, 537–542. [Google Scholar]
- Jain, N.K.; Kulkarni, S.K. Antinociceptive and anti-inflammatory effects of tanacetum parthenium l. Extract in mice and rats. J. Ethnopharmacol. 1999, 68, 251–259. [Google Scholar] [CrossRef]
- Kim, J.H.; Liu, L.; Lee, S.O.; Kim, Y.T.; You, K.R.; Kim, D.G. Susceptibility of cholangiocarcinoma cells to parthenolide-induced apoptosis. Cancer Res. 2005, 65, 6312–6320. [Google Scholar] [CrossRef]
- Park, J.H.; Liu, L.; Kim, I.H.; Kim, J.H.; You, K.R.; Kim, D.G. Identification of the genes involved in enhanced fenretinide-induced apoptosis by parthenolide in human hepatoma cells. Cancer Res. 2005, 65, 2804–2814. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, H.; Gu, P.; Bai, J.; Margolick, J.B.; Zhang, Y. Nf-kappab pathway inhibitors preferentially inhibit breast cancer stem-like cells. Breast Cancer Res. Treat. 2008, 111, 419–427. [Google Scholar] [CrossRef]
- Guzman, M.L.; Rossi, R.M.; Karnischky, L.; Li, X.; Peterson, D.R.; Howard, D.S.; Jordan, C.T. The sesquiterpene lactone parthenolide induces apoptosis of human acute myelogenous leukemia stem and progenitor cells. Blood 2005, 105, 4163–4169. [Google Scholar]
- Wen, J.; You, K.R.; Lee, S.Y.; Song, C.H.; Kim, D.G. Oxidative stress-mediated apoptosis. The anticancer effect of the sesquiterpene lactone parthenolide. J. Biol. Chem. 2002, 277, 38954–38964. [Google Scholar]
- Sobota, R.; Szwed, M.; Kasza, A.; Bugno, M.; Kordula, T. Parthenolide inhibits activation of signal transducers and activators of transcription (stats) induced by cytokines of the il-6 family. Biochem. Biophys. Res. Commun. 2000, 267, 329–333. [Google Scholar] [CrossRef]
- Hehner, S.P.; Heinrich, M.; Bork, P.M.; Vogt, M.; Ratter, F.; Lehmann, V.; Schulze-Osthoff, K.; Droge, W.; Schmitz, M.L. Sesquiterpene lactones specifically inhibit activation of nf-kappa b by preventing the degradation of i kappa b-alpha and i kappa b-beta. J. Biol. Chem. 1998, 273, 1288–1297. [Google Scholar]
- Bernardi, R.; Negri, C.; Donzelli, M.; Guano, F.; Torti, M.; Prosperi, E.; Scovassi, A.I. Activation of poly(adp-ribose)polymerase in apoptotic human cells. Biochimie 1995, 77, 378–384. [Google Scholar] [CrossRef]
- Yafi, F.A.; Kassouf, W. Management of patients with advanced bladder cancer following major response to systemic chemotherapy. Expert Rev. Anticancer Ther. 2009, 9, 1757–1764. [Google Scholar] [CrossRef]
- Shanmugam, R.; Kusumanchi, P.; Cheng, L.; Crooks, P.; Neelakantan, S.; Matthews, W.; Nakshatri, H.; Sweeney, C.J. A water-soluble parthenolide analogue suppresses in vivo prostate cancer growth by targeting nfkappab and generating reactive oxygen species. Prostate 2010, 70, 1074–1086. [Google Scholar] [CrossRef]
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nature 2001, 411, 342–348. [Google Scholar] [CrossRef]
- Hayashi, S.; Sakurai, H.; Hayashi, A.; Tanaka, Y.; Hatashita, M.; Shioura, H. Inhibition of nf-kappab by combination therapy with parthenolide and hyperthermia and kinetics of apoptosis induction and cell cycle arrest in human lung adenocarcinoma cells. Int. J. Mol. Med. 2010, 25, 81–87. [Google Scholar]
- Zanotto-Filho, A.; Braganhol, E.; Schroder, R.; de Souza, L.H.; Dalmolin, R.J.; Pasquali, M.A.; Gelain, D.P.; Battastini, A.M.; Moreira, J.C. Nfkappab inhibitors induce cell death in glioblastomas. Biochem. Pharmacol. 2011, 81, 412–424. [Google Scholar]
- Shanmugam, R.; Kusumanchi, P.; Appaiah, H.; Cheng, L.; Crooks, P.; Neelakantan, S.; Peat, T.; Klaunig, J.; Matthews, W.; Nakshatri, H.; et al. A water soluble parthenolide analog suppresses in vivo tumor growth of two tobacco-associated cancers, lung and bladder cancer, by targeting nf-kappab and generating reactive oxygen species. Int. J. Cancer 2011, 128, 2481–2494. [Google Scholar]
- Mowat, M.R. P53 in tumor progression: Life, death, and everything. Adv. Cancer Res. 1998, 74, 25–48. [Google Scholar] [CrossRef]
- Cooper, M.J.; Haluschak, J.J.; Johnson, D.; Schwartz, S.; Morrison, L.J.; Lippa, M.; Hatzivassiliou, G.; Tan, J. P53 mutations in bladder carcinoma cell lines. Oncol. Res. 1994, 6, 569–579. [Google Scholar]
- Brosh, R.; Rotter, V. When mutants gain new powers: News from the mutant p53 field. Nat. Rev. Cancer 2009, 9, 701–713. [Google Scholar]
- Wulfing, C.; Eltze, E.; von Struensee, D.; Wulfing, P.; Hertle, L.; Piechota, H. Cyclooxygenase-2 expression in bladder cancer: Correlation with poor outcome after chemotherapy. Eur. Urol. 2004, 45, 46–52. [Google Scholar] [CrossRef]
- Wang, W.; Adachi, M.; Kawamura, R.; Sakamoto, H.; Hayashi, T.; Ishida, T.; Imai, K.; Shinomura, Y. Parthenolide-induced apoptosis in multiple myeloma cells involves reactive oxygen species generation and cell sensitivity depends on catalase activity. Apoptosis 2006, 11, 2225–2235. [Google Scholar] [CrossRef]
- Yao, H.; Tang, X.; Shao, X.; Feng, L.; Wu, N.; Yao, K. Parthenolide protects human lens epithelial cells from oxidative stress-induced apoptosis via inhibition of activation of caspase-3 and caspase-9. Cell Res. 2007, 17, 565–571. [Google Scholar] [CrossRef]
- Hidalgo, M.; Eckhardt, S.G. Development of matrix metalloproteinase inhibitors in cancer therapy. J. Natl. Cancer Inst. 2001, 93, 178–193. [Google Scholar] [CrossRef]
- Liu, J.W.; Cai, M.X.; Xin, Y.; Wu, Q.S.; Ma, J.; Yang, P.; Xie, H.Y.; Huang, D.S. Parthenolide induces proliferation inhibition and apoptosis of pancreatic cancer cells in vitro. J. Exp. Clin. Cancer Res. 2010, 29, 108. [Google Scholar]
- Curry, E.A., 3rd.; Murry, D.J.; Yoder, C.; Fife, K.; Armstrong, V.; Nakshatri, H.; O'Connell, M.; Sweeney, C.J. Phase i dose escalation trial of feverfew with standardized doses of parthenolide in patients with cancer. Invest. New Drugs 2004, 22, 299–305. [Google Scholar] [CrossRef]
- Neelakantan, S.; Nasim, S.; Guzman, M.L.; Jordan, C.T.; Crooks, P.A. Aminoparthenolides as novel anti-leukemic agents: Discovery of the nf-kappab inhibitor, dmapt (lc-1). Bioorg. Med. Chem. Lett. 2009, 19, 4346–4349. [Google Scholar] [CrossRef]
- Samples Availability: Samples of the compounds are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cheng, G.; Xie, L. Parthenolide Induces Apoptosis and Cell Cycle Arrest of Human 5637 Bladder Cancer Cells In Vitro. Molecules 2011, 16, 6758-6768. https://doi.org/10.3390/molecules16086758
Cheng G, Xie L. Parthenolide Induces Apoptosis and Cell Cycle Arrest of Human 5637 Bladder Cancer Cells In Vitro. Molecules. 2011; 16(8):6758-6768. https://doi.org/10.3390/molecules16086758
Chicago/Turabian StyleCheng, Guang, and Liping Xie. 2011. "Parthenolide Induces Apoptosis and Cell Cycle Arrest of Human 5637 Bladder Cancer Cells In Vitro" Molecules 16, no. 8: 6758-6768. https://doi.org/10.3390/molecules16086758