In Vitro and in Vivo Anticancer Activity of Aconitine on Melanoma Cell Line B16

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
2.1.1. B16 Mouse Melanoma Cells Were Sensitive to Aconitine


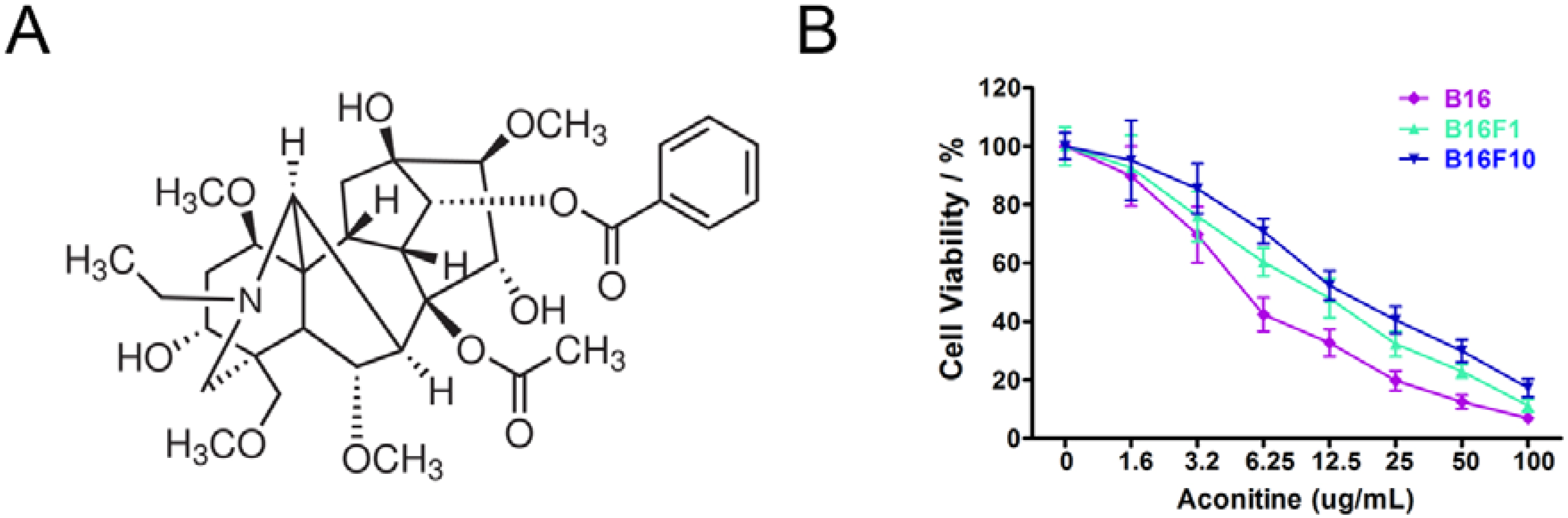{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Melanoma cell lines | IC50 (µg/mL) |
|---|---|
| B16 | 7.58 ± 0.99 *,# |
| B16F1 | 12.01 ± 1.12 |
| B16F10 | 17.09 ± 1.03 |
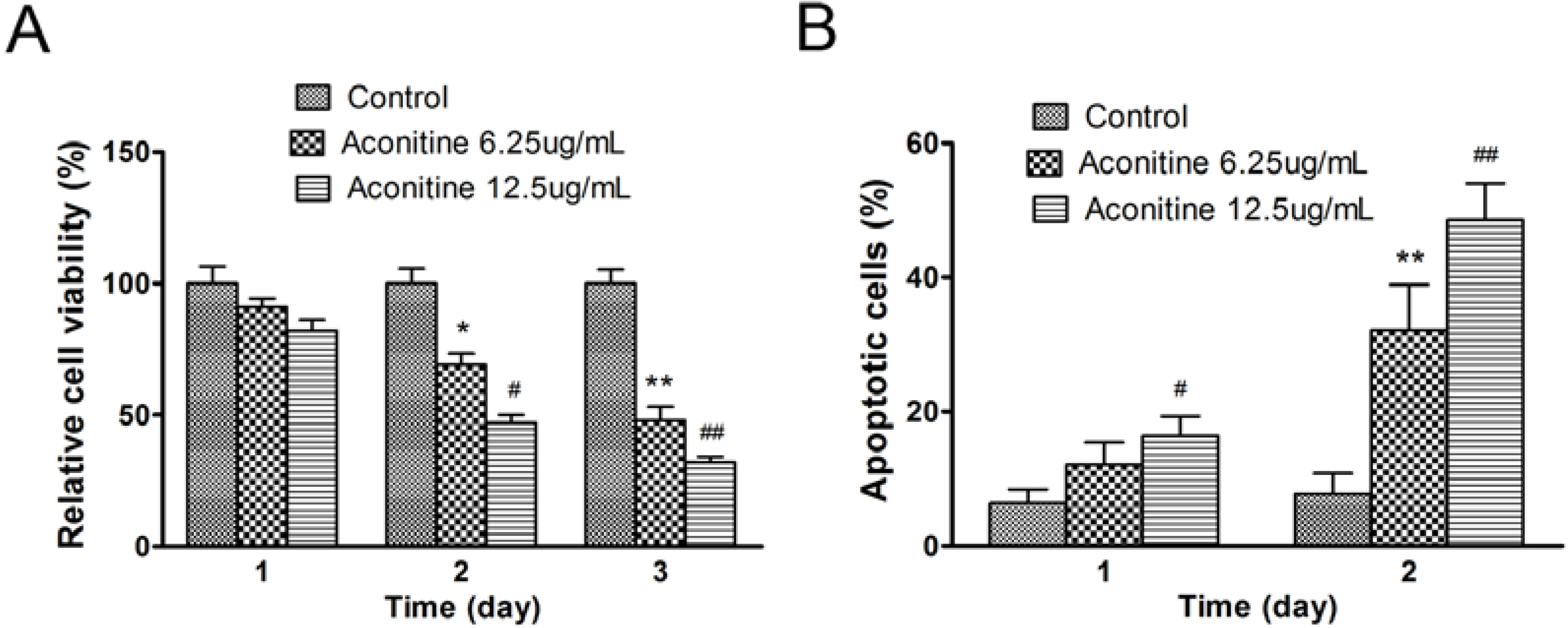
2.1.2. Aconitine Suppressed Cell Growth and Increased Apoptosis of B16 Cells
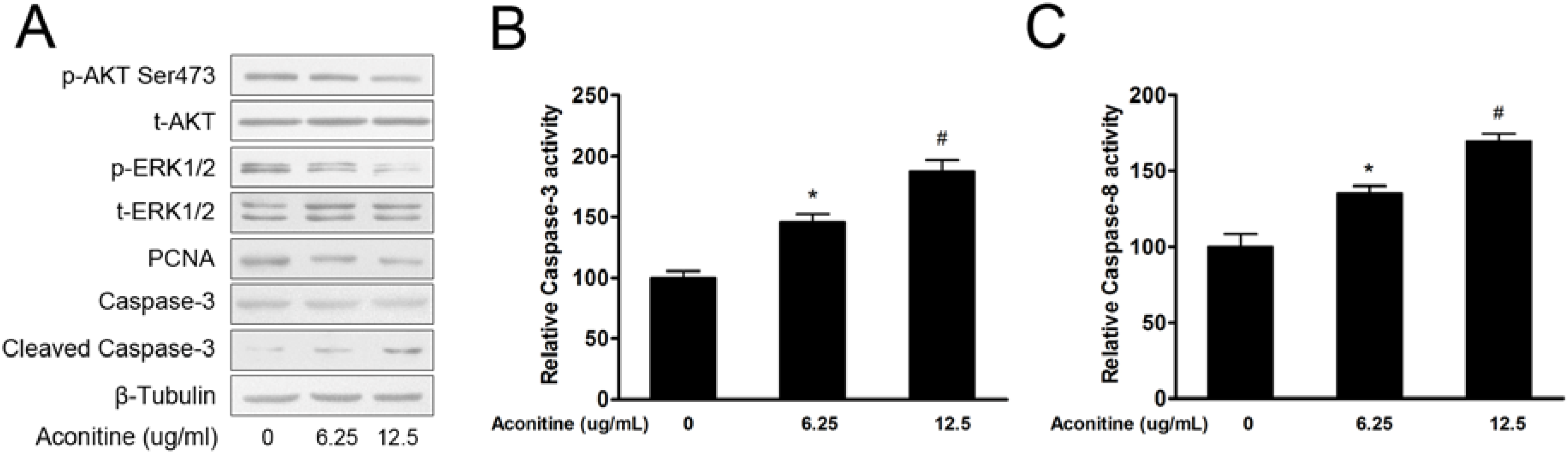
2.1.3. Aconitine Inactivated PI3K/AKT and MAPK/ERK1/2 Signaling Pathways


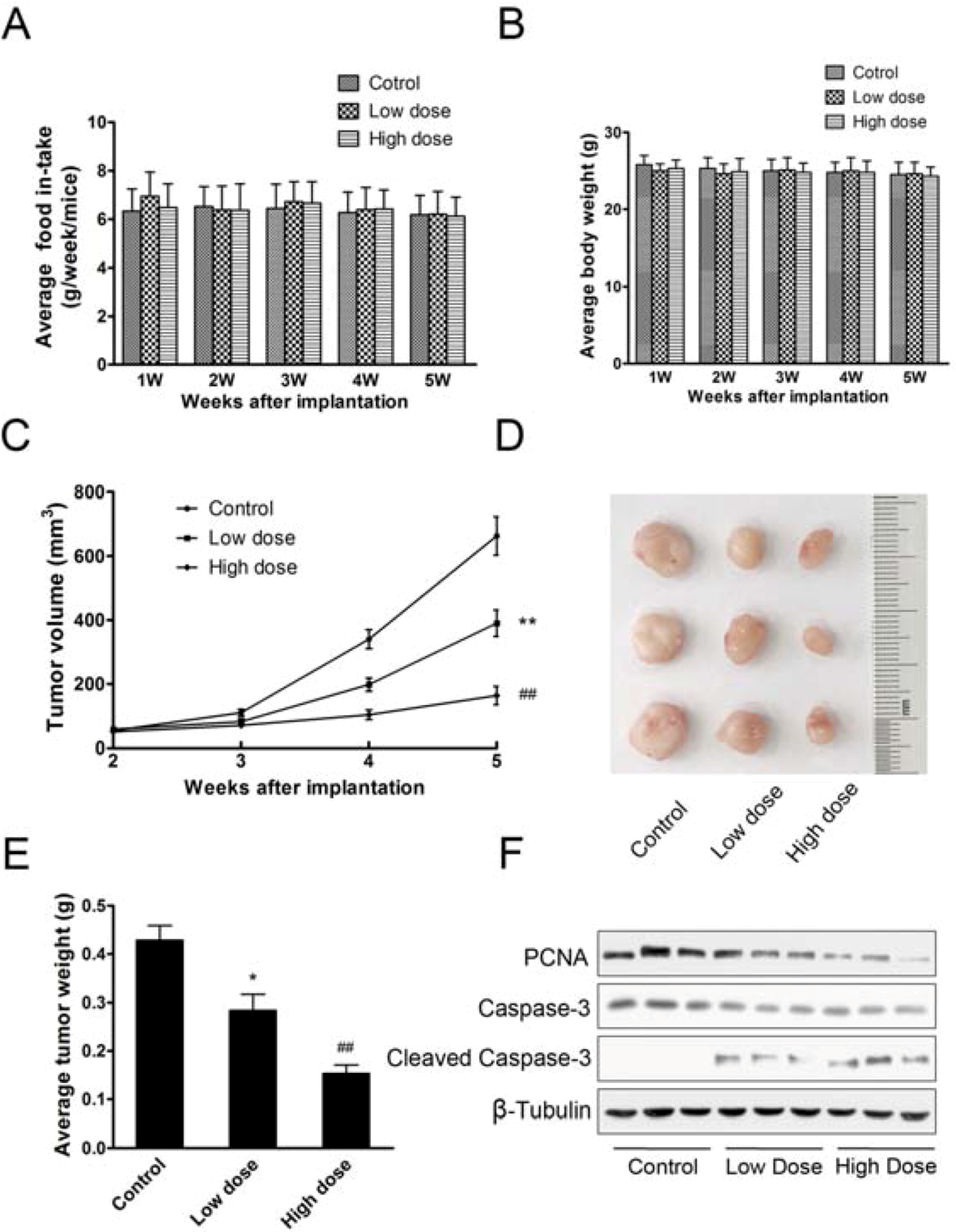
2.1.4. Aconitine Impaired in Vivo Tumor Growth

2.2. Discussion
3. Experimental
3.1. Cell Culture and Reagents
3.2. In Vitro Chemosensitivity Assay
3.3. Annexin-V apoptotic Assay
3.4. Western Blotting Assay
3.5. Caspase Activity
3.6. In Vivo Study
3.7. Statistical Analysis
4. Conclusions
Conflicts of Interest
Acknowledgments
- Sample Availability: Samples of the Aconitine are not available from the authors.
References
- Negroiu, G.; Dwek, R.A.; Petrescu, S.M. Tyrosinase-related protein-2 and -1 are trafficked on distinct routes in B16 melanoma cells. Biochem. Biophys. Res. Commun. 2005, 328, 914–921. [Google Scholar] [CrossRef]
- Zapas, J.L.; Coley, H.C.; Beam, S.L.; Brown, S.D.; Jablonski, K.A.; Elias, E.G. The risk of regional lymph node metastases in patients with melanoma less than 1.0 mm thick: Recommendations for sentinel lymph node biopsy. J. Am. Coll. Surg. 2003, 197, 403–407. [Google Scholar] [CrossRef]
- Manganoni, A.M.; Pavoni, L.; Facchetti, F.; Farisoglio, C.; Sereni, E.; Calzavara-Pinton, P. Melanoma in a patient in treatment with eculizumab. Ann. Hematol. 2012, 91, 135–136. [Google Scholar]
- Michot, C.; Dereure, O.; Baurain, J.F.; Brichard, V.; Guillot, B. Dermatomyositis occurring during treatment of a patient with metastatic melanoma. Melanoma Res. 2004, 14, 435–436. [Google Scholar] [CrossRef]
- Sutherland, C.M.; Chmiel, J.S.; Bieligk, S.; Henson, D.E.; Winchester, D.P. Patient characteristics, Treatment, and Outcome of unknown primary melanoma in the United States for the years 1981 and 1987. Am. Surg. 1996, 62, 400–406. [Google Scholar]
- Hegde, U.P.; Chakraborty, N.; Mukherji, B.; Grant Kels, J.M. Metastatic melanoma in the older patient: Immunologic insights and treatment outcomes. Expert Rev. Pharmacoecon. Outcomes Res. 2011, 11, 185–193. [Google Scholar] [CrossRef]
- Agarwala, S.S.; Hellstrand, K.; Gehlsen, K.; Naredi, P. Immunotherapy with histamine and interleukin 2 in malignant melanoma with liver metastasis. Cancer Immunol. Immunother. 2004, 53, 840–841. [Google Scholar]
- Masucci, G.V.; Mansson-Brahme, E.; Ragnarsson-Olding, B.; Nilsson, B.; Wagenius, G.; Hansson, J. Alternating chemo-immunotherapy with temozolomide and low-dose interleukin-2 in patients with metastatic melanoma. Melanoma Res. 2006, 16, 357–363. [Google Scholar] [CrossRef]
- Scheibenbogen, C.; Hunstein, W.; Keilholz, U. Vitiligo-like lesions following immunotherapy with IFN alpha and IL-2 in melanoma patients. Eur. J. Cancer 1994, 30A, 1209–1211. [Google Scholar]
- Middleton, M.; Hauschild, A.; Thomson, D.; Anderson, R.; Burdette-Radoux, S.; Gehlsen, K.; Hellstrand, K.; Naredi, P. Results of a multicenter randomized study to evaluate the safety and efficacy of combined immunotherapy with interleukin-2, Interferon-{alpha}2b and histamine dihydrochloride versus dacarbazine in patients with stage IV melanoma. Ann. Oncol. 2007, 18, 1691–167. [Google Scholar] [CrossRef]
- Hauschild, A.; Gogas, H.; Tarhini, A.; Middleton, M.R.; Testori, A.; Dreno, B.; Kirkwood, J.M. Practical guidelines for the management of interferon-alpha-2b side effects in patients receiving adjuvant treatment for melanoma: Expert opinion. Cancer 2008, 112, 982–994. [Google Scholar] [CrossRef]
- Rubin, K.M.; Vona, K.; Madden, K.; McGettigan, S.; Braun, I.M. Side effects in melanoma patients receiving adjuvant interferon alfa-2b therapy: A Nurse’s perspective. Support. Care Cancer 2012, 20, 1601–1611. [Google Scholar] [CrossRef]
- Capodanno, A.; Camerini, A.; Orlandini, C.; Baldini, E.; Resta, M.L.; Bevilacqua, G.; Collecchi, P. Dysregulated PI3K/Akt/PTEN pathway is a marker of a short disease-free survival in node-negative breast carcinoma. Hum. Pathol. 2009, 40, 1408–1417. [Google Scholar]
- Bhatt, A.P.; Bhende, P.M.; Sin, S.H.; Roy, D.; Dittmer, D.P.; Damania, B. Dual inhibition of PI3K and mTOR inhibits autocrine and paracrine proliferative loops in PI3K/Akt/mTOR-addicted lymphomas. Blood 2010, 115, 4455–4463. [Google Scholar]
- Babchia, N.; Calipel, A.; Mouriaux, F.; Faussat, A.M.; Mascarelli, F. The PI3K/Akt and mTOR/P70S6K signaling pathways in human uveal melanoma cells: interaction with B-Raf/ERK. Invest. Ophthalmol. Vis. Sci. 2010, 51, 421–429. [Google Scholar]
- Sinnberg, T.; Sauer, B.; Holm, P.; Spangler, B.; Kuphal, S.; Bosserhoff, A.; Schittek, B. MAPK and PI3K/AKT mediated YB-1 activation promotes melanoma cell proliferation which is counteracted by an autoregulatory loop. Exp. Dermatol. 2012, 21, 265–270. [Google Scholar] [CrossRef]
- Meier, F.; Schittek, B.; Busch, S.; Garbe, C.; Smalley, K.; Satyamoorthy, K.; Li, G.; Herlyn, M. The RAS/RAF/MEK/ERK and PI3K/AKT signaling pathways present molecular targets for the effective treatment of advanced melanoma. Front. Biosci. 2005, 10, 2986–3001. [Google Scholar] [CrossRef]
- Davies, M.A. The role of the PI3K-AKT pathway in melanoma. Cancer J. 2012, 18, 142–147. [Google Scholar] [CrossRef]
- Shin, J.H.; Choi, G.S.; Kang, W.H.; Myung, K.B. Sphingosine 1-phosphate triggers apoptotic signal for B16 melanoma cells via ERK and caspase activation. J. Korean Med. Sci. 2007, 22, 298–304. [Google Scholar]
- Guo, B.F.; Liu, S.; Ye, Y.Y.; Han, X.H. Inhibitory effects of osthole, Psoralen and aconitine on invasive activities of breast cancer MDA-MB-231BO cell line and the mechanisms. Zhong Xi Yi Jie He Xue Bao 2011, 9, 1110–1117. [Google Scholar] [CrossRef]
- Fan, Y.F.; Xie, Y.; Liu, L.; Ho, H.M.; Wong, Y.F.; Liu, Z.Q.; Zhou, H. Paeoniflorin reduced acute toxicity of aconitine in rats is associated with the pharmacokinetic alteration of aconitine. J. Ethnopharmacol. 2012, 141, 701–708. [Google Scholar] [CrossRef]
- Xiao, K.; Wang, L.; Liu, Y.; Peng, C.; Yan, G.; Zhang, J.; Zhuo, Y.; Li, H. Study of aconitine toxicity in rat embryos in vitro. Birth Defects Res. B Dev. Reprod. Toxicol. 2007, 80, 208–212. [Google Scholar] [CrossRef]
- Ruth, M.C.; Xu, Y.; Maxwell, I.H.; Ahn, N.G.; Norris, D.A.; Shellman, Y.G. RhoC promotes human melanoma invasion in a PI3K/Akt-dependent pathway. J. Invest. Dermatol. 2006, 126, 862–868. [Google Scholar] [CrossRef]
- Passarella, R.J.; Zhou, L.; Phillips, J.G.; Wu, H.; Hallahan, D.E.; Diaz, R. Recombinant peptides as biomarkers for tumor response to molecular targeted therapy. Clin. Cancer Res. 2009, 15, 6421–649. [Google Scholar]
- Ji, X.M.; Ouyang, B.; Liu, H.; Liu, G.W.; Wu, Z.C.; Yu, H.Y.; Wang, C.Y.; Wang, Z.X.; Wang, W.P. In vitro and in vivo inhibitory effect of the combination of Wenxia Changfu formula [see text] with cisplatin in non-small cell lung cancer. Chin. J. Integr. Med. 2011, 17, 908–916. [Google Scholar]
- Ogawa, K.; Sun, C.; Horii, A. Exploration of genetic alterations in human endometrial cancer and melanoma: Distinct tumorigenic pathways that share a frequent abnormal PI3K/AKT cascade. Oncol. Rep. 2005, 14, 1481–1485. [Google Scholar]
- Meier, F.; Busch, S.; Lasithiotakis, K.; Kulms, D.; Garbe, C.; Maczey, E.; Herlyn, M.; Schittek, B. Combined targeting of MAPK and AKT signalling pathways is a promising strategy for melanoma treatment. Br. J. Dermatol. 2007, 156, 1204–1213. [Google Scholar] [CrossRef]
- Isenovic, E.R.; Kedees, M.H.; Tepavcevic, S.; Milosavljevic, T.; Koricanac, G.; Trpkovic, A.; Marche, P. Role of PI3K/AKT, cPLA2 and ERK1/2 signaling pathways in insulin regulation of vascular smooth muscle cells proliferation. Cardiovasc. Hematol. Disord. Drug Targets 2009, 9, 172–180. [Google Scholar] [CrossRef]
- Gentilini, D.; Busacca, M.; Di Francesco, S.; Vignali, M.; Vigano, P.; Di Blasio, A.M. PI3K/Akt and ERK1/2 signalling pathways are involved in endometrial cell migration induced by 17beta-estradiol and growth factors. Mol. Hum. Reprod. 2007, 13, 317–322. [Google Scholar] [CrossRef]
- Dai, J.; Peng, L.; Fan, K.; Wang, H.; Wei, R.; Ji, G.; Cai, J.; Lu, B.; Li, B.; Zhang, D.; et al. Osteopontin induces angiogenesis through activation of PI3K/AKT and ERK1/2 in endothelial cells. Oncogene 2009, 28, 3412–3422. [Google Scholar] [CrossRef]
- Heshe, D.; Hoogestraat, S.; Brauckmann, C.; Karst, U.; Boos, J.; Lanvers-Kaminsky, C. Dichloroacetate metabolically targeted therapy defeats cytotoxicity of standard anticancer drugs. Cancer Chemother. Pharmacol. 2011, 67, 647–655. [Google Scholar] [CrossRef]
- Olszewski, U.; Hamilton, G. Mechanisms of cytotoxicity of anticancer titanocenes. Anticancer Agents Med. Chem. 2010, 10, 302–311. [Google Scholar]
- Mockey, M.; Bourseau, E.; Chandrashekhar, V.; Chaudhuri, A.; Lafosse, S.; Le Cam, E.; Quesniaux, V.F.; Ryffel, B.; Pichon, C.; Midoux, P. mRNA-based cancer vaccine: Prevention of B16 melanoma progression and metastasis by systemic injection of MART1 mRNA histidylated lipopolyplexes. Cancer Gene Ther. 2007, 14, 802–814. [Google Scholar] [CrossRef]
- Park, Y.; Ruan, F.; Hakomori, S.; Igarashi, Y. Cooperative inhibitory effect of n,n,n-trimethylsphingosine and sphingosine-1-phosphate, co-incorporated in liposomes, on b16 melanoma cell metastasis - cell-membrane signaling as a target in cancer-therapy. Int. J. Oncol. 1995, 7, 487–494. [Google Scholar]
- Katz, M.H.; Takimoto, S.; Spivack, D.; Moossa, A.R.; Hoffman, R.M.; Bouvet, M. An imageable highly metastatic orthotopic red fluorescent protein model of pancreatic cancer. Clin. Exp. Metastasis 2004, 21, 7–12. [Google Scholar] [CrossRef]
- Wen, Z.S.; Zhang, T.J.; Rong, T.H. Clinical adjuvant chemotherapy base on chemosensitivity in nude mouse subrenal capsule assay for non-small cell lung cancer. Ai Zheng 2003, 22, 1355–1358. [Google Scholar]
- Valeri, N.; Gasparini, P.; Braconi, C.; Paone, A.; Lovat, F.; Fabbri, M.; Sumani, K.M.; Alder, H.; Amadori, D.; Patel, T.; et al. MicroRNA-21 induces resistance to 5-fluorouracil by down-regulating human DNA MutS homolog 2 (hMSH2). Proc. Natl. Acad. Sci. USA 2010, 107, 21098–21103. [Google Scholar]
- Meng, Z.X.; Nie, J.; Ling, J.J.; Sun, J.X.; Zhu, Y.X.; Gao, L.; Lv, J.H.; Zhu, D.Y.; Sun, Y.J.; Han, X. Activation of liver X receptors inhibits pancreatic islet beta cell proliferation through cell cycle arrest. Diabetologia 2009, 52, 125–135. [Google Scholar] [CrossRef]
- Borralho, P.M.; Kren, B.T.; Castro, R.E.; da Silva, I.B.; Steer, C.J.; Rodrigues, C.M. MicroRNA-143 reduces viability and increases sensitivity to 5-fluorouracil in HCT116 human colorectal cancer cells. FEBS J. 2009, 276, 6689–6700. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Du, J.; Lu, X.; Long, Z.; Zhang, Z.; Zhu, X.; Yang, Y.; Xu, J. In Vitro and in Vivo Anticancer Activity of Aconitine on Melanoma Cell Line B16. Molecules 2013, 18, 757-767. https://doi.org/10.3390/molecules18010757
Du J, Lu X, Long Z, Zhang Z, Zhu X, Yang Y, Xu J. In Vitro and in Vivo Anticancer Activity of Aconitine on Melanoma Cell Line B16. Molecules. 2013; 18(1):757-767. https://doi.org/10.3390/molecules18010757
Chicago/Turabian StyleDu, Juan, Xiaonian Lu, Ziwen Long, Zhen Zhang, Xiaohua Zhu, Yongsheng Yang, and Jinhua Xu. 2013. "In Vitro and in Vivo Anticancer Activity of Aconitine on Melanoma Cell Line B16" Molecules 18, no. 1: 757-767. https://doi.org/10.3390/molecules18010757