Molecular Properties of Water-Unextractable Proteoglycans from Hypsizygus marmoreus and Their in Vitro Immunomodulatory Activities


Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Composition Analysis
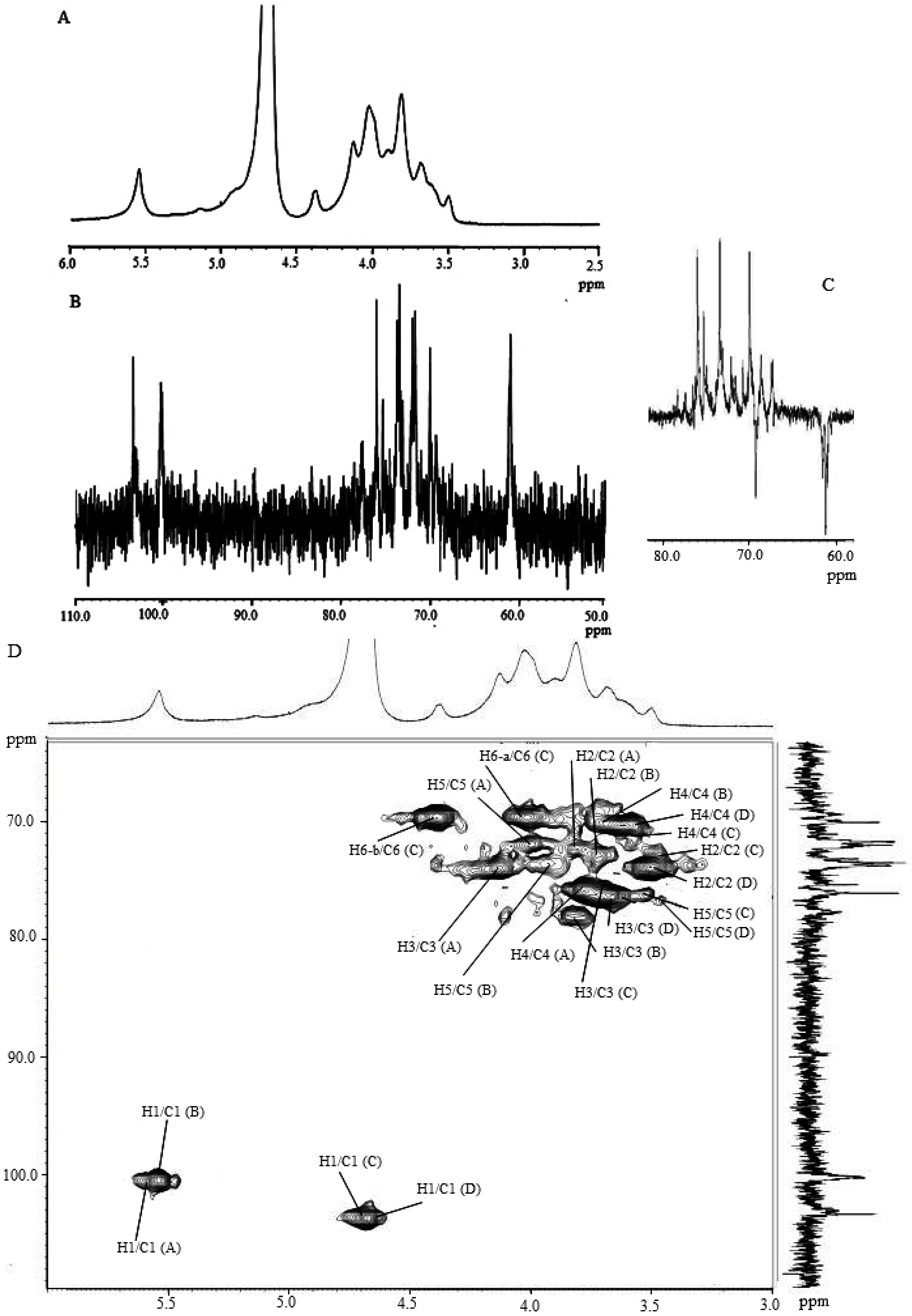
2.2. Molecular Characteristics of Proteoglycans


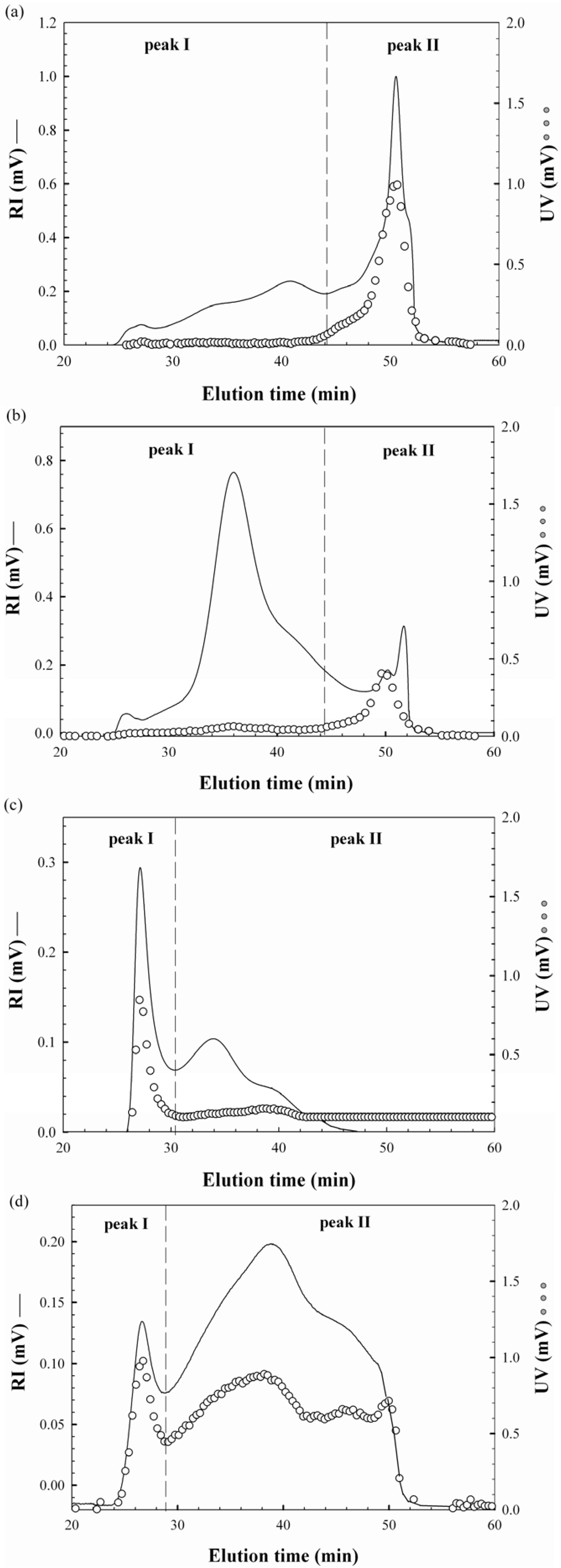{kind=link}
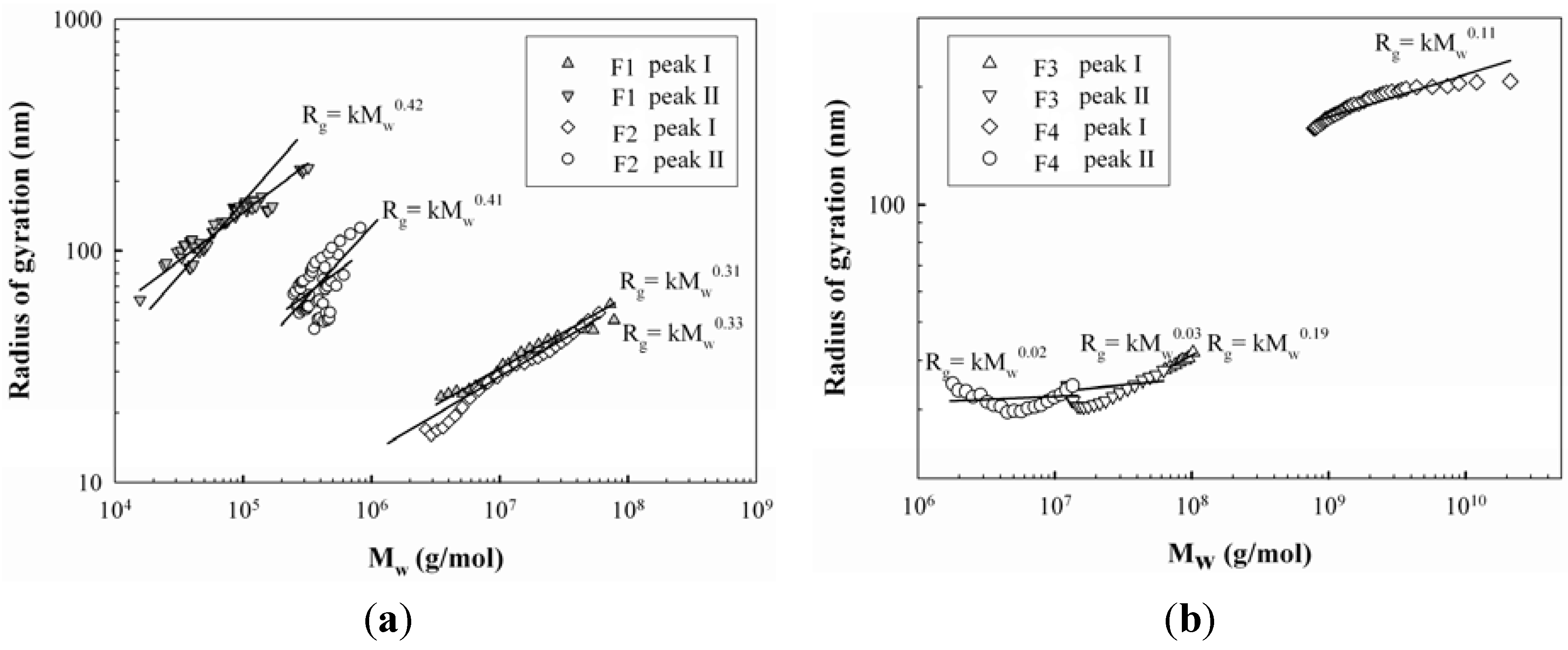{kind=link}
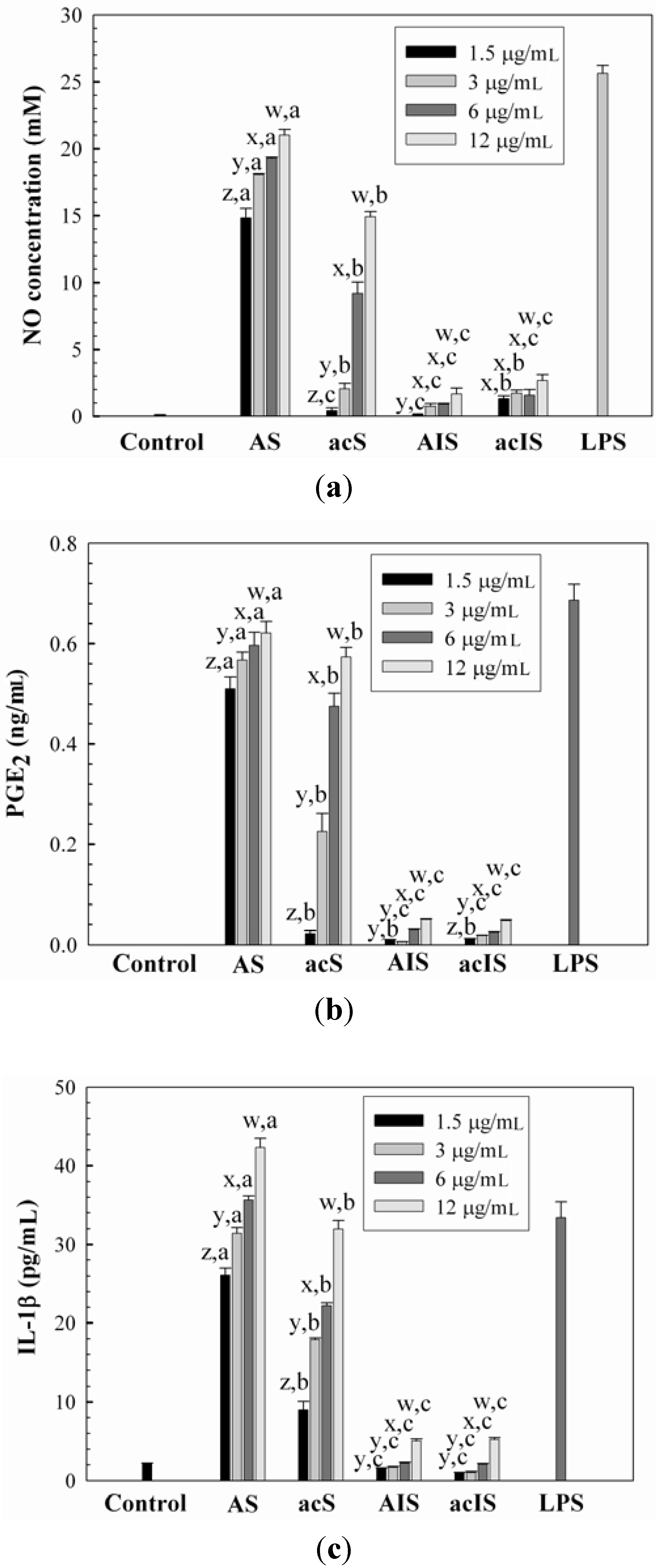{kind=link}
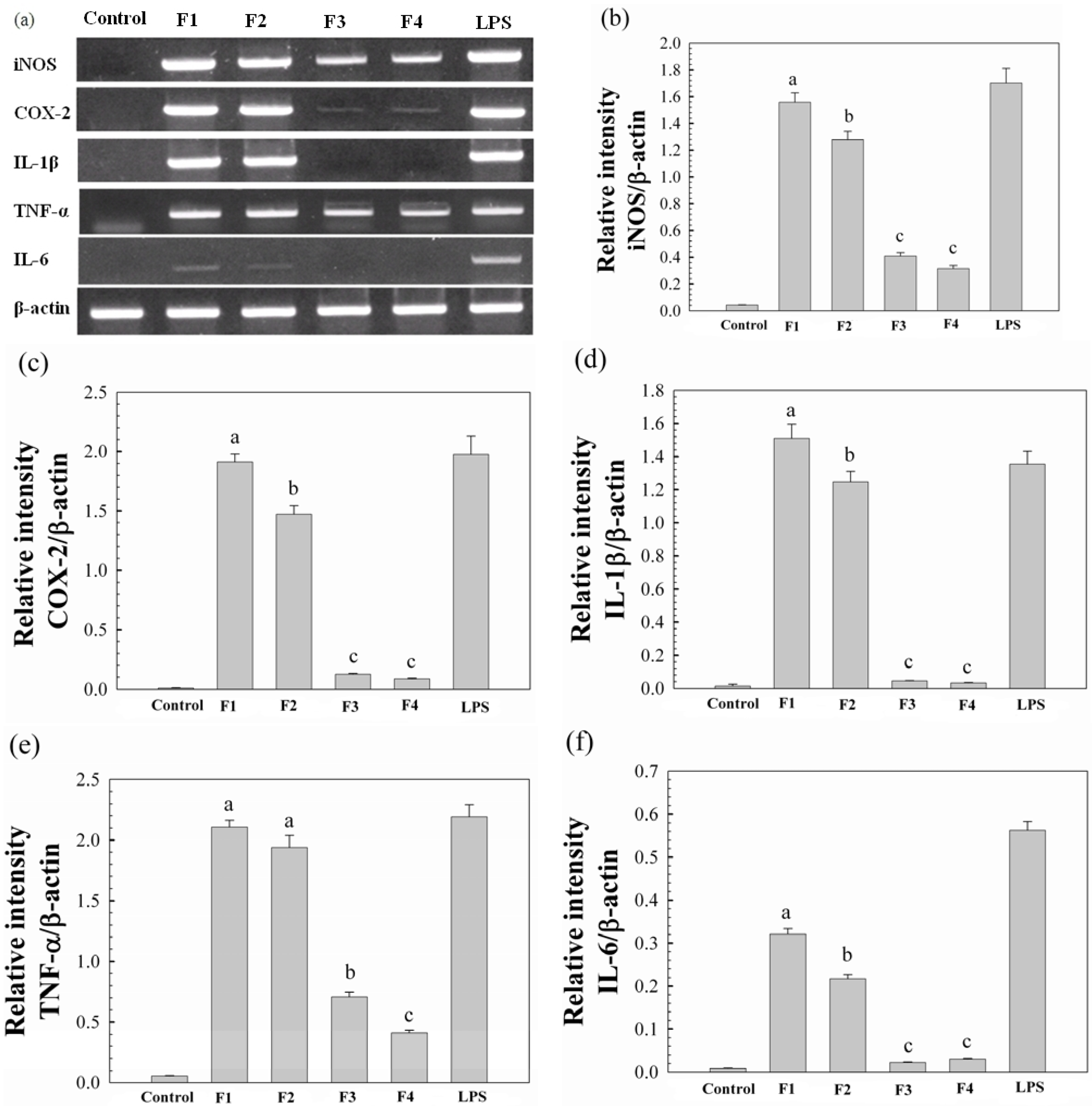{kind=link}
{kind=link}
{kind=link}
| Sample | Ratio (%) | Mw × 104 (g/mol) | Rg (nm) | SVg (cm3/g) | ||||
|---|---|---|---|---|---|---|---|---|
| Peak I | Peak II | Peak I | Peak II | Peak I | Peak II | Peak I | Peak II | |
| F1 | 48.0 | 52.0 | 753 ± 8 | 16 ± 1 | 73 ± 1 | 136 ± 1 | 0.133 ± 0.005 | 40.5 ± 2.06 |
| F2 | 82.0 | 18.0 | 259 ± 4 | 25 ± 1 | 50 ± 1 | 148 ± 6 | 0.124 ± 0.008 | 32.8 ± 1.66 |
| F3 | 41.1 | 58.9 | 8428 ± 48 | 1797 ± 18 | 37 ± 0 | 35 ± 1 | 0.002 ± 0.000 | 0.006 ± 0.000 |
| F4 | 10.0 | 90.0 | 19,545 ± 98 | 444 ± 5 | 123 ± 6 | 40 ± 2 | 0.024 ± 0.001 | 0.037 ± 0.001 |

2.3. Immunomodulatory Activity of Water-Unextractable Proteoglycans

2.4. Glycosidic Linkage Analysis of F1 and F2

| Methylated sugars a | Retention time (min) | Mass fragmentation (m/z) | Glycosidic linkage | Peak area (%) | |
|---|---|---|---|---|---|
| F1 | F2 | ||||
| 2,3,4,6-Me4-Glc | 9.64 | 43, 59, 71, 87, 102, 118, 129, 145, 162, 174, 205 | Glc-(1→ | 12.9 | 21.2 |
| 2,4,6-Me3-Glc | 11.29 | 43, 59, 71, 87, 101, 118, 129, 161, 234 | →3)-Glc-(1→ | 8.7 | 29.0 |
| 2,3,6-Me3-Glc | 11.47 | 43, 59, 71, 87, 102, 118, 129, 142, 162, 173, 233 | →4)-Glc-(1→ | 57.0 | 19.1 |
| 2,3,4-Me3-Glc | 11.77 | 43, 59, 71, 87, 102, 118, 129, 143, 162, 173, 189, 233 | →6)-Glc-(1→ | 8.4 | 10.2 |
| 2,3-Me2-Glc | 13.44 | 43, 59, 74, 85, 102, 118, 127, 142, 162, 201, 261 | →4,6)-Glc-(1→ | 6.2 | 2.9 |
| 2,4-Me2-Glc | 13.62 | 43, 59, 74, 87, 101, 118, 129, 139, 160, 174, 189, 234 | →3,6)-Glc-(1→ | 6.8 | 17.6 |
| 2,3,4-Me3-Gal | 12.26 | 43, 59, 71, 87, 102, 118, 129, 142, 162, 173, 189, 233 | →6)-Gal-(1→ | nd | nd |
| Sample | Residue | C-1/H-1 | C-2/H-2 | C-3/H-3 | C-4/H-4 | C-5/H-5 | C-6/H-6a, H-6b |
|---|---|---|---|---|---|---|---|
| F1 | A α (1→4) Glu | 100.2/5.52 | 71.9/3.81 | 73.7/4.13 | 75.3/3.77 | 71.7/3.99 | 61.0/3.89, 4.05 |
| B α (1→3) Glu | 100.0/5.51 | 72.1/3.72 | 77.7/3.81 | 70.0/3.60 | 73.1/3.90 | 61.3/3.88, 4.00 | |
| C β (1→6) Glu | 103.1/4.68 | 74.0/3.47 | 76.1/3.71 | 70.0/3.58 | 76.3/3.52 | 69.3/4.01, 4.37 | |
| D β (1→) Glu | 103.3/4.68 | 73.8/3.49 | 76.0/3.67 | 70.0/3.58 | 76.6/3.45 | 60.9/3.51, 3.85 | |
| F2 | A α (1→4) Glu | 100.2/5.53 | 71.9/3.80 | 73.7/4.12 | 76.1/3.75 | 71.6/3.99 | 61.0/3.89, 4.05 |
| B α (1→3) Glu | 100.0/5.52 | 72.4/3.73 | 77.5/3.80 | 70.2/3.60 | 73.5/3.90 | 61.3/3.88, 4.00 | |
| C β (1→6) Glu | 103.1/4.68 | 74.0/3.55 | 76.1/3.69 | 70.0/3.58 | 76.3/3.52 | 69.3/4.01, 4.37 | |
| D β (1→) Glu | 103.3/4.67 | 73.9/3.49 | 76.3/3.67 | 70.0/3.58 | 76.5/3.5 | 60.9/3.51, 3.85 |

3. Experimental
3.1. Materials
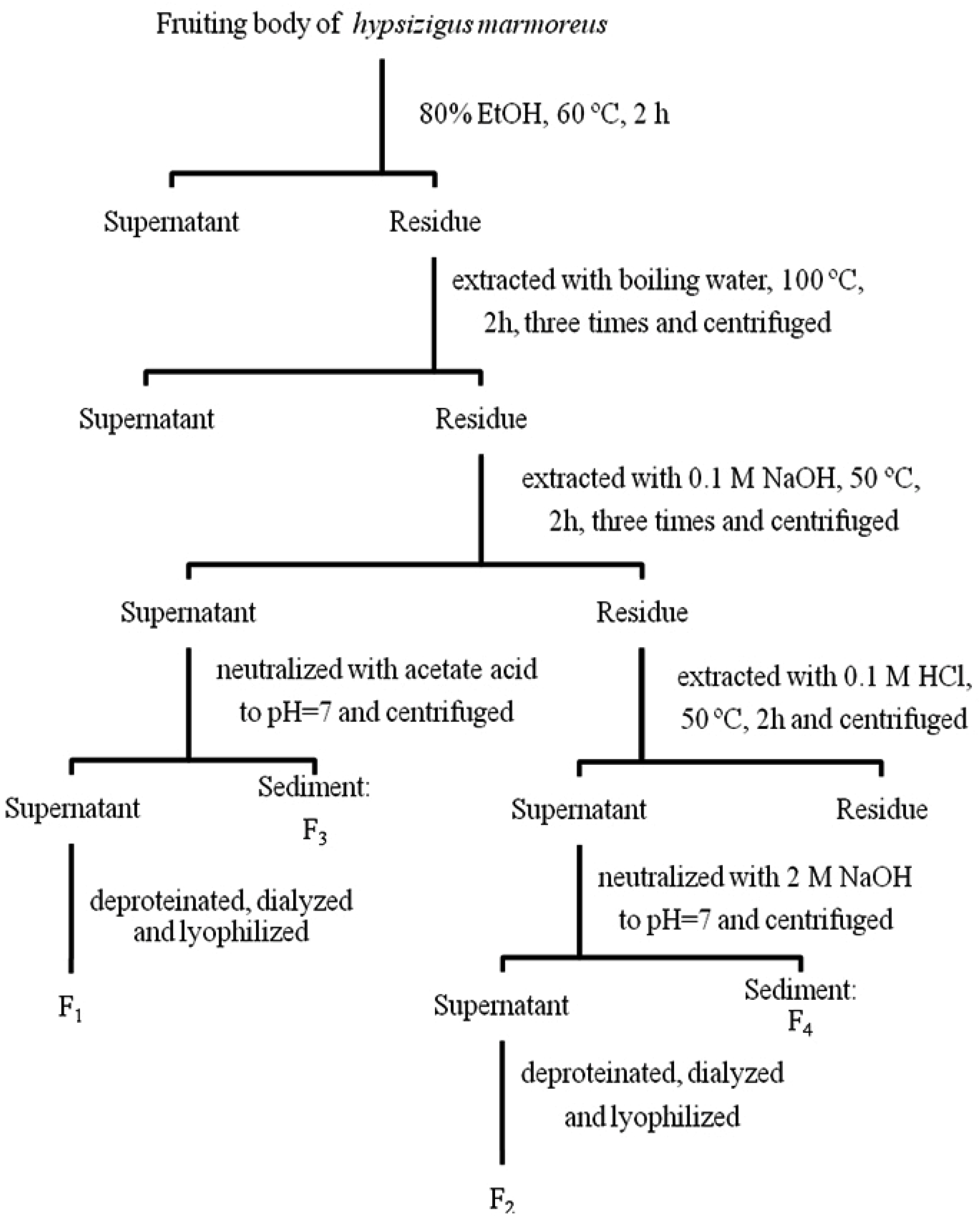
3.2. Preparation of the Alkali and Acid Extractable Proteoglycans

3.3. Determination of the Total Carbohydrate, Protein and Monosaccharide Composition of the Proteoglycans
3.4. Glycosidic Linkage Analysis
3.5. Nuclear Magnetic Resonance (NMR) Spectroscopy
3.6. Determination of the Weight Average Molecular Weight
3.7. Assays for Macrophage Proliferation
3.8. Nitric Oxide Production
3.9. Assay for Cytokine Secretion
3.10. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
3.11. Statistical Analyses
4. Conclusions
Acknowledgment
References and Notes
- Sasaki, T.; Takasuka, N. Further study of the structure of lentinan, an anti-tumor polysaccharide from Lentinus edodes. Carbohydr. Res. 1976, 47, 99–104. [Google Scholar] [CrossRef]
- Komatsu, N.; Okubo, S.; Kikumoto, S.; Kimura, K.; Saito, G.; Sasaki, S. Host mediated antitumor action of schizophyllan, a glucan produced by Schizophyllum commune. Gann 1971, 60, 557–563. [Google Scholar]
- Tsugagoshi, S.; Ohash, F. Protein-bound polysaccharides preparation, PS-K. Protein-bound polysaccharide preparation, PS-K, effective against mouse sarcoma-180 and rat ascites hepatoma AH-13 by oral use. Gann 1974, 65, 557–565. [Google Scholar]
- Wang, H.X.; Liu, W.K.; Ng, T.B.; Ooi, V.E.C.; Chang, S.T. Immunomodulatory and antitumor activities of a polysaccharide-peptide complex from a mycelial culture of Tricholoma sp., a local edible mushroom. Life Sci. 1995, 57, 269–281. [Google Scholar]
- Lavi, I.; Friesem, D.; Geresh, S.; Hadar, Y.; Schwartz, B. An aqueous polysaccharide extract from the edible mushroom Pleurotus ostreatus induces anti proliferative and pro-apoptotic effects on HT-29 colon cancer cells. Cancer Lett. 2006, 244, 61–70. [Google Scholar] [CrossRef]
- Wong, S.M.; Wong, K.K.; Chiu, L.C.M.; Cheung, C.K. Non-starch polysaccharides form different developmental stages of Pleurotus tuber-regium inhibited the growth of human acute promyelocytic leukemia HL-60 cells by cell-cycle arrest an/or apoptotic induction. Carbohydr. Polym. 2007, 68, 206–217. [Google Scholar] [CrossRef]
- Surenjav, U.; Zhang, L.; Xu, X.; Zhagn, X.; Zeng, F. Effects of molecular structure on antitumor activities of (1→3)-β-D-glucans from different Lentinus Edodes. Carbohydr. Polym. 2006, 63, 97–104. [Google Scholar] [CrossRef]
- Demleitner, S.; Kraus, J.; Franz, G. Synthesis and antitumor activity of sulfoalkyl derivatives of curdlan and lichenan. Carbohydr. Res. 1992, 226, 247–252. [Google Scholar]
- Mizuno, T. The extraction and development of antitumor active polysaccharides from medicinal mushrooms in Japan. Int. J. Med. Mushrooms 1999, 1, 9–30. [Google Scholar]
- Peng, Y.; Zhang, L.; Zeng, F.; Kennedy, J. Structure and antitumor activities of the water-soluble polysaccharides from Ganoderma tsugae mycelium. Carbohydr. Polym. 2005, 59, 385–392. [Google Scholar]
- Maeda, Y.Y.; Watanabe, S.T.; Chihara, C.; Rokunada, M. Denaturation and renaturation of β-1,6; 1,3-glucan, Lentinan, for the expression of T-cell mediated responses. Cancer Res. 1988, 48, 671–675. [Google Scholar]
- Mizuno, T.; Yeohlui, T.; Kinoshita, T.; Zhuang, C.; Ito, H.; Mayuzumi, Y. Antitumor activity and chemical modification of polysaccharides from niohshimeji mushroom, Tricholma giganteum. Biosci. Biotechnol. Biochem. 1996, 60, 30–33. [Google Scholar]
- Mizuno, T.; Kato, N.; Totsuka, A.; Takenaka, K.; Shinakai, K.; Shimizu, M. Fractionation, structural features and antitumor activity of water-soluble polysaccahrides from “Reishi”, the fruit body of Ganoderma lucidum. J. Agric. Chem. Soc. Jpn. 1984, 58, 871–880. [Google Scholar]
- Mizuno, T.; Suuzki, E.; Maki, K.; Tamaki, H. Fractionation, chemical modification and antitumor activity of water-insoluble polysaccharides of the fruiting body of Ganoderma lucidum. J. Agric. Chem. Soc. Jpn. 1985, 59, 1143–1151. [Google Scholar]
- Bao, H.H.; Choi, W.S.; You, S.G. Effect of sulfated modification on the molecular characteristics and biological activities of polysaccharides from Hypsizygus marmoreus. Biosci. Biotechnol. Biochem. 2010, 74, 1408–1414. [Google Scholar] [CrossRef]
- Hromadkova, Z.; Ebringerova, A.; Valachovic, P. Comparison of classical and ultrasound-assisted extraction of polysaccharides from Salvia officinalis L. Ultrason. Sonochem. 1999, 5, 163–168. [Google Scholar] [CrossRef]
- Lindsey, J.W. An alkali-soluble factor present in normal brain tissue inhibits antigen-specific lymphocyte proliferation. J. Neuroimmunol. 2000, 103, 76–83. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, L.; Wang, Y.; Cheung, P.C.K. Chain conformation of sulfated derivatives of β-glucan from sclerotia of Pleurotus tuber-regium. Carbohydr. Res. 2003, 338, 2863–2870. [Google Scholar] [CrossRef]
- Synytsya, A.; Mickova, K.; Synytsya, A.; Jablonsky, I.; Spevacek, J.; Erban, V.; Kovarikova, E.; Copikova, J. Glucans from fruit bodies of cultivated mushrooms Pleurotus ostreatus and Pleurotus eryngii: Structure and potential prebiotic activity. Carbohydr. Polym. 2009, 76, 548–556. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Agafonova, S.V.; Borovskii, G.B.; Penzina, T.A.; Rokhin, A.V. Alkali-soluble polysaccharides of Laetiporus Sulphureus (Bull.: Fr.) Murr fruit bodies. Appl. Biochem. Microbiol. 2009, 45, 693–697. [Google Scholar]
- You, S.G.; Lim, S.T. Molecular characterization of corn starch using an aqueous HPSEC-MALLS-RI system under various dissolution and analytical conditions. Cereal Chem. 2000, 77, 303–308. [Google Scholar] [CrossRef]
- Bandtlow, C.E.; Zimmermann, D.R. Proteoglycans in the developing brain: New conceptual insight for old proteins. Physiol. Rev. 2000, 80, 1267–1290. [Google Scholar]
- Porcheray, F.; Viaud, S.; Rimaniol, A.C.; Leone, C.; Samah, B.; Dereuddre-Bosquet, N.; Dormont, D.; Gras, G. Macrophage activation switching: An asset for the resolution of inflammation. Clin. Exp. Immunol. 2005, 142, 481–489. [Google Scholar]
- Jiao, L.; Li, X.; Li, T.; Jiang, P.; Zhang, L.; Wu, M.; Zhang, L. Characterization and anti-tumor activity of alkali-extracted polysaccharide from Enteromorpha intestinalis. Int. Immunopharmacol. 2009, 9, 324–329. [Google Scholar] [CrossRef]
- Yadomae, T.; Ohno, N. Structure-activity relationship of immunomodulating (1,3)-β-D-glucans. Recent Res. Dev. Chem. Pharm. Sci. 1996, 1, 23–33. [Google Scholar]
- You, S.G.; Yang, C.; Lee, H.Y.; Lee, B.Y. Molecular characteristics of partially hydrolyzed fucoidans from sporophyll of Undaria Pinnatida and their in vitro anticancer activity. Food Chem. 2010, 119, 554–559. [Google Scholar] [CrossRef]
- Liu, C.; Xi, T.; Lin, Q.; Xing, Y.; Ye, L.; Luo, X.; Wang, F. Immunomodulatory activity of polysaccharides isolated from Strongylocentrotus nudus eggs. Int. Immunopharmacol. 2008, 8, 1835–1841. [Google Scholar] [CrossRef]
- Na, Y.S.; Kim, W.J.; Kim, S.M.; Park, F.K.; Lee, S.M.; Kim, S.O.; Synytsya, A.; Park, Y.I. Purification, characterization and immunostimulating activity of water-soluble polysaccharide isolated from Capsosiphon fulvescens. Int. Immunopharmacol. 2010, 10, 364–370. [Google Scholar]
- Hakomori, S.I. A rapid permethylation of glycolipid, and polysaccharide catalyzed by methylsulfinyl carbanion in dimethyl sulfoxide. J.Biochem. 1964, 55, 205–208. [Google Scholar]
- Cui, F.J.; Tao, W.Y.; Xu, Z.H.; Guo, W.J.; Xu, H.Y.; Ao, Z.H.; Jin, J.; Wei, Y.Q. Structural analysis of anti-tumor heteropolysaccharide GFPS1b from the cultured mycelia of Grifola frondosa GF9801. Bioresour. Technol. 2007, 98, 395–401. [Google Scholar]
- Agrawal, P.K. NMR spectroscopy in the structural elucidation of oligosaccharides and glycosides. Phytochemistry 1992, 31, 3307–3330. [Google Scholar]
- Maeda, Y.Y.; Chihara, G. Lentinan and other antitumoral polysaccharides. In Immunomodulatory Agentsform Plants; Wagner, H., Ed.; Birkhauser Verlag: Basel, Switzerland, 1999; pp. 203–221. [Google Scholar]
- Sevag, M.G.; Lackman, D.B.; Smolens, J. The isolation of the components of streptococcal nucleoproteins in serologically active form. J. Biol. Chem. 1938, 124, 425–436. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin-Phenol reagents. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Bao, H.; You, S.G. Molecular characteristics of water-soluble extracts from Hipsizigus marmoreus and their in vitro growth inhibition of various cancer cell lines and immunomodulatory function in Raw264.7 cells. Biosci. Biotechnol. Biochem. 2011, 75, 891–898. [Google Scholar] [CrossRef]
- Green, L.C.; Wanger, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–136. [Google Scholar]
- Sample Availability: Samples of the mushroom polysaccharides are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bao, H.H.; Tarbasa, M.; Chae, H.M.; You, S.G. Molecular Properties of Water-Unextractable Proteoglycans from Hypsizygus marmoreus and Their in Vitro Immunomodulatory Activities. Molecules 2012, 17, 207-226. https://doi.org/10.3390/molecules17010207
Bao HH, Tarbasa M, Chae HM, You SG. Molecular Properties of Water-Unextractable Proteoglycans from Hypsizygus marmoreus and Their in Vitro Immunomodulatory Activities. Molecules. 2012; 17(1):207-226. https://doi.org/10.3390/molecules17010207
Chicago/Turabian StyleBao, Hong Hui, Mehdi Tarbasa, Hee Mun Chae, and Sang Guan You. 2012. "Molecular Properties of Water-Unextractable Proteoglycans from Hypsizygus marmoreus and Their in Vitro Immunomodulatory Activities" Molecules 17, no. 1: 207-226. https://doi.org/10.3390/molecules17010207
APA StyleBao, H. H., Tarbasa, M., Chae, H. M., & You, S. G. (2012). Molecular Properties of Water-Unextractable Proteoglycans from Hypsizygus marmoreus and Their in Vitro Immunomodulatory Activities. Molecules, 17(1), 207-226. https://doi.org/10.3390/molecules17010207