Liquiritigenin Inhibits Tumor Growth and Vascularization in a Mouse Model of Hela Cells

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
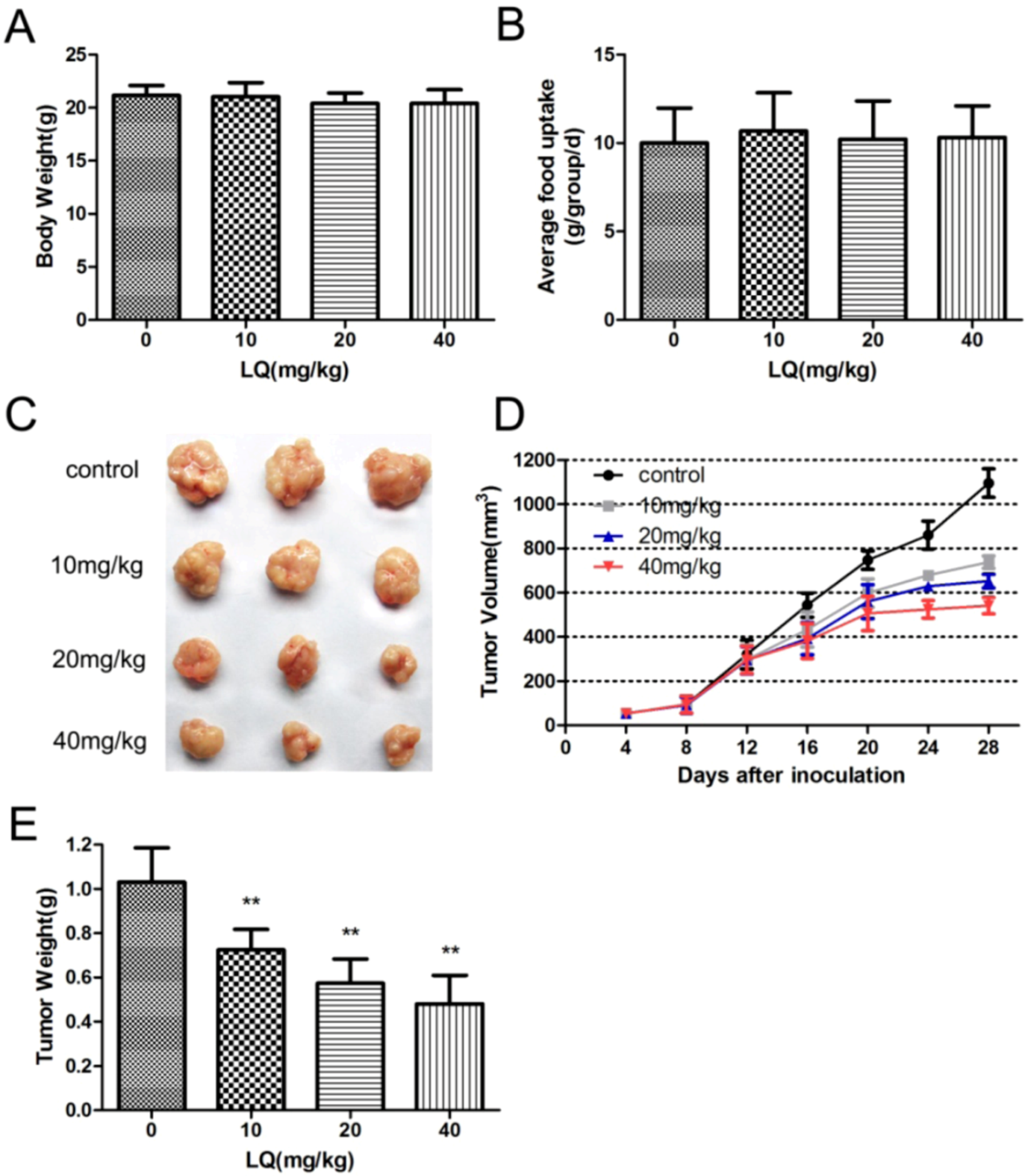
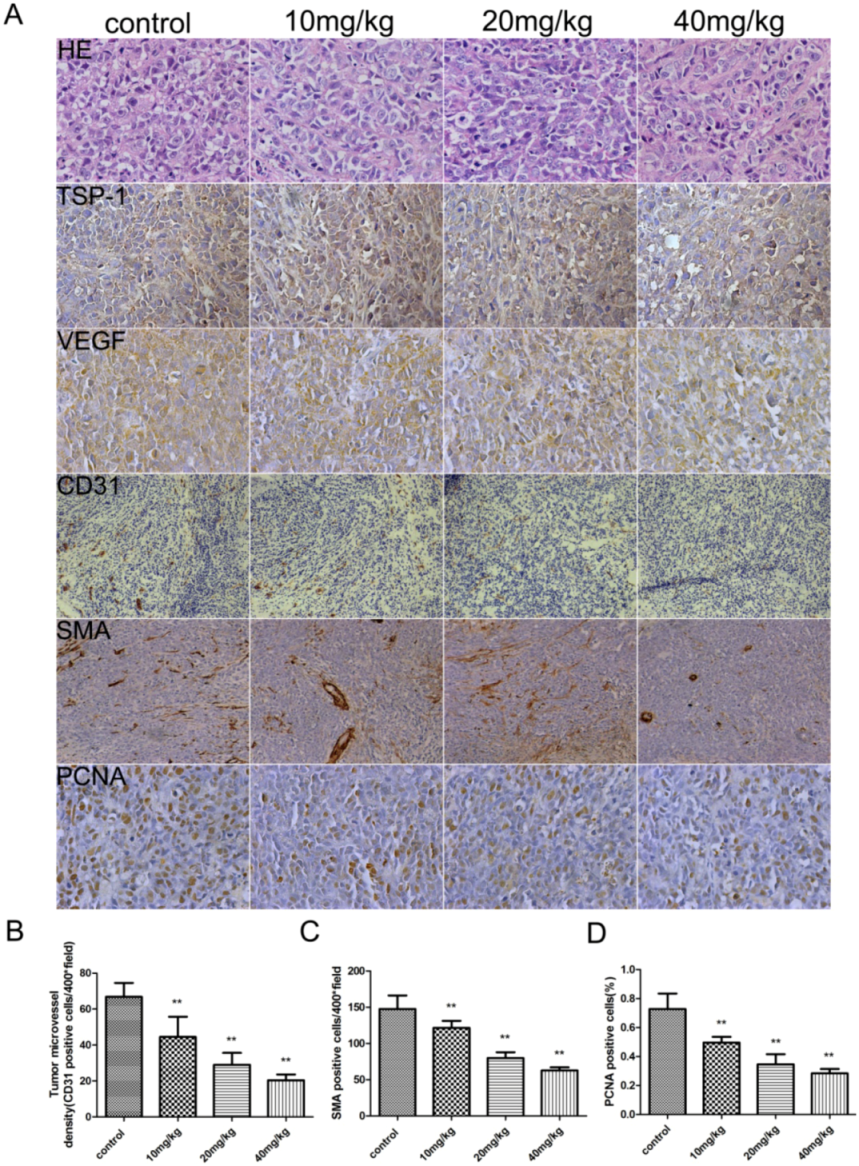
2.1. Liquiritigenin Attenuate the Rate of Tumor Growth in Vivo


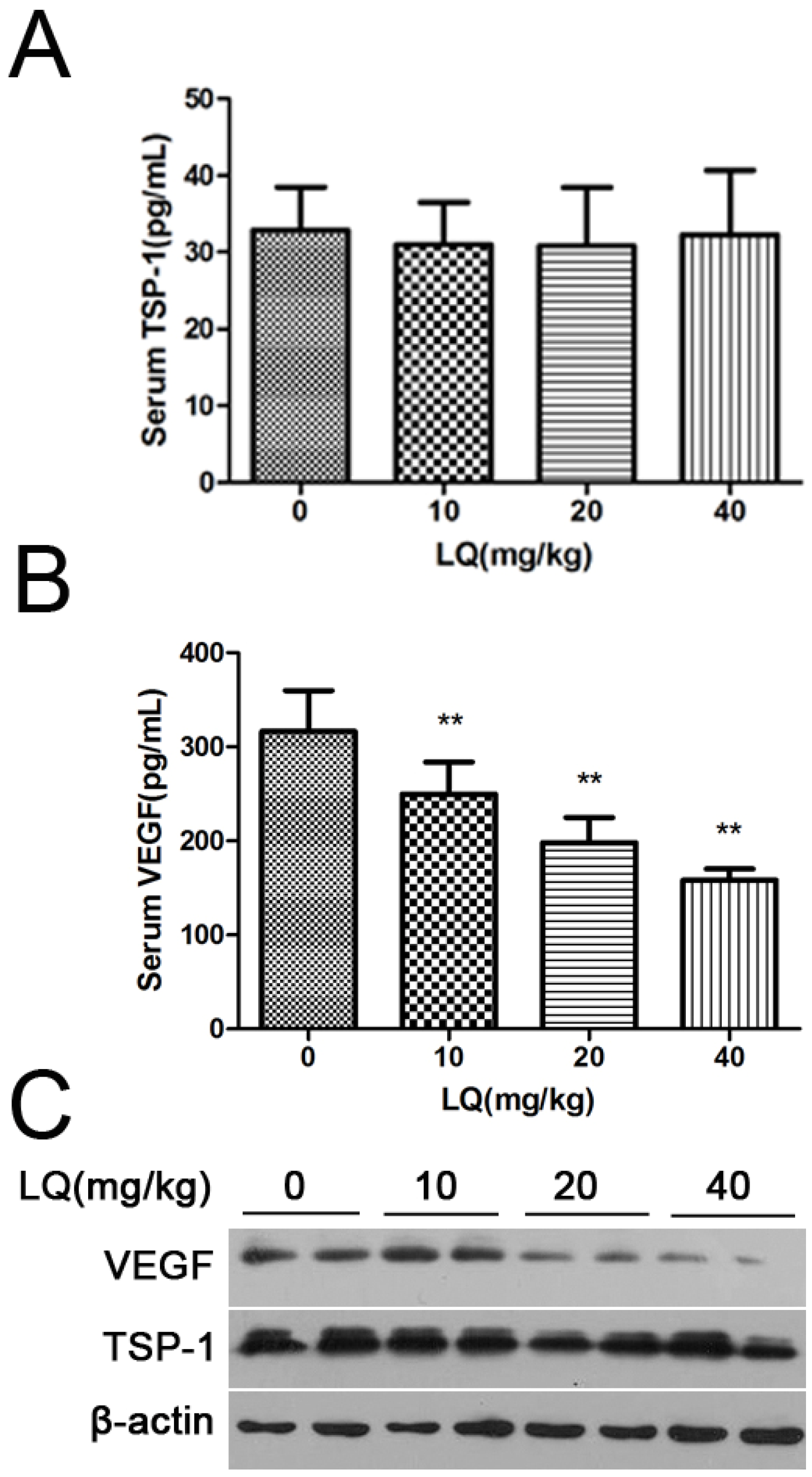
2.2. Liquiritigenin Reduced the Expression of VEGF in HeLa Cells

2.3. Discussion
3. Experimental
3.1. Chemicals and Drugs
3.2. Cell Culture
3.3. Mice and Tumor Xenografts
3.4. Histopathological Evaluation
3.5. Immunohistochemistry
3.6. Elisa Measurements
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Acknowledgements
Conflict of Interests
- Sample Availability: Samples of the compounds are not available from the authors.
References and Notes
- Folkman, J. Role of angiogenesis in tumor growth and metastasis. Semin. Oncol. 2002, 29, 15–18. [Google Scholar]
- Folkman, J. What is the evidence that tumors are angiogenesis dependent? J. Natl. Cancer Inst. 1990, 82, 4–6. [Google Scholar] [CrossRef]
- Holmgren, L.; O'Reilly, M.S.; Folkman, J. Dormancy of micrometastases: Balanced proliferation and apoptosis in the presence of angiogenesis suppression. Nat. Med. 1995, 1, 149–153. [Google Scholar] [CrossRef]
- Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef]
- Bhat, T.A.; Singh, R.P. Tumor angiogenesis—A potential target in cancer chemoprevention. Food Chem. Toxicol. 2008, 46, 1334–1345. [Google Scholar] [CrossRef]
- Korpanty, G.; Smyth, E. Anti-VEGF Strategies—From Antibodies to Tyrosine Kinase Inhibitors: Background and Clinical Development in Human Cancer. Curr. Pharm. Des. 2012, 18, 2680–2701. [Google Scholar]
- Bergers, G.; Song, S.; Meyer-Morse, N.; Bergsland, E.; Hanahan, D. Benefits of targeting both pericytes and endothelial cells in the tumor vasculature with kinase inhibitors. J. Clin. Invest. 2003, 111, 1287–1295. [Google Scholar]
- Kamba, T.; McDonald, D.M. Mechanisms of adverse effects of anti-VEGF therapy for cancer. Br. J. Cancer 2007, 96, 1788–1795. [Google Scholar] [CrossRef]
- Summers, J.; Cohen, M.H.; Keegan, P.; Pazdur, R. FDA drug approval summary: Bevacizumab plus interferon for advanced renal cell carcinoma. Oncologist 2010, 15, 104–111. [Google Scholar] [CrossRef]
- Ramos, S. Cancer chemoprevention and chemotherapy: Dietary polyphenols and signalling pathways. Mol. Nutr. Food Res. 2008, 52, 507–526. [Google Scholar] [CrossRef]
- Prasad, S.; Phromnoi, K.; Yadav, V.R.; Chaturvedi, M.M.; Aggarwal, B.B. Targeting inflammatory pathways by flavonoids for prevention and treatment of cancer. Planta Med. 2010, 76, 1044–1063. [Google Scholar] [CrossRef]
- Kim, Y.W.; Kang, H.E.; Lee, M.G.; Hwang, S.J.; Kim, S.C.; Lee, C.H.; Kim, S.G. Liquiritigenin, a flavonoid aglycone from licorice, has a choleretic effect and the ability to induce hepatic transporters and phase-II enzymes. Am. J. Physiol. Gastrointest. Liver. Physiol. 2009, 296, G372–G381. [Google Scholar]
- Kim, Y.W.; Kim, Y.M.; Yang, Y.M.; Kay, H.Y.; Kim, W.D.; Lee, J.W.; Hwang, S.J.; Kim, S.G. Inhibition of LXRalpha-dependent steatosis and oxidative injury by liquiritigenin, a licorice flavonoid, as mediated with Nrf2 activation. Antioxid. Redox Signal. 2011, 14, 733–745. [Google Scholar] [CrossRef]
- Zhang, S.P.; Zhou, Y.J.; Liu, Y.; Cai, Y.Q. Effect of liquiritigenin, a flavanone existed from Radix glycyrrhizae on pro-apoptotic in SMMC-7721 cells. Food Chem. Toxicol. 2009, 47, 693–701. [Google Scholar] [CrossRef]
- Liu, C.; Wang, Y.; Xie, S.; Zhou, Y.; Ren, X.; Li, X.; Cai, Y. Liquiritigenin induces mitochondria-mediated apoptosis via cytochrome c release and caspases activation in HeLa Cells. Phytother. Res. 2011, 25, 277–283. [Google Scholar]
- Zhou, M.; Higo, H.; Cai, Y. Inhibition of hepatoma 22 tumor by Liquiritigenin. Phytother. Res. 2010, 24, 827–833. [Google Scholar]
- Xie, S.R.; Wang, Y.; Liu, C.W.; Luo, K.; Cai, Y.Q. Liquiritigenin Inhibits Serum-induced HIF-1alpha and VEGF Expression via the AKT/mTOR-p70S6K Signalling Pathway in HeLa Cells. Phytother. Res. 2011. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, S.; Liu, C.; Wu, Y.; Liu, Y.; Cai, Y. Inhibitory Effect of Liquiritigenin on Migration Via Downregulation ProMMP-2 and PI3K/Akt Signaling Pathway in Human Lung Adenocarcinoma A549 cells. Nutr. Cancer 2012, 64, 627–634. [Google Scholar] [CrossRef]
- Shultz, M.D.; Wilson, J.D.; Fuller, C.E.; Zhang, J.; Dorn, H.C.; Fatouros, P.P. Metallofullerene-based nanoplatform for brain tumor brachytherapy and longitudinal imaging in a murine orthotopic xenograft model. Radiology 2011, 261, 136–143. [Google Scholar]
- Tarallo, V.; Vesci, L.; Capasso, O.; Esposito, M.T.; Riccioni, T.; Pastore, L.; Orlandi, A.; Pisano, C.; De Falco, S. A placental growth factor variant unable to recognize vascular endothelial growth factor (VEGF) receptor-1 inhibits VEGF-dependent tumor angiogenesis via heterodimerization. Cancer Res. 2010, 70, 1804–1813. [Google Scholar]
- Kang, S.Y.; Watnick, R.S. Regulation of tumor dormancy as a function of tumor-mediated paracrine regulation of stromal Tsp-1 and VEGF expression. APMIS 2008, 116, 638–647. [Google Scholar] [CrossRef]
- Guan, P.; Howell-Jones, R.; Li, N.; Bruni, L.; De Sanjose, S.; Franceschi, S.; Clifford, G.M. Human papillomavirus (HPV) types in 115,789 HPV-positive women: A meta-analysis from cervical infection to cancer. Int. J. Cancer 2012. [Google Scholar] [CrossRef]
- Joo, Y.H.; Jung, C.K.; Sun, D.I.; Park, J.O.; Cho, K.J.; Kim, M.S. High-risk human papillomavirus and cervical lymph node metastasis in patients with oropharyngeal cancer. Head Neck 2012, 34, 10–14. [Google Scholar] [CrossRef]
- Hisamatsu, T.; Mabuchi, S.; Yoshino, K.; Fujita, M.; Enomoto, T.; Hamasaki, T.; Kimura, T. Prediction of Progression-Free Survival and Response to Paclitaxel Plus Carboplatin in Patients With Recurrent or Advanced Cervical Cancer. Int. J. Gynecol. Cancer 2012, 22, 623–629. [Google Scholar] [CrossRef]
- Mathur, R.S.; Mathur, S.P. Vascular endothelial growth factor (VEGF) up-regulates epidermal growth factor receptor (EGF-R) in cervical cancer in vitro: This action is mediated through HPV-E6 in HPV-positive cancers. Gynecol. Oncol. 2005, 97, 206–213. [Google Scholar] [CrossRef]
- Bachtiary, B.; Selzer, E.; Knocke, T.H.; Potter, R.; Obermair, A. Serum VEGF levels in patients undergoing primary radiotherapy for cervical cancer: Impact on progression-free survival. Cancer Lett. 2002, 179, 197–203. [Google Scholar] [CrossRef]
- Clere, N.; Faure, S.; Martinez, M.C.; Andriantsitohaina, R. Anticancer properties of flavonoids: Roles in various stages of carcinogenesis. Cardiovasc. Hematol. Agents Med. Chem. 2011, 9, 62–77. [Google Scholar]
- Gao, L.; Chen, L.; Fei, X.H.; Qiu, H.Y.; Zhou, H.; Wang, J.M. STI571 combined with vincristine greatly suppressed the tumor formation of multidrug-resistant K562 cells in a human-nude mice xenograft model. Chin. Med. J. (Engl) 2006, 119, 911–918. [Google Scholar]
- Villaume, K.; Blanc, M.; Gouysse, G.; Walter, T.; Couderc, C.; Nejjari, M.; Vercherat, C.; Cordier-Bussat, M.; Roche, C.; Scoazec, J.Y. VEGF secretion by neuroendocrine tumor cells is inhibited by octreotide and by inhibitors of the PI3K/AKT/mTOR pathway. Neuroendocrinology 2010, 91, 268–278. [Google Scholar] [CrossRef]
- Ferrara, N. Pathways mediating VEGF-independent tumor angiogenesis. Cytokine Growth Factor Rev. 2010, 21, 21–26. [Google Scholar] [CrossRef]
- Donmez, G.; Sullu, Y.; Baris, S.; Yildiz, L.; Aydin, O.; Karagoz, F.; Kandemir, B. Vascular endothelial growth factor (VEGF), matrix metalloproteinase-9 (MMP-9), and thrombospondin-1 (TSP-1) expression in urothelial carcinomas. Pathol. Res. Pract. 2009, 205, 854–847. [Google Scholar] [CrossRef]
- Karavasilis, V.; Malamou-Mitsi, V.; Briasoulis, E.; Tsanou, E.; Kitsou, E.; Kalofonos, H.; Fountzilas, G.; Fotsis, T.; Pavlidis, N. Angiogenesis in cancer of unknown primary: Clinicopathological study of CD34, VEGF and TSP-1. BMC Cancer 2005, 5, 25. [Google Scholar] [CrossRef] [Green Version]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, Y.; Xie, S.; Wang, Y.; Luo, K.; Wang, Y.; Cai, Y. Liquiritigenin Inhibits Tumor Growth and Vascularization in a Mouse Model of Hela Cells. Molecules 2012, 17, 7206-7216. https://doi.org/10.3390/molecules17067206
Liu Y, Xie S, Wang Y, Luo K, Wang Y, Cai Y. Liquiritigenin Inhibits Tumor Growth and Vascularization in a Mouse Model of Hela Cells. Molecules. 2012; 17(6):7206-7216. https://doi.org/10.3390/molecules17067206
Chicago/Turabian StyleLiu, Yuxin, Sirou Xie, Yu Wang, Kang Luo, Yang Wang, and Yunqing Cai. 2012. "Liquiritigenin Inhibits Tumor Growth and Vascularization in a Mouse Model of Hela Cells" Molecules 17, no. 6: 7206-7216. https://doi.org/10.3390/molecules17067206