Effect of Soluble Sulfide on the Activity of Luminescent Bacteria

Abstract
:
1. Introduction
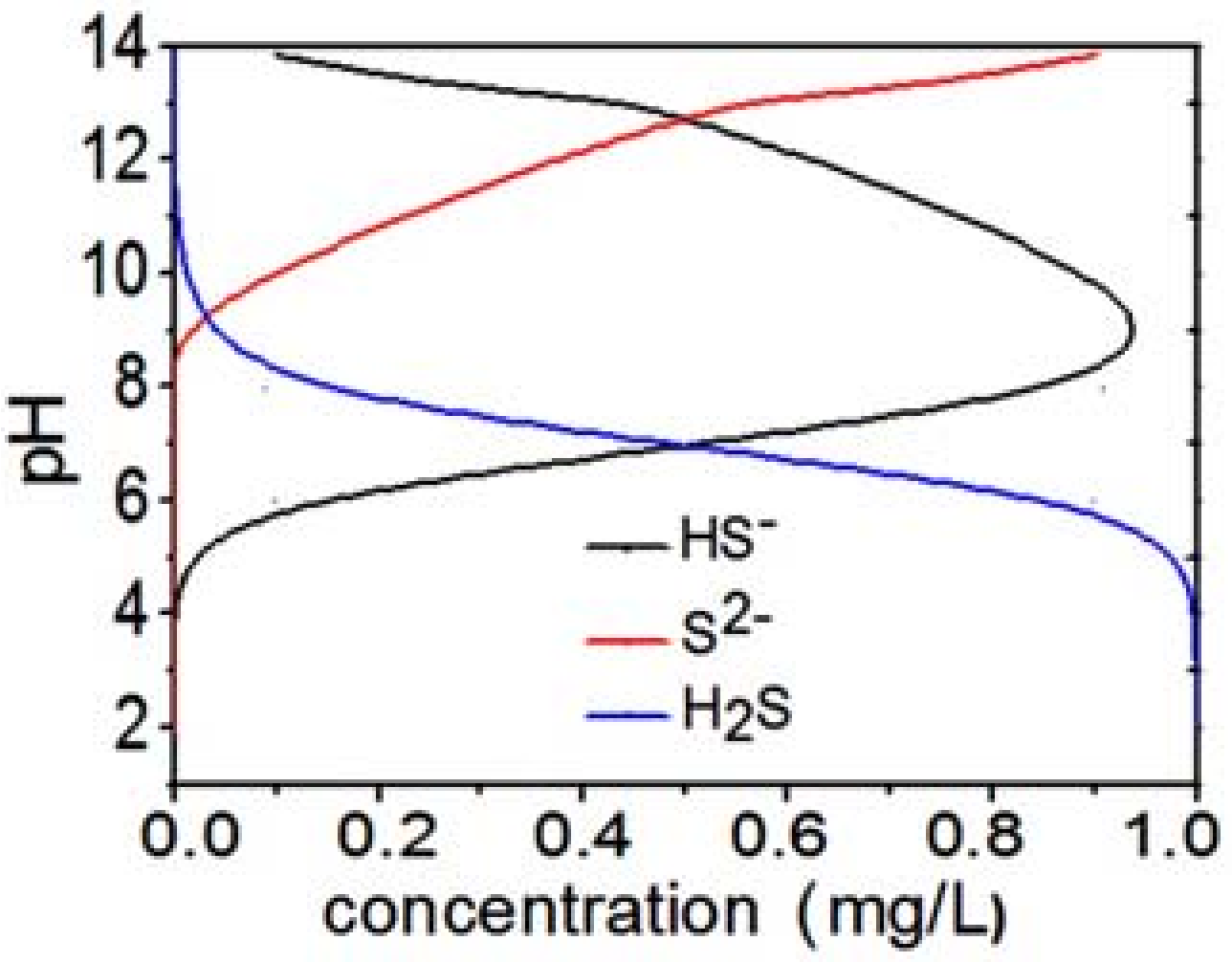
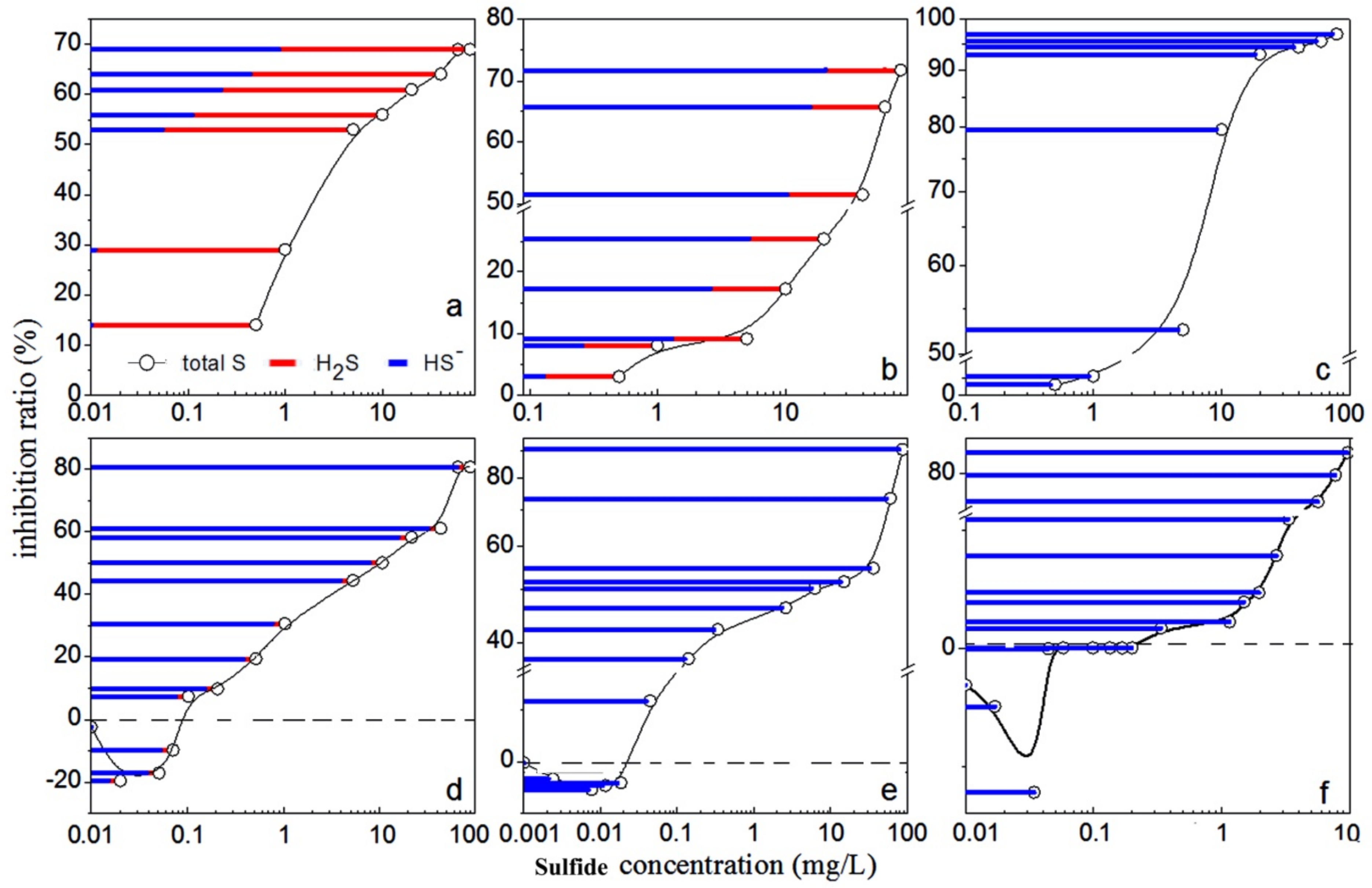
2. Results and Discussion
3. Experimental
3.1. Materials
3.2. Toxicity Tests
4. Conclusions
Acknowledgements
Conflicts of Interest
References and Notes
- Al, T.A.; Martin, C.J.; Blowes, D.W. Carbonate-mineral/water interactions in sulfide-rich mine tailings. Geochim. Cosmochim. Acta 2000, 64, 3933–3948. [Google Scholar] [CrossRef]
- Komori, K.; Miyajima, S.; Tsuru, T.; Fuji, T.; Mohri, S.; Ono, Y.; Sakai, Y. A rapid and simple evaluation system for gas toxicity using luminous bacteria entrapped by a polyion complex membrane. Chemosphere 2009, 77, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Bouillaud, F.; Blachier, F. Mitochondria and sulfide: A very old story of poisoning, feeding, and signaling? Antioxid. Redox. Signal. 2011, 15, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Earson, J.; Havill, D.C. The effect of hypoxia and sulphide on culture-grown wetland and non-wetland plants. II. Metabolic and physiological changes. Exp. Bot. 1988, 39, 431–439. [Google Scholar] [CrossRef]
- Pezhki, S.R.; Pan, S.Z.; Delaune, R.D. Sulfide-induced toxicity Inhibition of carbon assimilation in Spartina alterniflora. Photosynth. Res. 1988, 22, 437–442. [Google Scholar]
- Och, M.S.; Mendelssohn, L.A. Sulphide as a soil phytotoxin: Differential responses in two marsh species. J. Ecol. 1989, 77, 565–578. [Google Scholar]
- Och, M.S.; Mendelssohn, L.A.; Mckee, K.L. Mechanism for the hydrogen sulfide-induced growth limitation in wetland macrophytes. Limnol. Oceanogr. 1990, 35, 399–408. [Google Scholar]
- Prader, A.; Raymond, A.P. Adenine ncleotide ratios and adenylate energy charge in energy metabolism. Ann. Rev. Plant. Physiol. 1983, 34, 199–224. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, H.-J.; Cao, C.; Sun, Y.-Y.; Yang, L.; Wang, B.-Q.; Zhou, J.-G. Inhibition effects of protein-conjugated amorphous zinc sulfide nanoparticles on tumor cells growth. J. Nanopart. Res. 2011, 13, 2759–2767. [Google Scholar] [CrossRef]
- Prader, A.; Raymond, A.P. Adenine nncleotide ratios and adenylate energy charge in energy metabolism. Ann. Rev. Plant Physiol. 1983, 34, 199–224. [Google Scholar] [CrossRef]
- Sleber, M.; Braendle, R. Energy metabolism in rhizomes of Acorus calamus (L.) and in tubers of Solanum tuberosum (L.) with regard to their anoxia tolerance. Bot. Acta 1991, 104, 279–282. [Google Scholar]
- Eberhard, K.S.; Falk, D.; Rolf, A. Effects of hydrogen sulfide to vibrio fischeri, scenedes musvacuolatus, and daphnia magna. Environ. Toxicol. Chem. 2005, 24, 2621–2629. [Google Scholar]
- Julian, D.; Dalia, W.E.; Arp, A.J. Neuromuscular sensitivity to hydrogen sulfide in the marine invertebrate Urechis caupo. J. Exp. Biol. 1998, 201, 1393–1403. [Google Scholar] [PubMed]
- Arbuckle, W.B.; Alleman, J.E. Effluent toxicity testing using nitrifiers and MicrotoxTM. Water Environ. Res. 1992, 64, 263–267. [Google Scholar] [CrossRef]
- Guidotti, T.L. Hydrogen sulphide. Occup. Med. (Lond) 1999, 646, 367–371. [Google Scholar] [CrossRef]
- Toussaint, M.W.; Shedd, T.R.; Vander Schaile, W.H.; Leather, G.R. A comparison of standard acute toxicity tests with rapid-screening toxicity tests. Environ. Toxicol. Chem. 1995, 14, 907–915. [Google Scholar] [CrossRef]
- Ma, M.; Tong, Z.; Wang, Z.; Zhu, W. Acute toxicity bioassay using the freshwater luminescent bacterium Vibrio qinghaiensis sp. Nov.-Q67. Bull. Environ. Contam. Toxicol. 1999, 62, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.L.; Shen, C.F.; Lu, Y.; Tang, X.J.; Zhang, C.K.; Chen, X.C.; Shi, J.Y.; Lin, Q.; Chen, Y.X. Hormesis response of marine andfreshwater luminescent bacteria to metal exposure. Biol. Res. 2009, 42, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, S.S.; Liu, H.L. Effect of ionic liquid on the toxicity of pesticide to Vibrio-qinghaiensis sp.-Q67. J. Hazar. Mater. 2009, 170, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.M.; Zhang, C.Y.; Ge, Z.G.; Wang, Z.Y. Toxicity measurement of the fluorobenzene derivants against Vibrio Qinghaiensis (Q67) and their 2D, 3D-QSAR study. Chin. J. Struct. Chem. 2010, 29, 1467–1476. [Google Scholar]
- Zhou, X.F.; Sang, W.J.; Liu, S.S.; Zhang, Y.L.; Ge, H.L. Modeling and prediction for the acute toxicity of pesticide mixtures to the freshwater luminescent bacterium Vibrio qinghaiensis sp.-Q67. J. Environ. Sci. 2010, 22, 433–440. [Google Scholar] [CrossRef]
- Salizzato, M.; Bertato, V.; Pavoni, B.; Volpi Ghirardini, A.; Ghetti, P.F. Sensitivity limits and EC50 values of the Vibrio fischeri test for organic micropollutants in natural and spiked extracts from sediments. Environ. Toxicol. Chem. 1998, 17, 655–661. [Google Scholar] [CrossRef]
- Johnson, T.; Laise, C.; Férard, J.F. Small-scale Freshwater Toxicity Investigations—Microtox® Acute Toxicity Test; Springer: Dordrecht, The Netherlands, 2005; pp. 69–105. [Google Scholar]
- Hargrave, B.T.; Holmer, M.; Newcombe, C.P. Mechanism for the hydrogen sulfide-induced growth limitation in wetland macrophytes. Mar. Pollut. Bull. 2008, 56, 810–824. [Google Scholar] [CrossRef] [PubMed]
- Lavilla, I.; Pena-Pereira, F.; Gil, S.; Costas, M.; Bendicho, C. Microvolume turbidimetry for rapid and sensitive determination of the acid labile sulfide fraction in waters after headspace single-drop microextraction with in situ generation of volatile hydrogen sulfide. Anal. Chim. Acta 2009, 647, 112–116. [Google Scholar] [CrossRef] [PubMed]
- van den Bosch, P.L.F.; van Beusekom, O.C.; Buisman, C.J.N.; Janssen, A.J.H. Sulfide oxidation at halo-alkaline conditions in a fed-batch bioreactor. Biotechnol. Bioeng. 2007, 97, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Padan, E.; Schuldiner, S. Intracellular pH regulation in bacterial cells. Meth. Enzymol. 1986, 125, 337–352. [Google Scholar] [PubMed]
- Krulwich, T.H.; Ito, M.; Gilmore, R.; Guffanti, A.A. Mechanisms of cytoplasmic pH regulation in alkaliphilic strains of Bacillus. Extremophiles 1997, 1, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Johnson, F.H.; Eyring, H.; Steblay, R.; Chaplin, H.; Huber, C.; Gherardi, G. The nature and control of reactions in bioluminescence: With special reference to the mechanism of reversible and irreversible inhibitions by hydrogen and hydroxyl ions, temperature, pressure, pressure, alcohol, urethane, and sulfanilamide in bacteria. J. Gen. Physiol. 1945, 28, 463–453. [Google Scholar] [CrossRef] [PubMed]
- Kudryasheva, N.S. Mechanisms of the effect of xenobiotics on bacterial bioluminescence. Luminescence 1999, 14, 199–200. [Google Scholar] [CrossRef]
- Bose, J.L.; Kim, U.; Bartkowski, W.; Gunsalus, R.P.; Overley, A.M.; Lyell, N.L.; Visick, K.L.; Stabb, E.V. Bioluminescence in Vibrio fischeri is controlled by the redox-responsive regulator ArcA. Mol. Microbiol. 2007, 65, 538–553. [Google Scholar] [CrossRef] [PubMed]
- Samhan-Arias, A.K.; Garcia-Bereguiain, M.A.; Gutierrez-Merino, C. Hydrogen sulfide is a reversible inhibitor of the NADH oxidase activity of synaptic plasma membranes. Biochem. Biophys. Res. Commun. 2009, 388, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Nykamp, J.; Huang, X.D.; Gerhardt, K.; Dixon, D.G.; Greenberg, B.M. Examination of the mechanism of phenanthrenequinone toxicity to Vibrio fischeri: Evidence for a reactive oxygen species-mediated toxicity mechanism. Environ. Toxicol. Chem. 2009, 28, 1655–1662. [Google Scholar] [CrossRef] [PubMed]
- Yakovlev, A.Y.; Tsodikov, A.D.; Bass, L.A. Stochastic model of hormesis. Math. Biosci. 1993, 116, 197–219. [Google Scholar] [CrossRef]
- Bounias, M.; Navonenectoux, M.; Popeskovic, D.S. Toxicology of cupric salts in honeybees. I. Hormesis effects of organic derivatives on lethality parameters. Ecotoxicol. Environ. Saf. 1995, 31, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Bounias, M.; Kruk, I.; Nectoux, M.; Popeskovic, D. Toxicology of cupric salts on honeybees. 5. Gluconate and sulfate action on gut alkaline and acid phosphatases. Ecotoxicol. Environ. Saf. 1996, 35, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.F.; Duan, W.X.; Xu, L.; Song, S.Z.; Zhu, C.C.; Wu, L. Biphasic effect of cadmium on cell proliferation in human embryo lung fibroblast cells and its molecular mechanism. Toxicol. In Vitro 2009, 23, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.W.; Shen, J.W.; Tomiyama-Miyaji, C.; Watanabe, M.; Kainuma, E.; Inoue, M.; Kuwano, Y.; Abo, T. Augmentation of innate immunity by low-dose irradiation. Cell. Immunol. 2006, 244, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.; Howe, K. Stimulation of growth of peppermint (Mentha piperita) by Phosfon, a growth retardant. Physiol. Plant. 1976, 37, 163–165. [Google Scholar] [CrossRef]
- Calabrese, E.J. Evidence that hormesis represents an ‘‘overcompensation’’ response to a disruption in homeostasis. Ecotoxicol. Environ. Saf. 1999, 42, 135–137. [Google Scholar] [CrossRef]
- Stebbing, A.R.D. A theory for growth hormesis. Belle Newsl. 1997, 9, 6. [Google Scholar] [CrossRef]
- Earson, J.; Havill, D.C. The effect of hypoxia and sulphide on culture-grown wetland and non-wetland plants II. Metabolic and physiological changes. Exp. Bot. 1988, 39, 431–439. [Google Scholar] [CrossRef]
- Calabrese, E.; Baldwin, L. Reevaluation of the fundamental dose response relationship. Bioscience 1999, 49, 725–731. [Google Scholar] [CrossRef]
- Fiorucci, S.; Distrutti, E.; Cirino, G.; Wallace, J.L. The emerging roles of hydrogen sulfide in the gastrointestinal tract and liver. Antioxid. Redox Signal. 2006, 5, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C. Hydrogen sulphide and its therapeutic potential. Nat. Rev. Drug Discov. 2007, 6, 917–935. [Google Scholar] [CrossRef] [PubMed]
- Calvert, J.W.; Coetzee, W.A.; Lefer, D.J. Novel insights into hydrogen sulfide-mediated cytoprotection. Antioxid. Redox Signal. 2010, 12, 1203–1217. [Google Scholar] [CrossRef] [PubMed]
- Figura, M.; Chilton, L.; Liacini, A.; Viskovic, M.M.; Phan, V.; Knight, D. Blockade of K(ATP) channels reduces endothelial hyperpolarization and leukocyte recruitment upon reperfusion after hypoxia. Am. J. Transplant. 2009, 9, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Mathai, J.C.; Missner, A.; Kügler, P.; Saparovb, S.M.; Zeidel, M.L.; Lee, J.K.; Pohl, P. No facilitator required for membrane transport of hydrogen sulfide. Proc. Natl. Acad. Sci. USA 2009, 106, 16633–16638. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.Q.; Ge, H.L.; Liu, S.S. Microplate luminometry for toxicity bioassay of environmental pollutant on a new type of fresh water luminescent bacterium (Vibrio qinghaiensis sp.-Q67). Asian J. Ecotoxicol. 2006, 1, 186–191. [Google Scholar]
- Ma, M.; Tong, Z.; Wang, Z.; Zhu, W. Acute toxicity bioassay using the freshwater luminescent bacterium Vibrio qinghaiensis sp. Nov.-Q67. Bull. Environ. Contam. Toxicol. 1999, 62, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Liu, S.S.; Song, X.Q.; Ge, H.L. Prediction for the mixture toxicity of six organophosphorus pesticides to the luminescent bacterium Q67. Ecotoxicol. Environ. Saf. 2008, 71, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Hargrave, B.T.; Holmer, M.; Newcombe, C.P. Mechanism for the hydrogen sulfide-induced growth limitation in wetland macrophytes. Mar. Pollut. Bull. 2008, 56, 810–824. [Google Scholar] [CrossRef] [PubMed]
- Lavilla, I.; Pena-Pereira, F.; Gil, S.; Costas, M.; Bendicho, C. Microvolume turbidimetry for rapid and sensitive determination of the acid labile sulfide fraction in waters after headspace single-drop microextraction with in situ generation of volatile hydrogen sulfide. Anal. Chim. Acta 2009, 647, 112–116. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |



{kind=link}
{kind=link}
{kind=link}
| pH | Total S, mg/L | H2S, mg/L | HS−, mg/L |
|---|---|---|---|
| 5.0 | 1.68 | 1.66 | 0.002 |
| 6.5 | 45.30 | 33.62 | 11.68 |
| 7.4 | 33.19 | 8.78 | 22.4 |
| 8.0 | 34.20 | 2.85 | 31.35 |
| 9.0 | 38.89 | 0.35 | 38.35 |
| 10.0 | 8.32 | 0.01 | 8.31 |
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shao, Y.; Wu, L.-L.; Gao, H.-W.; Wang, F. Effect of Soluble Sulfide on the Activity of Luminescent Bacteria. Molecules 2012, 17, 6046-6055. https://doi.org/10.3390/molecules17056046
Shao Y, Wu L-L, Gao H-W, Wang F. Effect of Soluble Sulfide on the Activity of Luminescent Bacteria. Molecules. 2012; 17(5):6046-6055. https://doi.org/10.3390/molecules17056046
Chicago/Turabian StyleShao, Ying, Ling-Ling Wu, Hong-Wen Gao, and Feng Wang. 2012. "Effect of Soluble Sulfide on the Activity of Luminescent Bacteria" Molecules 17, no. 5: 6046-6055. https://doi.org/10.3390/molecules17056046
APA StyleShao, Y., Wu, L. -L., Gao, H. -W., & Wang, F. (2012). Effect of Soluble Sulfide on the Activity of Luminescent Bacteria. Molecules, 17(5), 6046-6055. https://doi.org/10.3390/molecules17056046