The Protection of Selenium against Cadmium-Induced Cytotoxicity via the Heat Shock Protein Pathway in Chicken Splenic Lymphocytes

Abstract
:1. Introduction
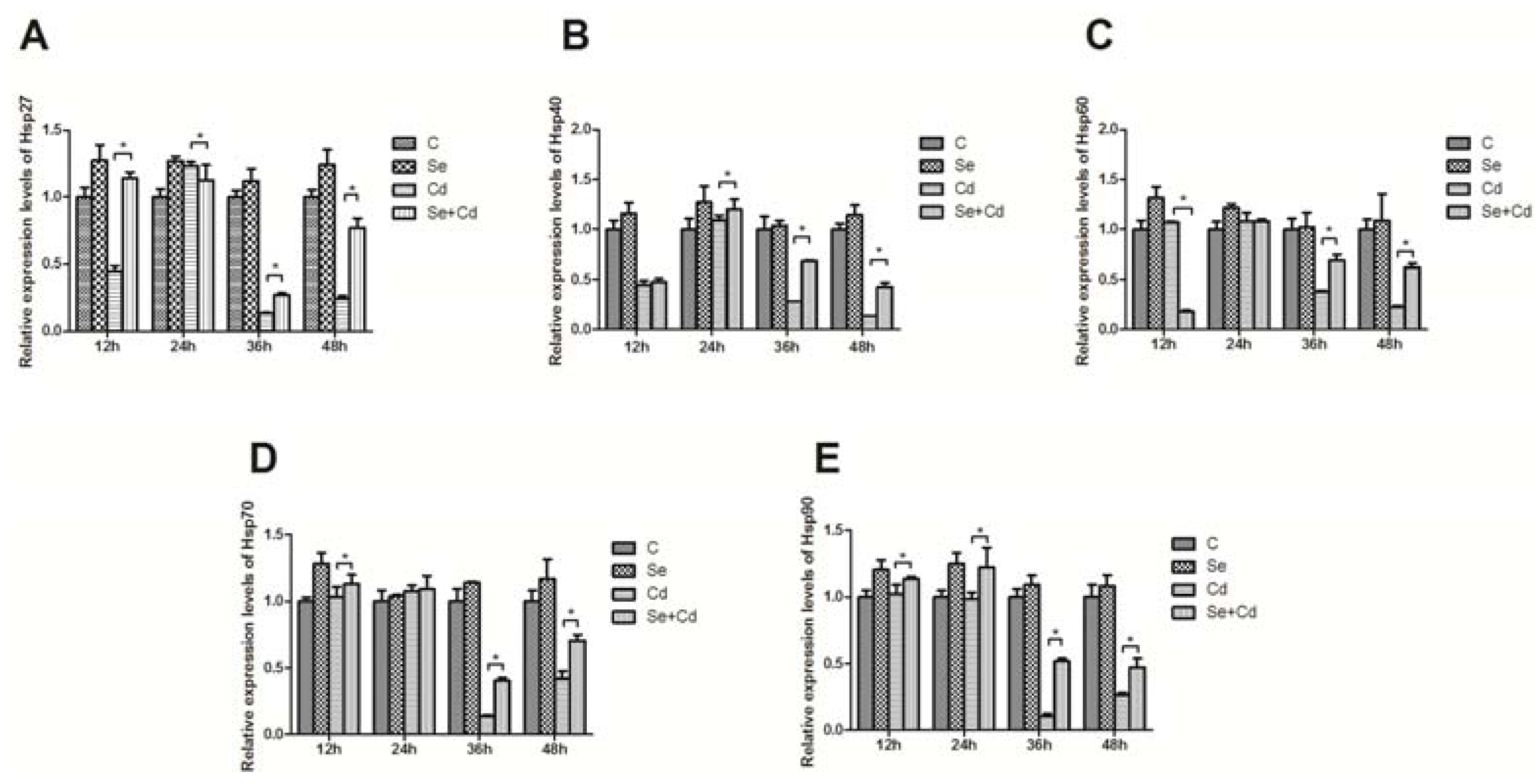
2. Results and Discussion

3. Experimental
3.1. Preparation of Chicken Splenic Lymphocytes Suspension and Treatment
3.2. Quantification of HSPs mRNA

{kind=link}
| Gene | Accession number | Primer (5′→3′) | Product size (bp) |
|---|---|---|---|
| HSP27 | NM_205290 | 5'-ACACGAGGAGAAACAGGATGAG-3' | 158 bp |
| 5'-ACTGGATGGCTGGCTTGG-3' | |||
| HSP40 | NM_001199325 | 5'-GGGCATTCAACAGCATAGA-3' | 151 bp |
| 5'-TTCACATCCCCAAGTTTAGG-3' | |||
| HSP60 | NM_001012916 | 5'-AGCCAAAGGGCAGAAATG-3' | 208 bp |
| 5'-TACAGCAACAACCTGAAGA-3' | |||
| HSP70 | NM_001006685 | 5'-CGGGCAAGTTTGACCTAA-3 | 250 bp |
| 5'-TTGGCTCCCACCCTATCTCT-3' | |||
| HSP90 | NM_001109785 | 5'-TCCTGTCCTGGCTTTAGTTT-3' | 143 bp |
| 5'-AGGTGGCATCTCCTCGGT-3' | |||
| β-actin | L08165 | 5'-CCGCTCTATGAAGGCTACGC-3' | 128 bp |
| 5'-CTCTCGGCTGTGGTGGTGAA-3' |
3.3. Statistical Analyses
4. Conclusions
Acknowledgments
- Sample Availability: Samples of the Se and Cd are available from the authors.
References
- Mah, V.; Jalilehvand, F. Cadmium(II) complex formation with glutathione. J. Biol. Inorg. Chem. 2010, 15, 441–458. [Google Scholar] [CrossRef]
- Pathak, N.; Khandelwal, S. Influence of cadmium on murine thymocytes: Potentiation of apoptosis and oxidative stress. Toxicol. Lett. 2006, 165, 121–132. [Google Scholar] [CrossRef]
- Pathak, N.; Khandelwal, S. Role of oxidative stress and apoptosis in cadmium induced thymic atrophy and splenomegaly in mice. Toxicol. Lett. 2007, 169, 95–108. [Google Scholar] [CrossRef]
- Pathak, N.; Khandelwal, S. Impact of cadmium in T lymphocyte subsets and cytokine expression: Differential regulation by oxidative stress and apoptosis. Biometals 2008, 21, 179–187. [Google Scholar] [CrossRef]
- Pathak, N.; Khandelwal, S. Oxidative stress and apoptotic changes in murine splenocytes exposed to cadmium. Toxicology 2006, 220, 26–36. [Google Scholar] [CrossRef]
- Pathak, N.; Khandelwal, S. Cytoprotective and immunomodulating properties of piperine on murine splenocytes: An in vitro study. Eur. J. Pharmacol. 2007, 576, 160–170. [Google Scholar] [CrossRef]
- Planello, R.; Martinez-Guitarte, J.L.; Morcillo, G. Effect of acute exposure to cadmium on the expression of heat-shock and hormone-nuclear receptor genes in the aquatic midge Chironomus riparius. Sci. Total Environ. 2010, 408, 1598–1603. [Google Scholar] [CrossRef]
- Yu, X.; Hong, S.; Faustman, E.M. Cadmium-induced activation of stress signaling pathways, disruption of ubiquitin-dependent protein degradation and apoptosis in primary rat Sertoli cell-gonocyte cocultures. Toxicol. Sci. 2008, 104, 385–396. [Google Scholar]
- Shilo, S.; Pardo, M.; Aharoni-Simon, M.; Glibter, S.; Tirosh, O. Selenium supplementation increases liver MnSOD expression: Molecular mechanism for hepato-protection. J. Inorg. Biochem. 2008, 102, 110–118. [Google Scholar] [CrossRef]
- Brigelius-Flohe, R.; Flohe, L. Is there a role of glutathione peroxidases in signaling and differentiation. Biofactors 2003, 17, 93–102. [Google Scholar] [CrossRef]
- Combs, G.F., Jr; Midthune, D.N.; Patterson, K.Y.; Canfield, W.K.; Hill, A.D.; Levander, O.A.; Taylor, P.R.; Moler, J.E.; Patterson, B.H. Effects of selenomethionine supplementation on selenium status and thyroid hormone concentrations in healthy adults. Am. J. Clin. Nutr. 2009, 89, 1808–1814. [Google Scholar] [CrossRef]
- Bansal, M.P.; Kaur, P. Selenium, a versatile trace element: Current research implications. Indian J. Exp. Biol. 2005, 43, 1119–1129. [Google Scholar]
- El-Sharaky, A.S.; Newairy, A.A.; Badreldeen, M.M.; Eweda, S.M.; Sheweita, S.A. Protective role of selenium against renal toxicity induced by cadmium in rats. Toxicology 2007, 235, 185–193. [Google Scholar] [CrossRef]
- Lazarus, M.; Orct, T.; Jurasoviae, J.; Blanusa, M. The effect of dietary selenium supplementation on cadmium absorption and retention in suckling rats. Biometals 2009, 22, 973–983. [Google Scholar] [CrossRef]
- Rivera, R.E.; Christensen, V.L.; Edens, F.W.; Wineland, M.J. Influence of selenium on heat shock protein 70 expression in heat stressed turkey embryos (Meleagris gallopavo). Comp. Biochem. Phys. A 2005, 142, 427–432. [Google Scholar] [CrossRef]
- Gabriel, J.E.; da, M.A.F.; Boleli, I.C.; Macari, M.; Coutinho, L.L. Effect of moderate and severe heat stress on avian embryonic hsp70 gene expression. Growth Dev. Aging 2002, 66, 27–33. [Google Scholar]
- Hassen, W.; El, G.E.; Baudrimont, I.; Mobio, A.T.; Ladjimi, M.M.; Creppy, E.E.; Bacha, H. Cytotoxicity and Hsp 70 induction in Hep G2 cells in response to zearalenone and cytoprotection by sub-lethal heat shock. Toxicology 2005, 207, 293–301. [Google Scholar] [CrossRef]
- Somji, S.; Ann, S.M.; Garrett, S.H.; Gurel, V.; Todd, J.H.; Sens, D.A. Expression of hsp 90 in the human kidney and in proximal tubule cells exposed to heat, sodium arsenite and cadmium chloride. Toxicol. Lett. 2002, 133, 241–254. [Google Scholar] [CrossRef]
- Simpson, S.A.; Alexander, D.J.; Reed, C.J. Induction of heat shock protein 70 in rat olfactory epithelium by toxic chemicals: In vitro and in vivo studies. Arch. Toxicol. 2005, 79, 224–230. [Google Scholar] [CrossRef]
- Kultz, D. Molecular and evolutionary basis of the cellular stress response. Annu. Rev. Physiol. 2005, 67, 225–257. [Google Scholar] [CrossRef]
- Sieck, G.C. Molecular biology of thermoregulation. J. Appl. Physiol. 2002, 92, 1365–1366. [Google Scholar]
- Freeman, M.L.; Borrelli, M.J.; Meredith, M.J.; Lepock, J.R. On the path to the heat shock response: destabilization and formation of partially folded protein intermediates, a consequence of protein thiol modification. Free Radic. Biol. Med. 1999, 26, 737–745. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, Z.; Jian, J.; Lu, Y. Cloning and expression of heat shock protein 70 gene in the haemocytes of pearl oyster (Pinctada fucata, Gould 1850) responding to bacterial challenge. Fish Shellfish Immunol. 2009, 26, 639–645. [Google Scholar] [CrossRef]
- Fu, D.; Chen, J.; Zhang, Y.; Yu, Z. Cloning and expression of a heat shock protein (HSP) 90 gene in the haemocytes of Crassostrea hongkongensis under osmotic stress and bacterial challenge. Fish Shellfish Immunol. 2011, 31, 118–125. [Google Scholar] [CrossRef]
- Werlin, R.; Priester, J.H.; Mielke, R.E.; Kramer, S.; Jackson, S.; Stoimenov, P.K.; Stucky, G.D.; Cherr, G.N.; Orias, E.; Holden, P.A. Biomagnification of cadmium selenide quantum dots in a simple experimental microbial food chain. Nat. Nanotechnol. 2011, 6, 65–71. [Google Scholar] [CrossRef]
- Timblin, C.R.; Janssen, Y.M.; Goldberg, J.L.; Mossman, B.T. GRP78, HSP72/73, and cJun stress protein levels in lung epithelial cells exposed to asbestos, cadmium, or H2O2. Free Radic. Biol. Med. 1998, 24, 632–642. [Google Scholar] [CrossRef]
- Hanson, M.L.; Holaskova, I.; Elliott, M.; Brundage, K.M.; Schafer, R.; Barnett, J.B. Prenatal cadmium exposure alters postnatal immune cell development and function. Toxicol. Appl. Pharmacol. 2012, 261, 196–203. [Google Scholar] [CrossRef]
- Jimi, S.; Uchiyama, M.; Takaki, A.; Suzumiya, J.; Hara, S. Mechanisms of cell death induced by cadmium and arsenic. Ann. NY Acad. Sci. 2004, 1011, 325–331. [Google Scholar]
- Gottschalg, E.; Moore, N.E.; Ryan, A.K.; Travis, L.C.; Waller, R.C.; Pratt, S.; Atmaca, M.; Kind, C.N.; Fry, J.R. Phenotypic anchoring of arsenic and cadmium toxicity in three hepatic-related cell systems reveals compound- and cell-specific selective up-regulation of stress protein expression: Implications for fingerprint profiling of cytotoxicity. Chem. Biol. Interact. 2006, 161, 251–261. [Google Scholar] [CrossRef]
- Thomson, C.D.; Chisholm, A.; McLachlan, S.K.; Campbell, J.M. Brazil nuts: An effective way to improve selenium status. Am. J. Clin. Nutr. 2008, 87, 379–384. [Google Scholar]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef]
- Manikandan, R.; Beulaja, M.; Thiagarajan, R.; Arumugam, M. Effect of curcumin on the modulation of alphaA- and alphaB-crystallin and heat shock protein 70 in selenium-induced cataractogenesis in Wistar rat pups. Mol. Vis. 2011, 17, 388–394. [Google Scholar]
- Karabulut-Bulan, O.; Bolkent, S.; Yanardag, R.; Bilgin-Sokmen, B. The role of vitamin C, vitamin E, and selenium on cadmium-induced renal toxicity of rats. Drug Chem. Toxicol. 2008, 31, 413–426. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Zhang, S.P.; Liu, C.W.; Cai, Y.Q. The protection of selenium on ROS mediated-apoptosis by mitochondria dysfunction in cadmium-induced LLC-PK(1) cells. Toxicol. In Vitro 2009, 23, 288–294. [Google Scholar] [CrossRef]
- Li, J.L.; Gao, R.; Li, S.; Wang, J.T.; Tang, Z.X.; Xu, S.W. Testicular toxicity induced by dietary cadmium in cocks and ameliorative effect by selenium. Biometals 2010, 23, 695–705. [Google Scholar] [CrossRef]
- Li, J.; Li, S.; Li, H.; Tang, Z.; Xu, S.; Wang, X. Oxidative Stress-Mediated Cytotoxicity of Cadmium in Chicken Splenic Lymphocytes. Pol. J. Environ. Stud. 2010, 19, 947–956. [Google Scholar]
- Ruan, H.; Zhang, Z.; Wu, Q.; Yao, H.; Li, J.; Li, S.; Xu, S. Selenium regulates gene expression of selenoprotein W in chicken skeletal muscle system. Biol. Trace Elem. Res. 2012, 145, 59–65. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, X.; Zhu, Y.-H.; Cheng, X.-Y.; Zhang, Z.-W.; Xu, S.-W. The Protection of Selenium against Cadmium-Induced Cytotoxicity via the Heat Shock Protein Pathway in Chicken Splenic Lymphocytes. Molecules 2012, 17, 14565-14572. https://doi.org/10.3390/molecules171214565
Chen X, Zhu Y-H, Cheng X-Y, Zhang Z-W, Xu S-W. The Protection of Selenium against Cadmium-Induced Cytotoxicity via the Heat Shock Protein Pathway in Chicken Splenic Lymphocytes. Molecules. 2012; 17(12):14565-14572. https://doi.org/10.3390/molecules171214565
Chicago/Turabian StyleChen, Xi, Yi-Hao Zhu, Xin-Yue Cheng, Zi-Wei Zhang, and Shi-Wen Xu. 2012. "The Protection of Selenium against Cadmium-Induced Cytotoxicity via the Heat Shock Protein Pathway in Chicken Splenic Lymphocytes" Molecules 17, no. 12: 14565-14572. https://doi.org/10.3390/molecules171214565