Gamma-Mangostin, a Micronutrient of Mangosteen Fruit, Induces Apoptosis in Human Colon Cancer Cells

Abstract
:1. Introduction
2. Results and Discussion
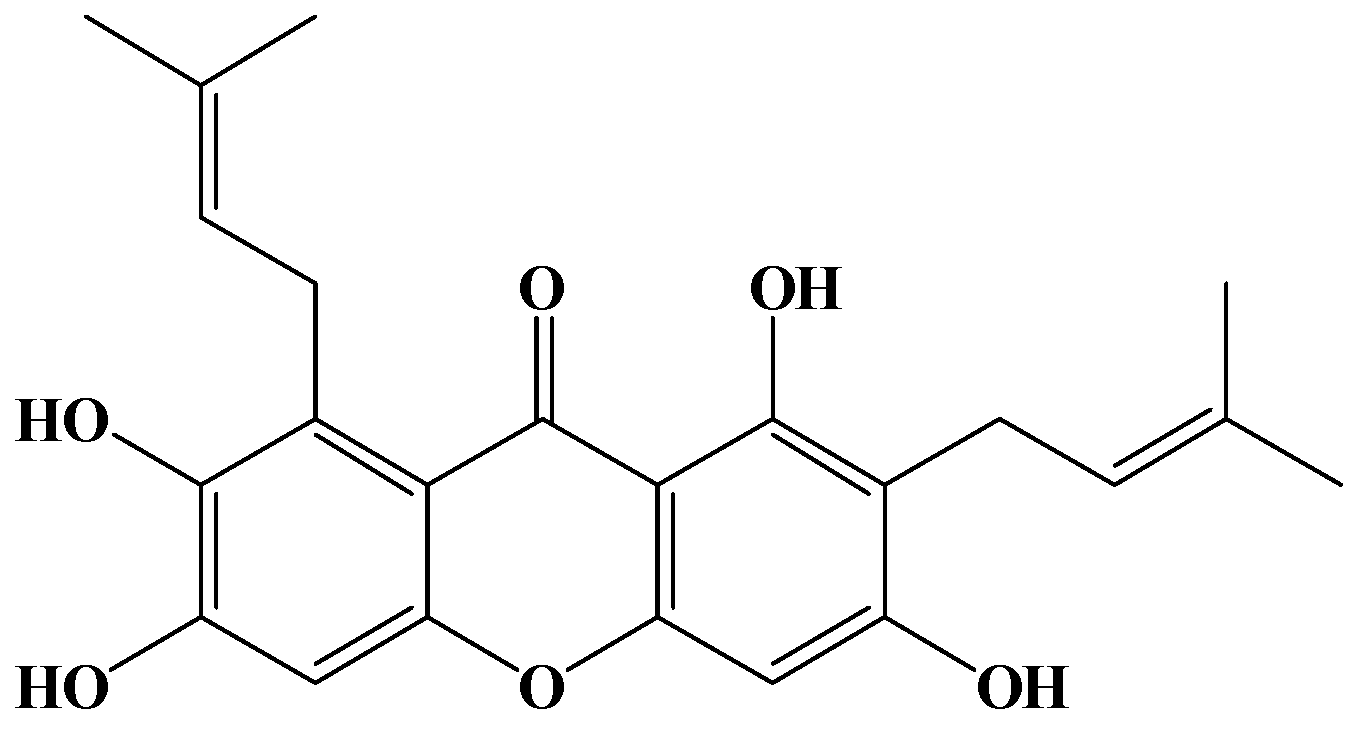
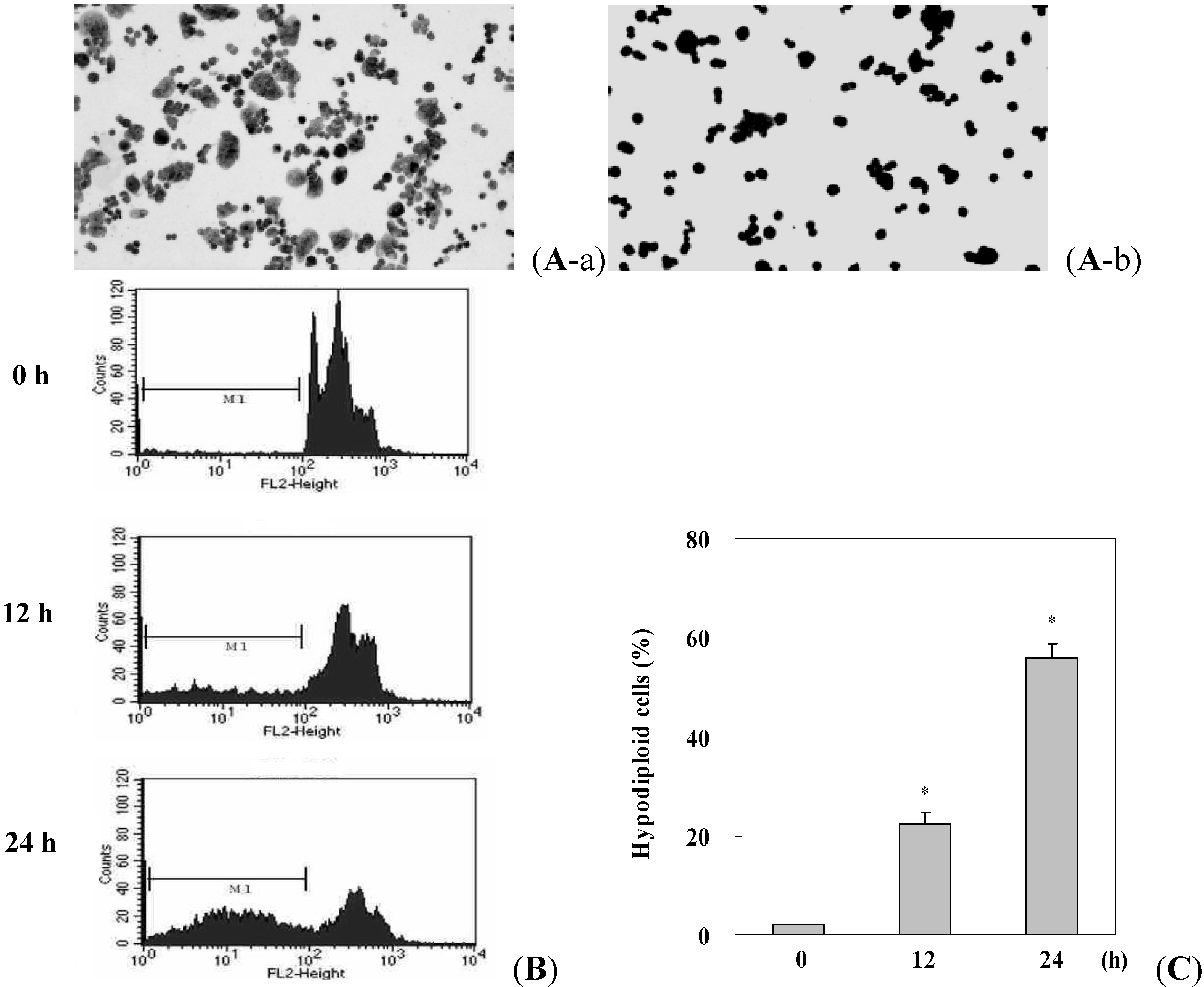
2.1. Antiproliferation of γ-Mangostin in HT29 Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| γ-Mangostin (μM) | Cytotoxicity (%) |
|---|---|
| 10 | 0.00 ± 4.67 |
| 20 | 13.16 ± 0.14 |
| 40 | 34.47 ± 3.13 |
| 80 | 80.01 ± 0.57 |
| 100 | 95.36 ± 3.25 |
| 200 | 99.90 ± 0.58 |
| IC50 (µM) | Cytotoxicity (%) | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| 68.48 ± 6.73 | 51.87 ± 4.53 | 69.15 ± 3.24 | 85.38 ± 4.75 |
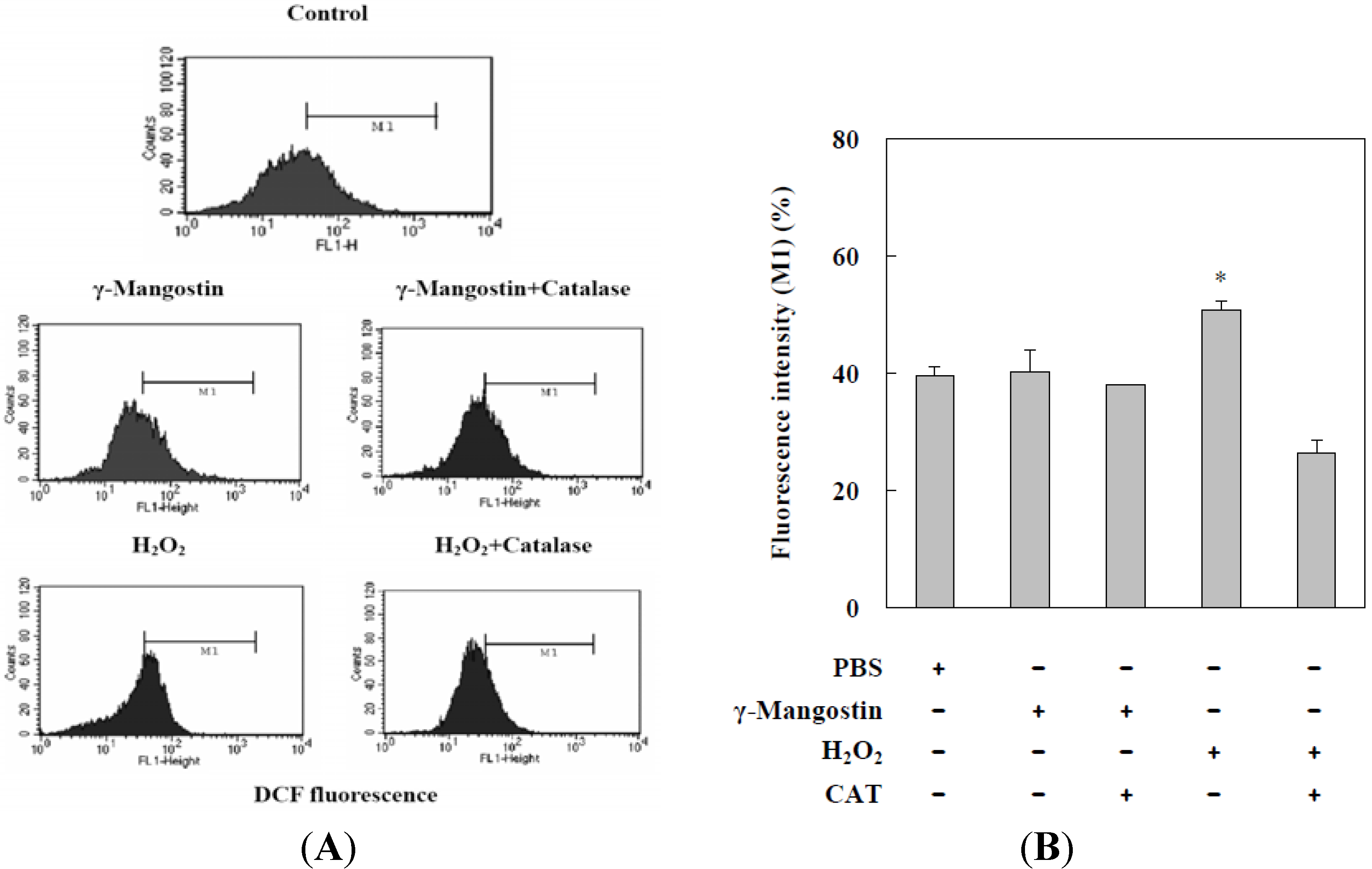
2.2. γ-Mangostin Induces the Apoptosis Process via Intracellular ROS Production


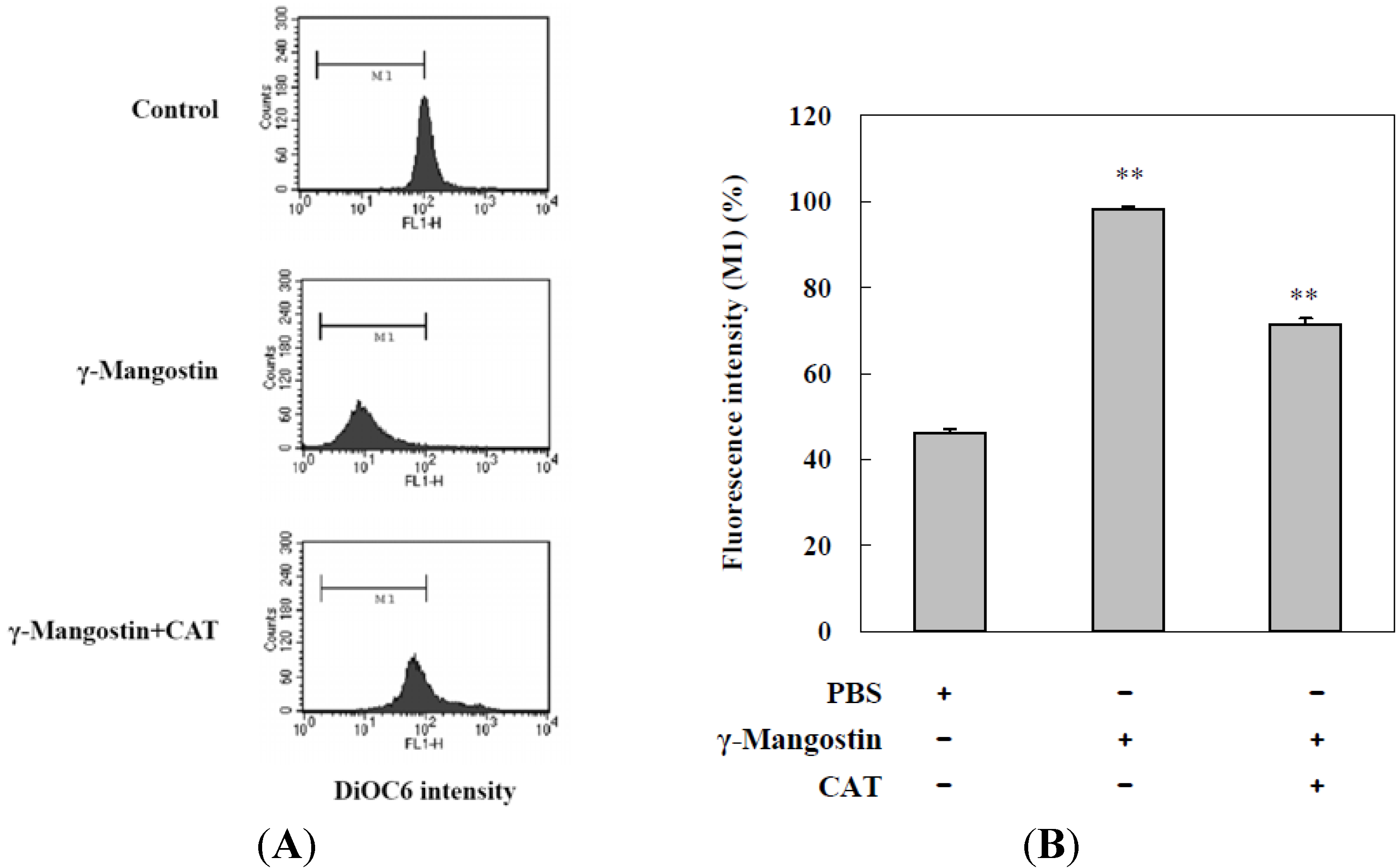
2.3. Cell Damage and ROS-Dependent Mitochondrial Dysfunction

3. Experimental
3.1. Chemicals
3.2. Extraction, Isolation, and Purification of γ-Mangostin
3.3. Cell Culture
3.4. Determination of Cell Viability by the MTT Assay
3.5. Morphological Study Using Fluorescence Microscopy
3.6. Apoptosis and Intracellular ROS Determination
3.7. Detection of the Mitochondrial Function
3.8. Statistical Analysis
4. Conclusions
Potential Conflicts of Interest
Acknowledgements
References
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Global cancer statistics, 2002. CA Cancer J. Clin. 2005, 55, 74–108. [Google Scholar] [CrossRef]
- Cunningham, D.; Atkin, W.; Lenz, H.J.; Lynch, H.T.; Minsky, B.; Nordlinger, B.; Starling, N. Colorectal cancer. Lancet 2010, 375, 1030–1047. [Google Scholar]
- Turktekin, M.; Konac, E.; Onen, H.I.; Alp, E.; Yilmaz, A.; Menevse, S. Evaluation of the effects of the flavonoid apigenin on apoptotic pathway gene expression on the colon cancer cell line (HT29). J. Med. Food 2011, 14, 1107–1117. [Google Scholar]
- Reyes-Zurita, F.J.; Pachon-Pena, G.; Lizarraga, D.; Rufino-Palomares, E.E.; Cascante, M.; Lupianez, J.A. The natural triterpene maslinic acid induces apoptosis in HT29 colon cancer cells by a JNK-p53-dependent mechanism. BMC Cancer 2011, 11, 154. [Google Scholar]
- Kang, H.J.; Youn, Y.K.; Hong, M.K.; Kim, L.S. Antiproliferation and redifferentiation in thyroid cancer cell lines by polyphenol phytochemicals. J. Korean Med. Sci. 2011, 26, 893–899. [Google Scholar]
- Shibata, M.A.; Iinuma, M.; Morimoto, J.; Kurose, H.; Akamatsu, K.; Okuno, Y.; Akao, Y.; Otsuki, Y. alpha-Mangostin extracted from the pericarp of the mangosteen (Garcinia mangostana Linn) reduces tumor growth and lymph node metastasis in an immunocompetent xenograft model of metastatic mammary cancer carrying a p53 mutation. BMC Med. 2011, 9, 69. [Google Scholar] [CrossRef]
- Chang, H.F.; Huang, W.T.; Chen, H.J.; Yang, L.L. Apoptotic effects of gamma-mangostin from the fruit hull of Garcinia mangostana on human malignant glioma cells. Molecules 2010, 15, 8953–8966. [Google Scholar] [CrossRef]
- Balunas, M.J.; Su, B.; Brueggemeier, R.W.; Kinghorn, A.D. Xanthones from the botanical dietary supplement mangosteen (Garcinia mangostana) with aromatase inhibitory activity. J. Nat. Prod. 2008, 71, 1161–1166. [Google Scholar] [CrossRef]
- Nabandith, V.; Suzui, M.; Morioka, T.; Kaneshiro, T.; Kinjo, T.; Matsumoto, K.; Akao, Y.; Iinuma, M.; Yoshimi, N. Inhibitory effects of crude alpha-mangostin, a xanthone derivative, on two different categories of colon preneoplastic lesions induced by 1,2-dimethylhydrazine in the rat. Asian Pac. J. Cancer Prev. APJCP 2004, 5, 433–438. [Google Scholar]
- Chen, S.X.; Wan, M.; Loh, B.N. Active constituents against HIV-1 protease from Garcinia mangostana. Planta Med. 1996, 62, 381–382. [Google Scholar] [CrossRef]
- Chairungsrilerd, N.; Furukawa, K.; Ohta, T.; Nozoe, S.; Ohizumi, Y. Histaminergic and serotonergic receptor blocking substances from the medicinal plant Garcinia mangostana. Planta Med. 1996, 62, 471–472. [Google Scholar] [CrossRef]
- Akao, Y.; Nakagawa, Y.; Iinuma, M.; Nozawa, Y. Anti-cancer effects of xanthones from pericarps of mangosteen. Int. J. Mol. Sci. 2008, 9, 355–370. [Google Scholar] [CrossRef]
- Chen, L.G.; Yang, L.L.; Wang, C.C. Anti-inflammatory activity of mangostins from Garcinia mangostana. Food Chem. Toxicol. 2008, 46, 688–693. [Google Scholar] [CrossRef]
- Chao, C.; Carmical, J.R.; Ives, K.L.; Wood, T.G.; Aronson, J.F.; Gomez, G.A.; Djukom, C.D.; Hellmich, M.R. CD133+ colon cancer cells are more interactive with the tumor microenvironment than CD133-cells. Lab. Invest. J. Technol. Methods Pathol. 2012, 92, 420–436. [Google Scholar] [CrossRef]
- Jeon, Y.K.; Kim, S.H.; Choi, S.H.; Kim, K.H.; Yoo, B.C.; Ku, J.L.; Park, J.G. Promoter hypermethylation and loss of CD133 gene expression in colorectal cancers. World J. Gastroenterol. 2010, 16, 3153–3160. [Google Scholar]
- Alley, M.C.; Scudiero, D.A.; Monks, A.; Hursey, M.L.; Czerwinski, M.J.; Fine, D.L.; Abbott, B.J.; Mayo, J.G.; Shoemaker, R.H.; Boyd, M.R. Feasibility of drug screening with panels of human tumor cell lines using a microculture tetrazolium assay. Cancer Res. 1988, 48, 589–601. [Google Scholar]
- Chen, Y.C.; Shen, S.C.; Lee, W.R.; Lin, H.Y.; Ko, C.H.; Shih, C.M.; Yang, L.L. Wogonin and fisetin induction of apoptosis through activation of caspase 3 cascade and alternative expression of p21 protein in hepatocellular carcinoma cells SK-HEP-1. Arch. Toxicol. 2002, 76, 351–359. [Google Scholar]
- Chow, J.M.; Huang, G.C.; Shen, S.C.; Wu, C.Y.; Lin, C.W.; Chen, Y.C. Differential apoptotic effect of wogonin and nor-wogonin via stimulation of ROS production in human leukemia cells. J. Cell Biochem. 2008, 103, 1394–1404. [Google Scholar] [CrossRef]
- Chairungsrilerd, N.; Furukawa, K.; Tadano, T.; Kisara, K.; Ohizumi, Y. Effect of gamma-mangostin through the inhibition of 5-hydroxy-tryptamine2A receptors in 5-fluoro-alpha-methyltryptamine-induced head-twitch responses of mice. Br. J. Pharmacol. 1998, 123, 855–862. [Google Scholar]
- Kakiuchi, N.; Hattori, M.; Nishizawa, M.; Yamagishi, T.; Okuda, T.; Namba, T. Studies on dental caries prevention by traditional medicines. VIII. Inhibitory effect of various tannins on glucan synthesis by glucosyltransferase from Streptococcus mutans. Chem. Pharm. Bull. (Tokyo) 1986, 34, 720–725. [Google Scholar]
- Nakatani, K.; Yamakuni, T.; Kondo, N.; Arakawa, T.; Oosawa, K.; Shimura, S.; Inoue, H.; Ohizumi, Y. gamma-Mangostin inhibits inhibitor-kappaB kinase activity and decreases lipopolysaccharide-induced cyclooxygenase-2 gene expression in C6 rat glioma cells. Mol. Pharmacol. 2004, 66, 667–674. [Google Scholar] [CrossRef]
- Yamakuni, T.; Aoki, K.; Nakatani, K.; Kondo, N.; Oku, H.; Ishiguro, K.; Ohizumi, Y. Garcinone B reduces prostaglandin E2 release and NF-kappaB-mediated transcription in C6 rat glioma cells. Neurosci. Lett. 2006, 394, 206–210. [Google Scholar] [CrossRef]
- Johnson, J.J.; Petiwala, S.M.; Syed, D.N.; Rasmussen, J.T.; Adhami, V.M.; Siddiqui, I.A.; Kohl, A.M.; Mukhtar, H. alpha-Mangostin, a xanthone from mangosteen fruit, promotes cell cycle arrest in prostate cancer and decreases xenograft tumor growth. Carcinogenesis 2012, 33, 413–419. [Google Scholar] [CrossRef]
- Jung, H.A.; Su, B.N.; Keller, W.J.; Mehta, R.G.; Kinghorn, A.D. Antioxidant xanthones from the pericarp of Garcinia mangostana (Mangosteen). J. Agric. Food Chem. 2006, 54, 2077–2082. [Google Scholar] [CrossRef]
- Doi, H.; Shibata, M.A.; Shibata, E.; Morimoto, J.; Akao, Y.; Iinuma, M.; Tanigawa, N.; Otsuki, Y. Panaxanthone isolated from pericarp of Garcinia mangostana L. suppresses tumor growth and metastasis of a mouse model of mammary cancer. Anticancer Res. 2009, 29, 2485–2495. [Google Scholar]
- Kondo, M.; Zhang, L.; Ji, H.; Kou, Y.; Ou, B. Bioavailability and antioxidant effects of a xanthone-rich Mangosteen (Garcinia mangostana) product in humans. J. Agric. Food Chem. 2009, 57, 8788–8792. [Google Scholar] [CrossRef]
- Kurose, H.; Shibata, M.A.; Iinuma, M.; Otsuki, Y. Alterations in cell cycle and induction of apoptotic cell death in breast cancer cells treated with alpha-mangostin extracted from mangosteen pericarp. J. Biomed. Biotechnol. 2012, 2012, 672428. [Google Scholar]
- Hung, S.H.; Shen, K.H.; Wu, C.H.; Liu, C.L.; Shih, Y.W. Alpha-mangostin suppresses PC-3 human prostate carcinoma cell metastasis by inhibiting matrix metalloproteinase-2/9 and urokinase-plasminogen expression through the JNK signaling pathway. J. Agric. Food Chem. 2009, 57, 1291–1298. [Google Scholar]
- Matsumoto, K.; Akao, Y.; Yi, H.; Ohguchi, K.; Ito, T.; Tanaka, T.; Kobayashi, E.; Iinuma, M.; Nozawa, Y. Preferential target is mitochondria in alpha-mangostin-induced apoptosis in human leukemia HL60 cells. Bioorg. Med. Chem. 2004, 12, 5799–5806. [Google Scholar]
- Sato, A.; Fujiwara, H.; Oku, H.; Ishiguro, K.; Ohizumi, Y. Alpha-mangostin induces Ca2+-ATPase-dependent apoptosis via mitochondrial pathway in PC12 cells. J. Pharmacol. Sci. 2004, 95, 33–40. [Google Scholar] [CrossRef]
- Matsumoto, K.; Akao, Y.; Ohguchi, K.; Ito, T.; Tanaka, T.; Iinuma, M.; Nozawa, Y. Xanthones induce cell-cycle arrest and apoptosis in human colon cancer DLD-1 cells. Bioorg. Med. Chem. 2005, 13, 6064–6069. [Google Scholar] [CrossRef]
- Chu, Y.F.; Sun, J.; Wu, X.; Liu, R.H. Antioxidant and antiproliferative activities of common vegetables. J. Agric. Food Chem. 2002, 50, 6910–6916. [Google Scholar]
- Block, G.; Patterson, B.; Subar, A. Fruit, vegetables, and cancer prevention: A review of the epidemiological evidence. Nutr. Cancer 1992, 18, 1–29. [Google Scholar] [CrossRef]
- Temple, N.J.; Gladwin, K.K. Fruit, vegetables, and the prevention of cancer: Research challenges. Nutrition 2003, 19, 467–470. [Google Scholar] [CrossRef]
- Sample Availability: Samples of γ-mangostin are available from the corresponding author.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chang, H.-F.; Yang, L.-L. Gamma-Mangostin, a Micronutrient of Mangosteen Fruit, Induces Apoptosis in Human Colon Cancer Cells. Molecules 2012, 17, 8010-8021. https://doi.org/10.3390/molecules17078010
Chang H-F, Yang L-L. Gamma-Mangostin, a Micronutrient of Mangosteen Fruit, Induces Apoptosis in Human Colon Cancer Cells. Molecules. 2012; 17(7):8010-8021. https://doi.org/10.3390/molecules17078010
Chicago/Turabian StyleChang, Hui-Fang, and Ling-Ling Yang. 2012. "Gamma-Mangostin, a Micronutrient of Mangosteen Fruit, Induces Apoptosis in Human Colon Cancer Cells" Molecules 17, no. 7: 8010-8021. https://doi.org/10.3390/molecules17078010