Aqueous Fraction of Nephelium ramboutan-ake Rind Induces Mitochondrial-Mediated Apoptosis in HT-29 Human Colorectal Adenocarcinoma Cells

Abstract
:1. Introduction
2. Results and Discussion
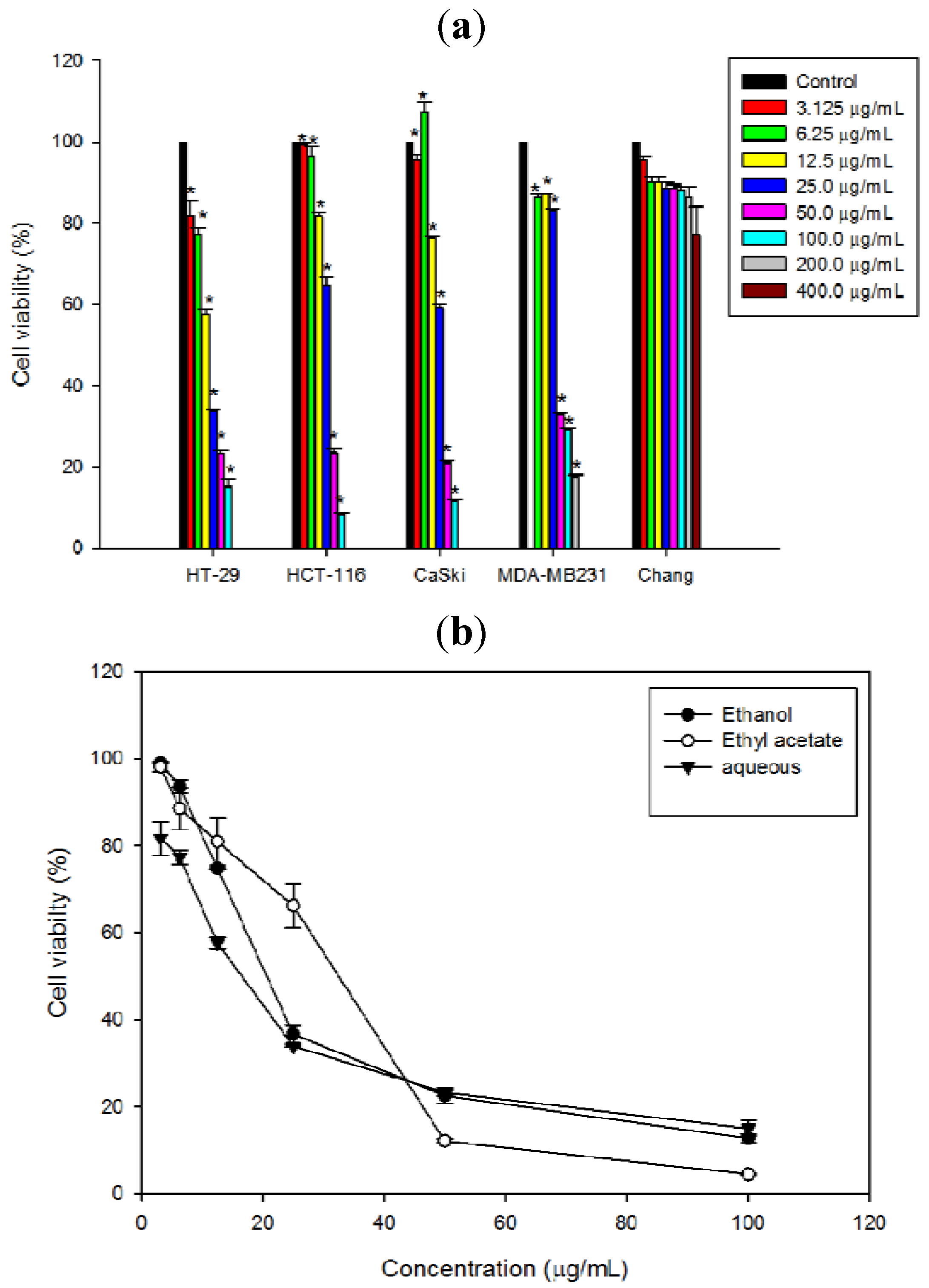
2.1. Reduction of HT-29 Cell Viability by NRAF
2.2. Preliminary Phytochemical Screening


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell lines | IC50 (µg/mL) | ||||
|---|---|---|---|---|---|
| Ethanol | Ethyl acetate | Aqueous | Curcumin * | ||
| HT-29 | 20.70 ± 0.49 | 32.24 ± 1.81 | 16.67 ± 0.55 | 21.32 ± 0.17 | |
| HCT-116 | 35.73 ± 0.56 | 47.23 ± 2.84 | 33.90 ± 1.06 | NA | |
| Ca Ski | 34.38 ± 0.66 | 44.90 ± 0.58 | 31.14 ± 0.41 | NA | |
| MDA-MB-231 | 51.09 ± 1.32 | 61.65 ± 0.42 | 41.53 ± 0.32 | NA | |
| Phytochemical test | Results * |
|---|---|
| Flavonoids | ++ |
| Tannins | +++ |
| Saponins | +++ |
| Alkaloids | − |
| Sterols | − |
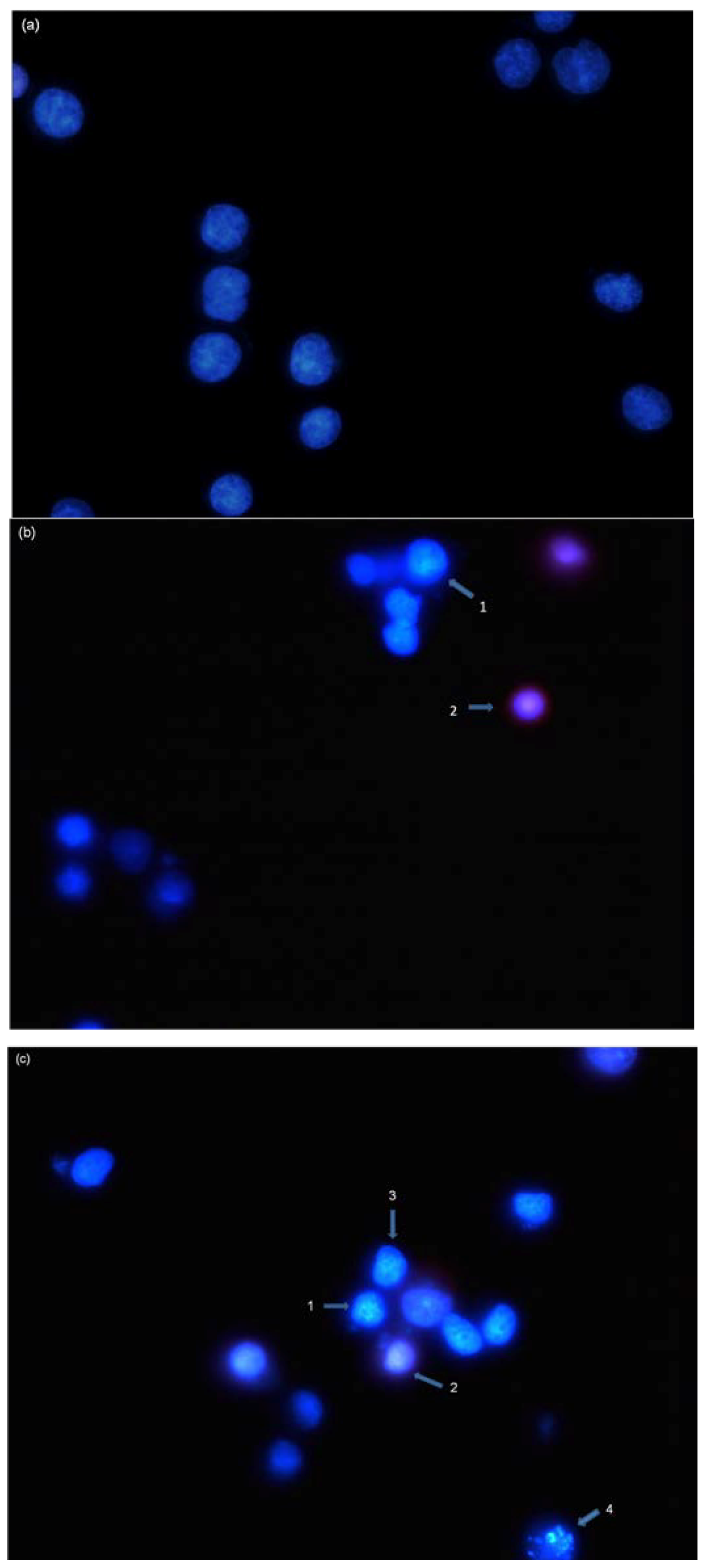
2.3. Induction of Nuclear Morphological Changes by NRAF

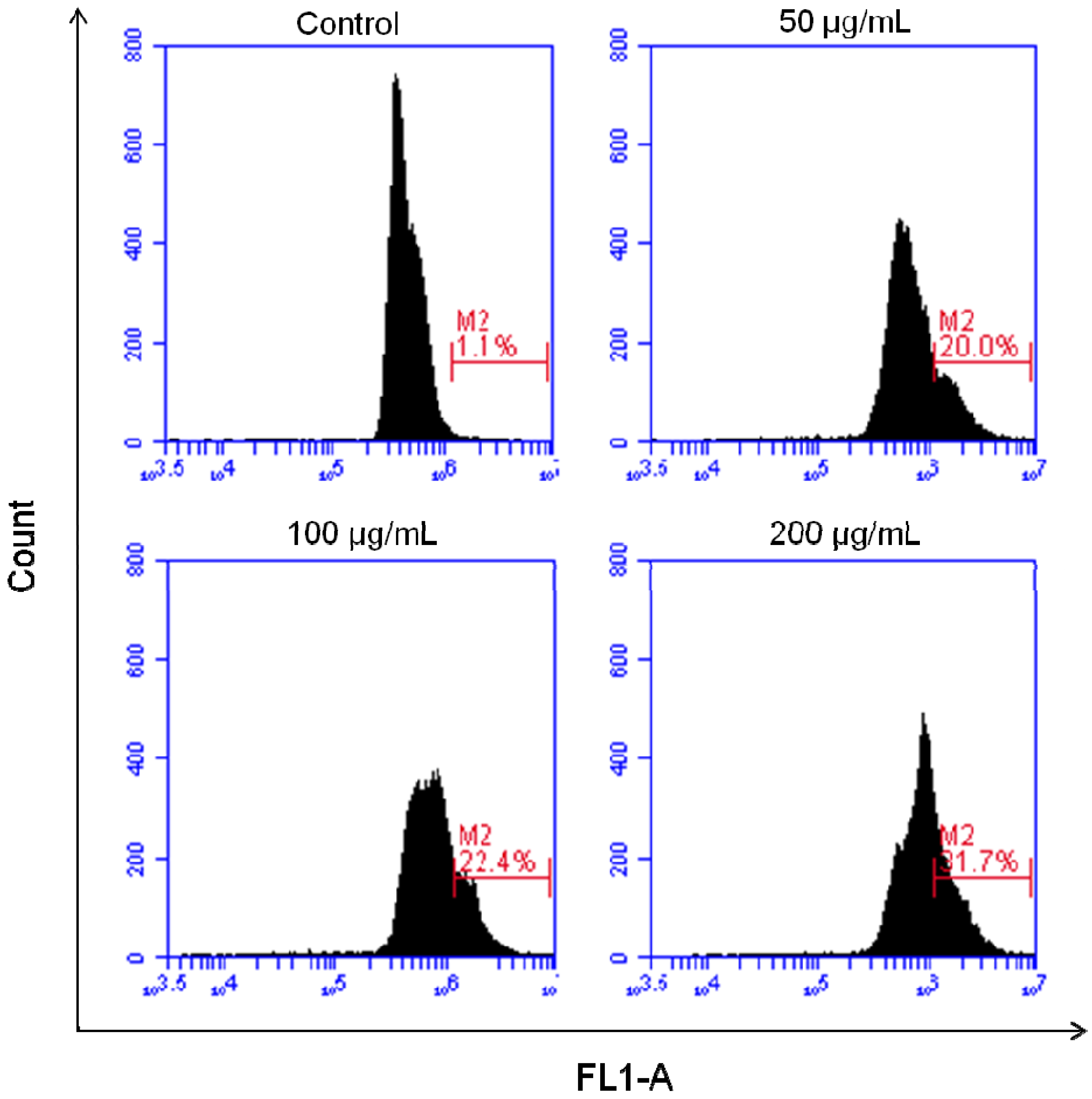
2.4. Induction of DNA Fragmentation Detected by TUNEL Assay

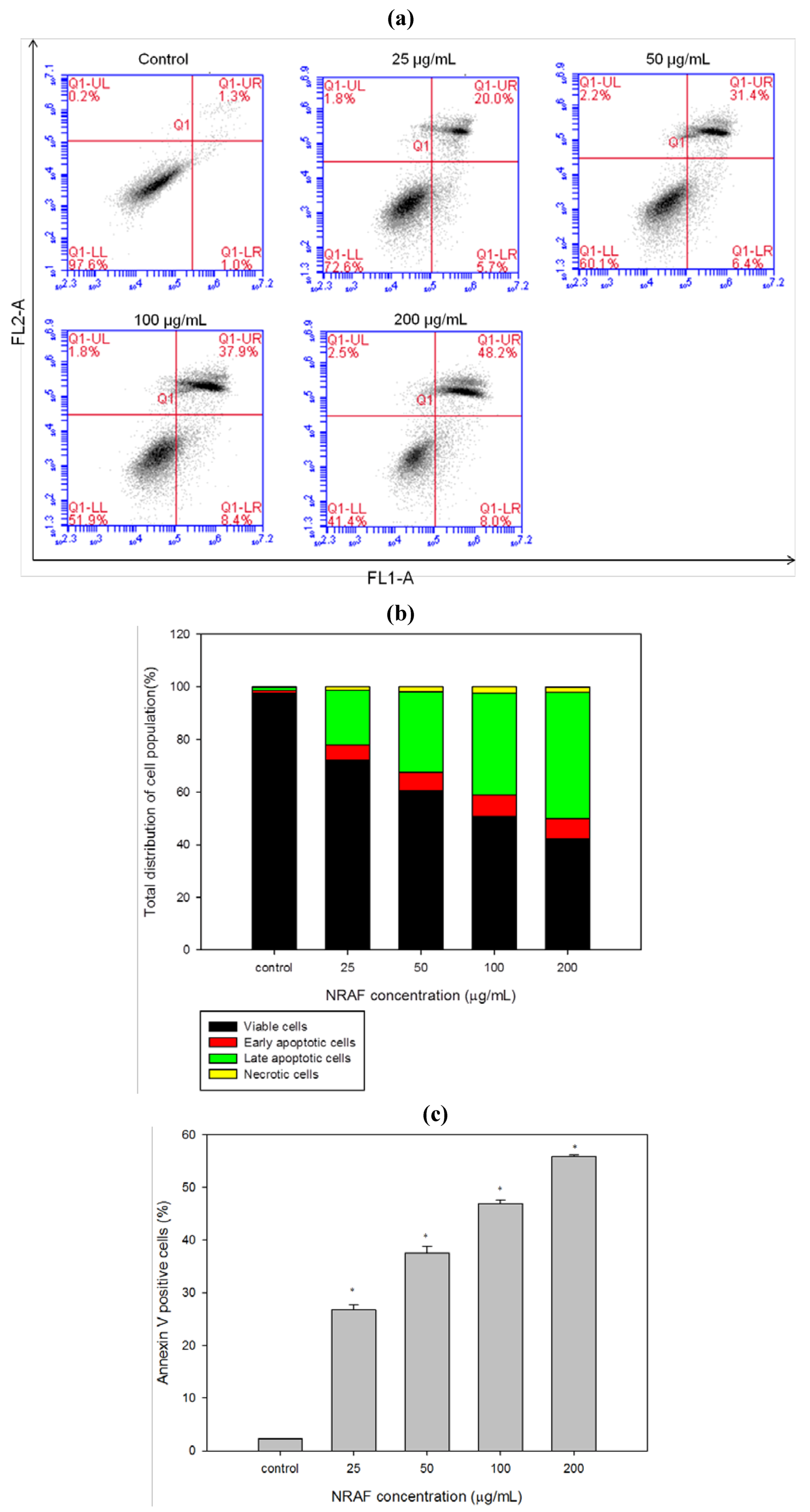
2.5. Induction of Early and Late Apoptosis Using Annexin V/PI Staining.

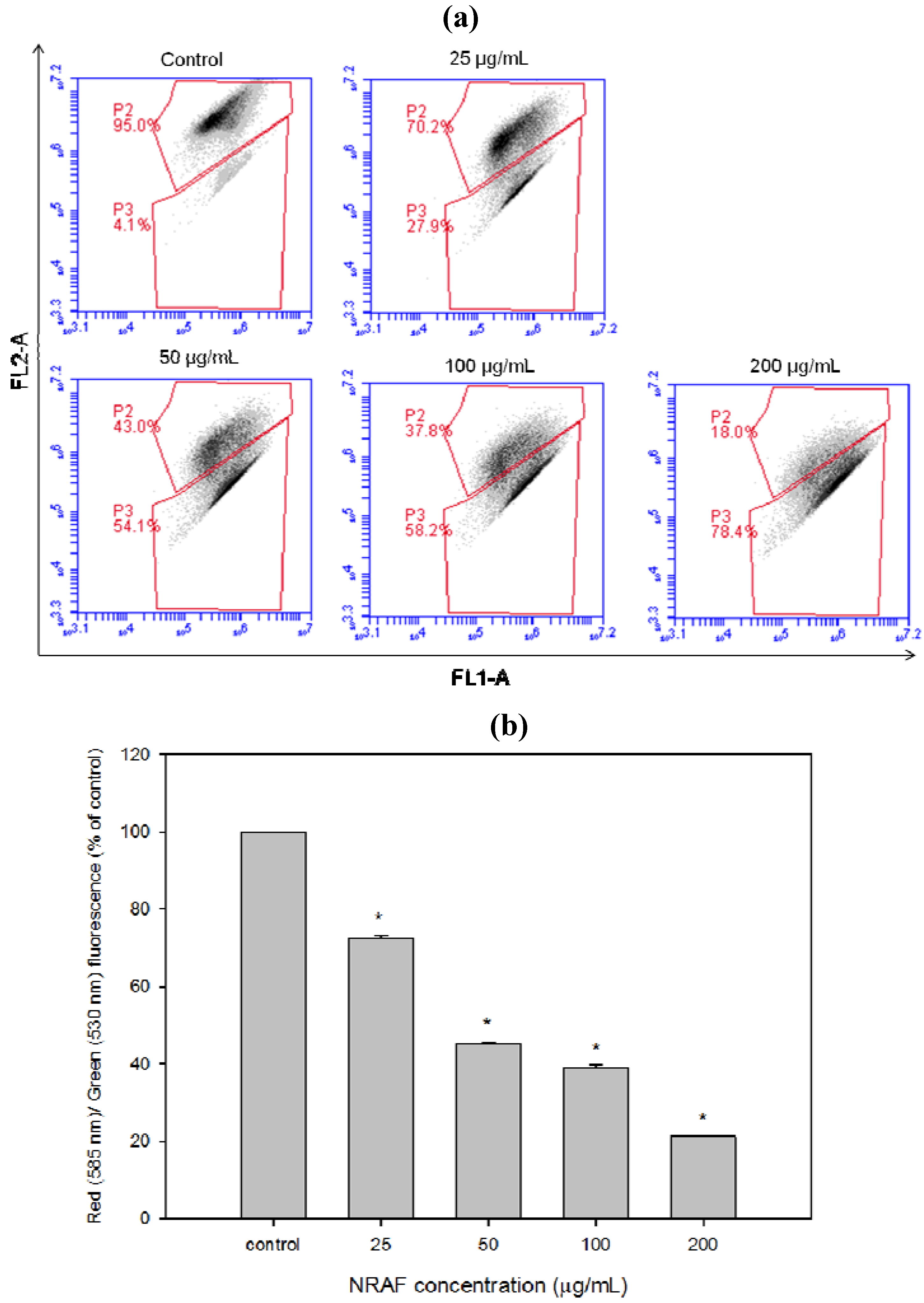
2.6. Dissipation of Mitochondrial Membrane Potential (Δψm) Triggered by NRAF in HT-29 Cells

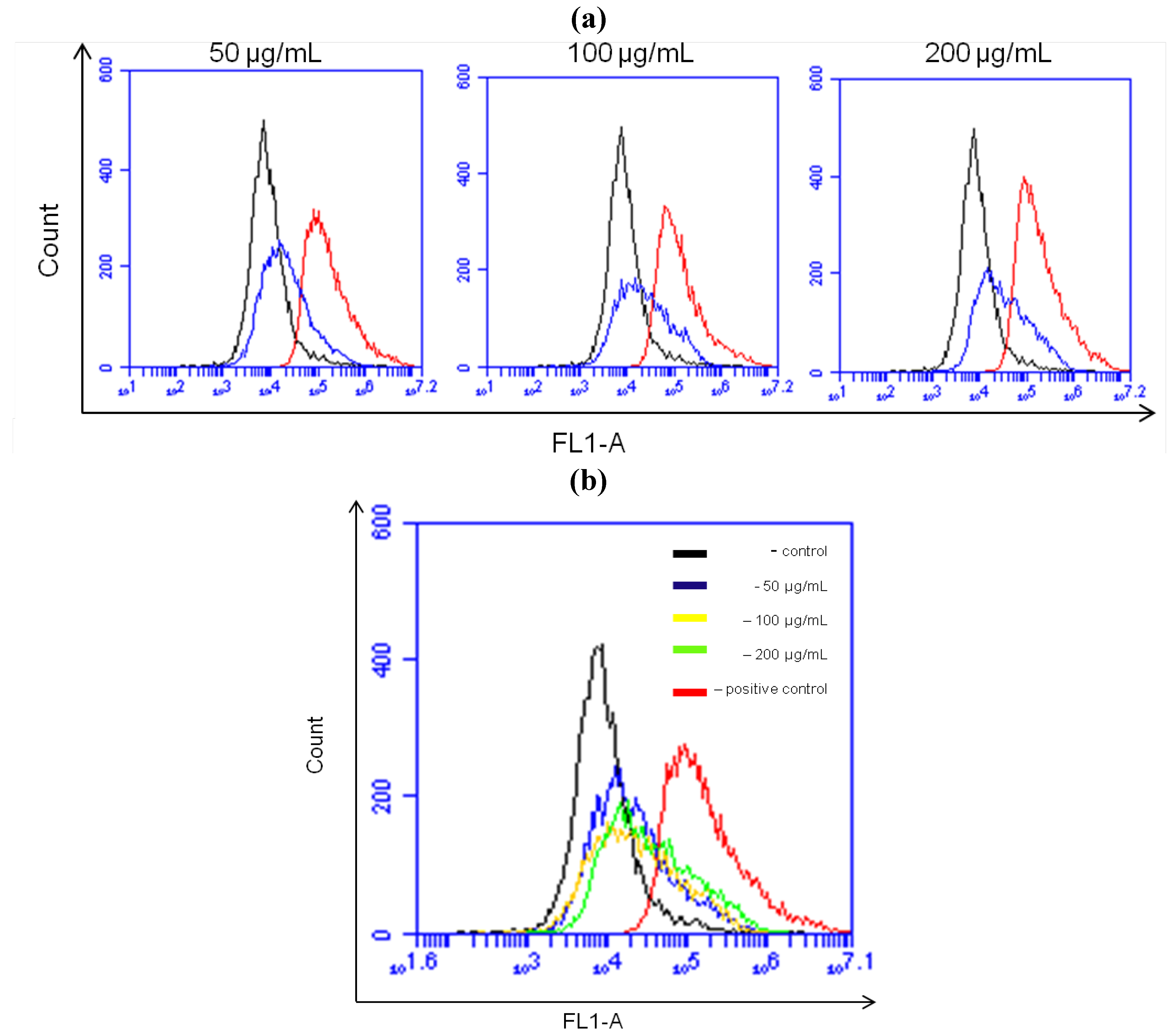
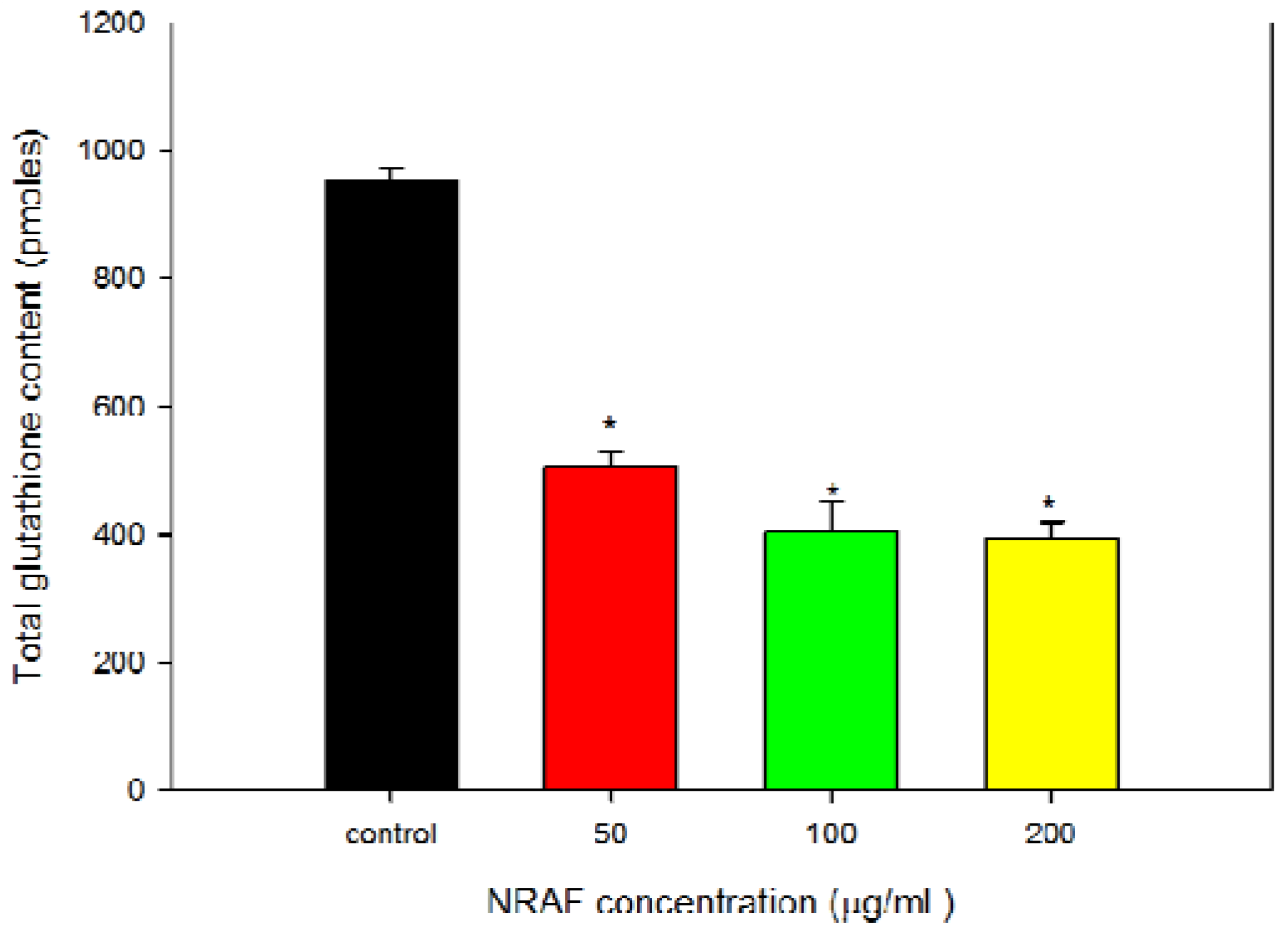
2.7. NRAF Effect on Intracellular ROS and GSH Levels in HT-29 Cells

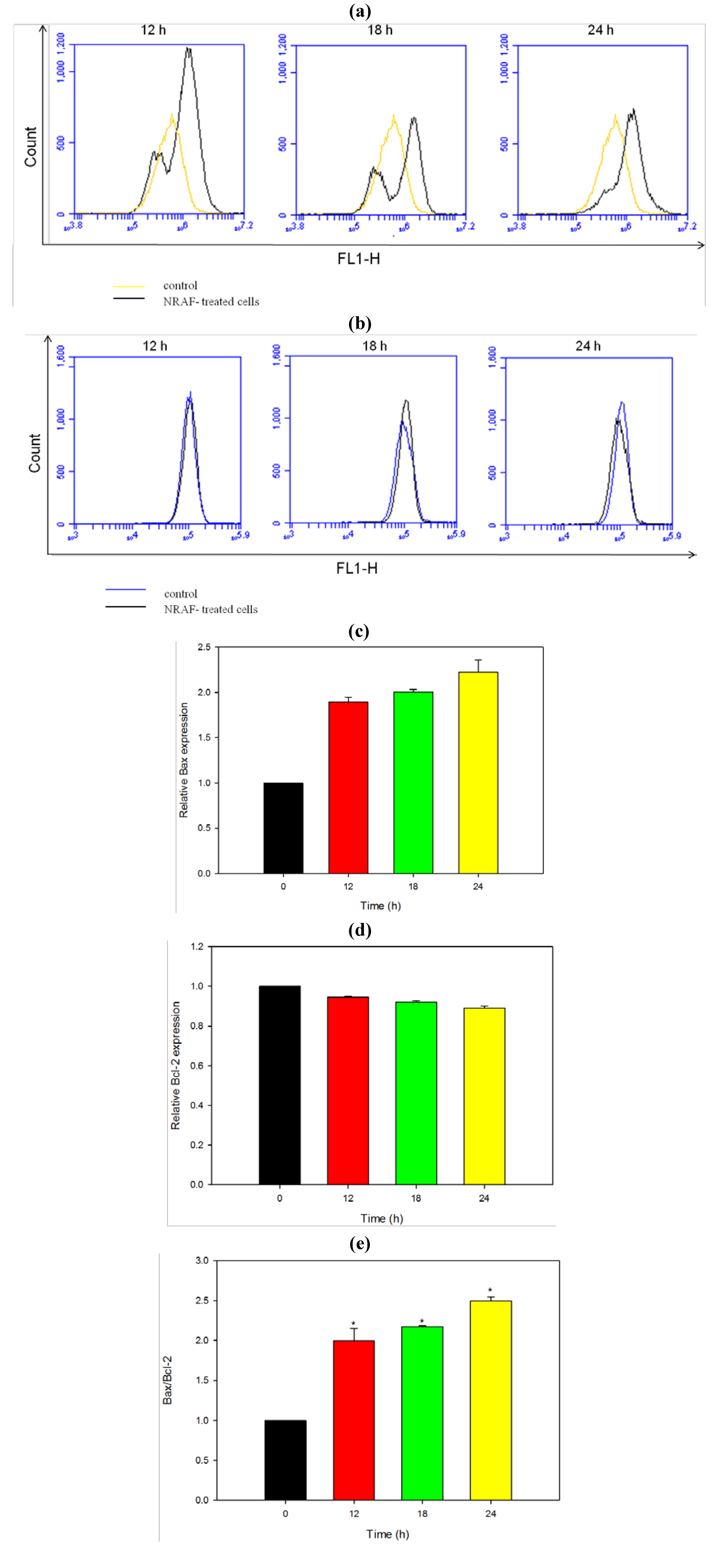
2.8. Modulation of Apoptotic Proteins by NRAF


2.9. NRAF Induced Caspase-3/7 and Caspase 9 Activation

3. Experimental
3.1. Plant Material
3.2. Preparation of Crude N. ramboutan-ake Extract and Fractions
3.3. Cell Culture
3.4. In Vitro MTT Cytotoxicity Assay
3.5. Preliminary Phytochemical Screening
3.5.1. Test for Alkaloids
3.5.2. Sakowski Test for Sterols
3.5.3. Frothing Test for Saponins
3.5.4. Test for Tannins
3.5.5. Test for Flavonoids
3.6. Nuclear Morphology Detection Using Hoechst 33342/PI
3.7. Terminal Deoxynucleotidyl Transferased UTP Nick End Labeling (TUNEL) Assay
3.8. Annexin V/PI Staining for the Assessment of Phosphatidylserine Externalization
3.9. Measurement of Mitochondrial Membrane Potential (Δψm)
3.10. Determination of Intracellular Total Glutathione (GSH) Content
3.11. Measurement of Intracellular Reactive Oxygen Species (ROS)
3.12. Determination of Bax and Bcl-2 Protein Expression Level
3.13. Measurement of Caspase-3/7 and Caspase-9 Activities
3.14. Statistical Analysis
4. Conclusions
Acknowledgments
- Sample Availability: Samples are available from the authors.
References and notes
- Cancer Facts & Figures 2012; American Cancer Society: Atlanta, GA, USA, 2012.
- Pan, M.H.; Chiou, Y.S.; Wang, Y.J.; Ho, C.T.; Lin, J.K. Multistage carcinogenesis process as molecular targets in cancer chemoprevention by epicatechin-3-gallate. Food Funct. 2011, 2, 101–110. [Google Scholar] [CrossRef]
- Fearon, E.R. Molecular genetics of colorectal cancer. Ann. Rev. Pathol. 2011, 6, 479–507. [Google Scholar]
- Grady, W.M.; Carethers, J.M. Genomic and epigenetic instability in colorectal cancer pathogenesis. Gastroenterology 2008, 135, 1079–1099. [Google Scholar] [CrossRef]
- Wyllie, A.H. Apoptosis: An overview. Br. Med. Bull. 1997, 53, 451–465. [Google Scholar] [CrossRef]
- Fleischer, A.; Ghadiri, A.; Dessauge, F.; Duhamel, M.; Rebollo, M.P.; Alvarez-Franco, F.; Rebollo, A. Modulating apoptosis as a target for effective therapy. Mol. Immunol. 2006, 43, 1065–1079. [Google Scholar] [CrossRef]
- Petit, P.X.; Lecoeur, H.; Zorn, E.; Dauguet, C.; Mignotte, B.; Gougeon, M.L. Alterations in mitochondrial structure and function are early events of dexamethasone-induced thymocyte apoptosis. J. Cell Biol. 1995, 130, 157–167. [Google Scholar] [CrossRef]
- Tan, S.; Sagara, Y.; Liu, Y.; Maher, P.; Schubert, D. The regulation of reactive oxygen species production during programmed cell death. J. Cell Biol. 1998, 141, 1423–1432. [Google Scholar] [CrossRef]
- Ka, H.; Park, H.; Jung, H.; Choi, J.; Cho, K.; Ha, J.; Lee, K. Cinnamaldehyde induces apoptosis by ROS-mediated mitochondrial permeability transition in human promyelocytic leukemia HL-60 cells. Cancer Lett. 2003, 196, 143–152. [Google Scholar] [CrossRef]
- Whitfield, J.F. Calcium, calcium-sensing receptor and colon cancer. Cancer Lett. 2009, 275, 9–16. [Google Scholar] [CrossRef]
- Hao, X.; Du, M.; Bishop, A.E.; Talbot, I.C. Imbalance between proliferation and apoptosis in the development of colorectal carcinoma. Virchows Archiv. 1998, 433, 523–527. [Google Scholar] [CrossRef]
- Xie, C.M.; Chan, W.Y.; Yu, S.; Zhao, J.; Cheng, C.H.K. Bufalin induces autophagy-mediated cell death in human colon cancer cells through reactive oxygen species generation and JNK activation. Free Radic. Biol. Med. 2011, 51, 1365–1375. [Google Scholar] [CrossRef]
- McCullough, M.L.; Patel, A.V.; Kushi, L.H.; Patel, R.; Willett, W.C.; Doyle, C.; Thun, M.J.; Gapstur, S.M. Following cancer prevention guidelines reduces risk of cancer, cardiovascular disease, and all-cause mortality. Cancer Epidemiol. Biomar. Prev. 2011, 20, 1089–1097. [Google Scholar] [CrossRef]
- Cooper, A.J.; Sharp, S.J.; Lentjes, M.A.; Luben, R.N.; Khaw, K.T.; Wareham, N.J.; Forouhi, N.G. A Prospective Study of the Association Between Quantity and Variety of Fruit and Vegetable Intake and Incident Type 2 Diabetes. Diabetes Care 2012, 35, 1293–1300. [Google Scholar] [CrossRef]
- Palanisamy, U.; Cheng, H.M.; Masilamani, T.; Subramaniam, T.; Ling, L.T.; Radhakrishna, A.K. Rind of rambutan, Nephelium lappaceum, a potential source of natural antioxidants. Food Chem. 2008, 109, 54–63. [Google Scholar]
- Seeram, N.P.; Adams, T.L.S.; Henning, S.M.; Niu, Y.; Zhang, Y.; Nair, M.G.; Heber, D. In vitro antiproliferative, apoptotic and antioxidant activities of punicalagin, ellagic acid and a total pomegranate tannin extract are enhanced in combination with other polyphenols as found in pomegranate juice. J. Nutr. Biochem. 2005, 16, 360–367. [Google Scholar] [CrossRef]
- Palanisamy, U.; Ling, L.T.; Manaharan, T.; Appleton, D. Rapid Isolation of geraniin from Nephelium lappaceum rind waste and its anti-hyperglycemic activity. Food Chem. 2011, 127, 21–27. [Google Scholar] [CrossRef]
- Moongkarndi, P.; Kosema, N.; Kaslungka, S.; Luanratana, O.; Pongpan, N.; Neungton, N. Antiproliferation, antioxidation and induction of apoptosis by Garcinia mangostana (mangosteen) on SKBR3 human breast cancer cell line. J. Ethnopharmacol. 2004, 90, 161–166. [Google Scholar] [CrossRef]
- Thitilertdecha, N.; Teerawutgulrag, A.; Kilburn, J.D.; Rakariyatham, N. Identification of major phenolic compounds from Nephelium lappaceum L. and their antioxidant activities. Molecules 2010, 15, 1453–1465. [Google Scholar] [CrossRef]
- Ito, A.; Chai, H.B.; Kardono, L.B.; Setowati, F.M.; Afriastini, J.J.; Riswan, S.; Farnsworth, N.R.; Cordell, G.A.; Pezzuto, J.M.; Swanson, S.M.; et al. Saponins from the bark of Nephelium maingayi. J. Nat. Prod. 2004, 67, 201–205. [Google Scholar] [CrossRef]
- Lee, J.C.; Tsai, C.Y.; Kao, J.Y.; Kao, M.C.; Tsai, S.C.; Chang, C.S.; Huang, L.J.; Kuo, S.C.; Lin, J.K.; Way, T.D. Geraniin-mediated apoptosis by cleavage of focal adhesion kinase through up-regulation of Fas ligand expression in human melanoma cells. Mol. Nutr. Food Res. 2008, 52, 655–663. [Google Scholar] [CrossRef]
- Martin, R.; Ibeas, E.; Carvalho-Tavares, J.; Hernandez, M.; Ruiz-Gutierrez, V.; Nieto, M.L. Natural triterpenic diols promote apoptosis in astrocytoma cells through ROS-mediated mitochondrial depolarization and JNK activation. PLoS One 2009, 4, e5975. [Google Scholar]
- Li, T.M.; Chen, G.W.; Su, C.C.; Lin, J.G.; Yeh, C.C.; Cheng, K.C.; Chung, J.G. Ellagic acid induced p53/p21 expression, G1 arrest and apoptosis in human bladder cancer T24 cells. Anticancer Res. 2005, 25, 971–979. [Google Scholar]
- Larrosa, M.; Tomas-Barberan, F.A.; Espin, J.C. The dietary hydrolysable tannin punicalagin releases ellagic acid that induces apoptosis in human colon adenocarcinoma Caco-2 cells by using the mitochondrial pathway. J. Nutr. Biochem. 2006, 17, 611–625. [Google Scholar] [CrossRef]
- Kerr, J.F.R.; Harmon, B.V. Definition and incidence of apoptosis: An historical perspective. In Apoptosis: The Molecular Basis of Programmed Cell Death; Tomei, L.D., Cope, F.O., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbour, NY, USA, 1991; pp. 5–29. [Google Scholar]
- Gavreli, Y.; Sherman, Y.; Ben-Sasson, S.A. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 1992, 119, 493–501. [Google Scholar]
- Compton, M.M. A biochemical hallmark of apoptosis: Internucleosomal degradation of the genome. Cancer Metastasis Rev. 1992, 11, 105–119. [Google Scholar] [CrossRef]
- Koopman, G.; Reutelingsperger, C.P.M.; Kuijten, G.A.M.; Keehnen, R.M.J.; Pals, S.T.; Van Oers, M.H.J. Annexin V for flow cytometric detection of phosphatidylserine expression on B cells undergoing apoptosis. Blood 1994, 84, 1415–1420. [Google Scholar]
- Fadok, V.A.; Voelker, D.R.; Campbell, P.A.; Cohen, J.J.; Bratton, D.L.; Henson, P.M. Exposure of phosphatidylserine on the surface of apoptotic lymphocytes triggers specific recognition and removal by macrophages. J. Immunol. 1992, 148, 2207–2216. [Google Scholar]
- Vermes, I.; Haanen, C.; Steffens-Nakken, H.; Reutelingsperger, C. A novel assay for apoptosis Flow cytometric detection of phosphatidylserine early apoptotic cells using fluorescein labeled expression on Annexin V. J. Immunol. Method. 1995, 184, 39–51. [Google Scholar] [CrossRef]
- Callahan, M.K.; Williamson, P.; Schlegel, R.A. Surface expression of Phosphatidylserine on macrophages is required for phagocytosis of apoptotic thymocytes. Cell Death Differ. 2000, 7, 645–653. [Google Scholar] [CrossRef]
- Chiu, H.C.; Chih, T.T.; Hsian, Y.M.; Tseng, C.H.; Wu, M.J.; Wu, Y.C. Bullatacin, a potent antitumor annonaceous acetogenin, induced apoptosis through a reduction of intracellular cAMP and cGMP levels in human hepatoma 2.2.15 cells. Biochem. Pharmacol. 2003, 65, 319–327. [Google Scholar]
- Kroemer, G.; Galluzzi, L.; Brenner, C. Mitochondrial membrane permeabilization in cell death. Physiol. Rev. 2007, 87, 99–163. [Google Scholar] [CrossRef]
- Han, J.; Goldstein, L.A.; Gastman, B.R.; Rabinowich, H. Interrelated roles for Mcl-1 and BIM in regulation of TRAIL-mediated mitochondrial apoptosis. J. Biol. Chem. 2006, 281, 10153–10163. [Google Scholar] [CrossRef]
- Fu, Y.R.; Yi, Z.J.; Yan, Y.R.; Qiu, Z.Y. Hydroxycamptothecin-induced apoptosis in hepatoma SMMC-7721 cells and the role of mitochondrial pathway. Mitochondrion 2006, 6, 211–217. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef]
- Pramanik, K.C.; Boreddy, S.R.; Srivastava, S.K. Role of mitochondrial electron transport chain complexes in capsaicin mediated oxidative stress leading to apoptosis in pancreatic cancer cells. PLoS One 2011, 6, e20151. [Google Scholar]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Lluis, J.M.; Buricchi, F.; Chiarugi, P.; Morales, A.; Fernandez-Checa, J.C. Dual role of mitochondrial reactive oxygen species in hypoxia signaling: Activation of nuclear factor-kB via c-SRC and oxidant-dependent cell death. Cancer Res. 2007, 67, 7368–7377. [Google Scholar] [CrossRef]
- Chen, W.Q.; Shen, W.; Shen, D.M. The changes of ROS and mitochondria membrane potential in HepG2 cells on the pressure of cisplatin. Zhonghua Gan Zang Bing Za Zhi 2005, 13, 531–533. [Google Scholar]
- Sandur, S.K.; Ichikawa, H.; Pandey, M.K.; Kunnumakkara, A.B.; Sung, B.; Sethi, G.; Aggarwal, B.B. Role of pro-oxidants and antioxidants in the anti-inflammatory and apoptotic effects of curcumin (diferuloylmethane). Free Radic. Biol. Med. 2007, 43, 568–580. [Google Scholar] [CrossRef]
- Desagher, S.; Osen-Sand, A.; Nichols, A.; Eskes, R.; Montessuit, S.; Lauper, S.; Maundrell, K.; Antonsson, B.; Mrtinou, J. Bid-induced conformational change of Bax is responsible for mitochondrial cytochrome c release during apoptosis. J. Cell Biol. 1999, 144, 891–901. [Google Scholar] [CrossRef]
- Huang, S.T.; Yang, R.C.; Yang, L.J.; Lee, P.N.; Pang, J.H. Phyllanthus urinaria triggers the apoptosis and Bcl-2 down-regulation in Lewis lung carcinoma cells. Life Sci. 2003, 72, 1705–1716. [Google Scholar] [CrossRef]
- Reed, J.; Miyashita, T.; Takayama, S.; Wang, H.G.; Sato, T.; Krajewski, S.; Aime-Sempe, C.; Bodrug, S.; Kitada, S.; Hanada, M. Bcl-2 family proteins: regulators of cell death involved in the pathogenesis of cancer and resistance to therapy. J. Cell Biochem. 1996, 60, 23–32. [Google Scholar] [CrossRef]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef]
- Sharpe, J.C.; Arnoult, D.; Youle, R.J. Control of mitochondrial permeability by Bcl-2 family members. Biochim. Biophys. Acta 1644, 107–113. [Google Scholar]
- Nomura, M.; Shimizu, S.; Ito, T.; Narita, M.; Matsuda, H.; Tsujimoto, Y. Apoptotic cytosol facilitates Bax translocation to mitochondria that involves cytosolic factor regulated by Bcl-2. Cancer Res. 1999, 59, 5542–5548. [Google Scholar]
- Murphy, K.M.; Ranganathan, V.; Farnsworth, M.L.; Kavallaris, M.; Lock, R.B. Bcl-2 inhibits Bax translocation from cytosol to mitochondria during drug-induced apoptosis of human tumor cells. Cell Death Differ. 2000, 7, 102–111. [Google Scholar] [CrossRef]
- Ju, H.K.; Lee, H.W.; Chung, K.S.; Choi, J.H.; Cho, J.G.; Baek, N.I.; Chung, H.G.; Lee, K.T. Standardized flavonoid-rich fraction of Artemisia princeps Pampanini cv. Sajabal induces apoptosis via mitochondrial pathway in human cervical cancer HeLa cells. J. Ethnopharmacol. 2012, 141, 460–468. [Google Scholar] [CrossRef]
- Jin, S.; Zhang, Q.Y.; Kang, X.M.; Wang, J.X.; Zhao, W.H. Daidzein induces MCF-7 breast cancer cell apoptosis via the mitochondrial pathway. Ann. Oncol. 2010, 21, 263–268. [Google Scholar] [CrossRef]
- Xiang, J.; Chao, D.T.; Korsmeyer, S.J. BAX-induced death may not require interleukin 1β-converting enzyme-like proteases. Proc. Natl. Acad. Sci. USA 1996, 93, 14559–14563. [Google Scholar] [CrossRef]
- Danial, N.N.; Korsmeyer, S.J. Cell death: Critical control points. Cell 2004, 116, 205–219. [Google Scholar] [CrossRef]
- Ghavami, S.; Hashemi, M.; Ande, S.R.; Yeganeh, B.; Xiao, W.; Eshraghi, M.; Bus, C.J.; Kadkhoda, K.; Wiechec, E.; Halayko, A.J.; et al. Apoptosis and cancer: mutations within caspase genes. J. Med. Genet. 2009, 46, 497–510. [Google Scholar] [CrossRef]
- Yang, J.; Xuesong, L. Prevention of apoptosis by Bcl-2: Release of cytochrome C from mitochondria blocked. Science 1997, 275, 1129–1132. [Google Scholar] [CrossRef]
- Sofowora, A. Medicinal Plants and Traditional Medicine in Africa, 2nd ed; Spectrum Books Limited: Ibadan, Nigeria, 1993. [Google Scholar]
- Trease, G.E.; Evans, W.C. Pharmacognosy, 13th ed; Bailliere Tindall: London, UK, 1989; pp. 683–684. [Google Scholar]
- Bibi, Y.N.S.; Waheed, A.; Zia, M.; Sarwar, S.; Ahmed, S.; Chaudhary, M.F. Evaluation of Viburnum foetens for anticancer and antibacterial potential and phytochemical analysis. Afr. J. Biotechnol. 2010, 9, 5611–5615. [Google Scholar]
- Roussi, S.; Gosse, F.; Aoude-Werner, D.; Zhang, X.; Marchioni, E.; Geoffroy, P.; Miesch, M.; Raul, F. Mitochondrial perturbation, oxidative stress and lysosomal destabilization are involved in 7beta-hydroxysitosterol and 7beta hydroxycholesterol triggered apoptosis in human colon cancer cells. Apoptosis 2007, 12, 87–96. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chan, C.K.; Goh, B.H.; Kamarudin, M.N.A.; Kadir, H.A. Aqueous Fraction of Nephelium ramboutan-ake Rind Induces Mitochondrial-Mediated Apoptosis in HT-29 Human Colorectal Adenocarcinoma Cells. Molecules 2012, 17, 6633-6657. https://doi.org/10.3390/molecules17066633
Chan CK, Goh BH, Kamarudin MNA, Kadir HA. Aqueous Fraction of Nephelium ramboutan-ake Rind Induces Mitochondrial-Mediated Apoptosis in HT-29 Human Colorectal Adenocarcinoma Cells. Molecules. 2012; 17(6):6633-6657. https://doi.org/10.3390/molecules17066633
Chicago/Turabian StyleChan, Chim Kei, Bey Hing Goh, Muhamad Noor Alfarizal Kamarudin, and Habsah Abdul Kadir. 2012. "Aqueous Fraction of Nephelium ramboutan-ake Rind Induces Mitochondrial-Mediated Apoptosis in HT-29 Human Colorectal Adenocarcinoma Cells" Molecules 17, no. 6: 6633-6657. https://doi.org/10.3390/molecules17066633