Elicitation of Induced Resistance against Pectobacterium carotovorum and Pseudomonas syringae by Specific Individual Compounds Derived from Native Korean Plant Species

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
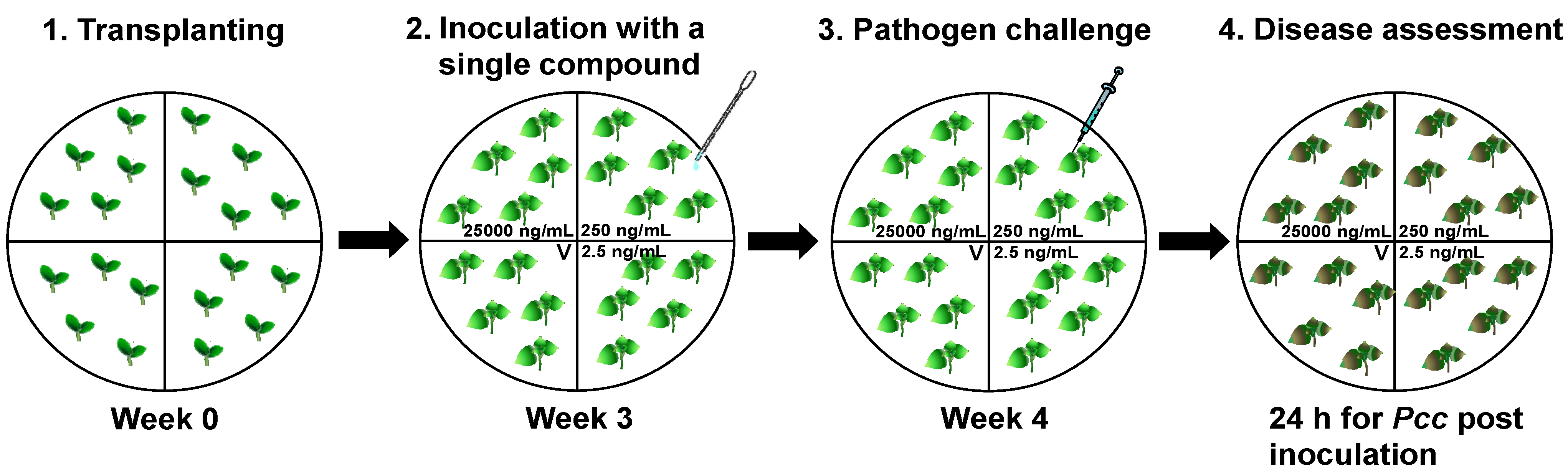
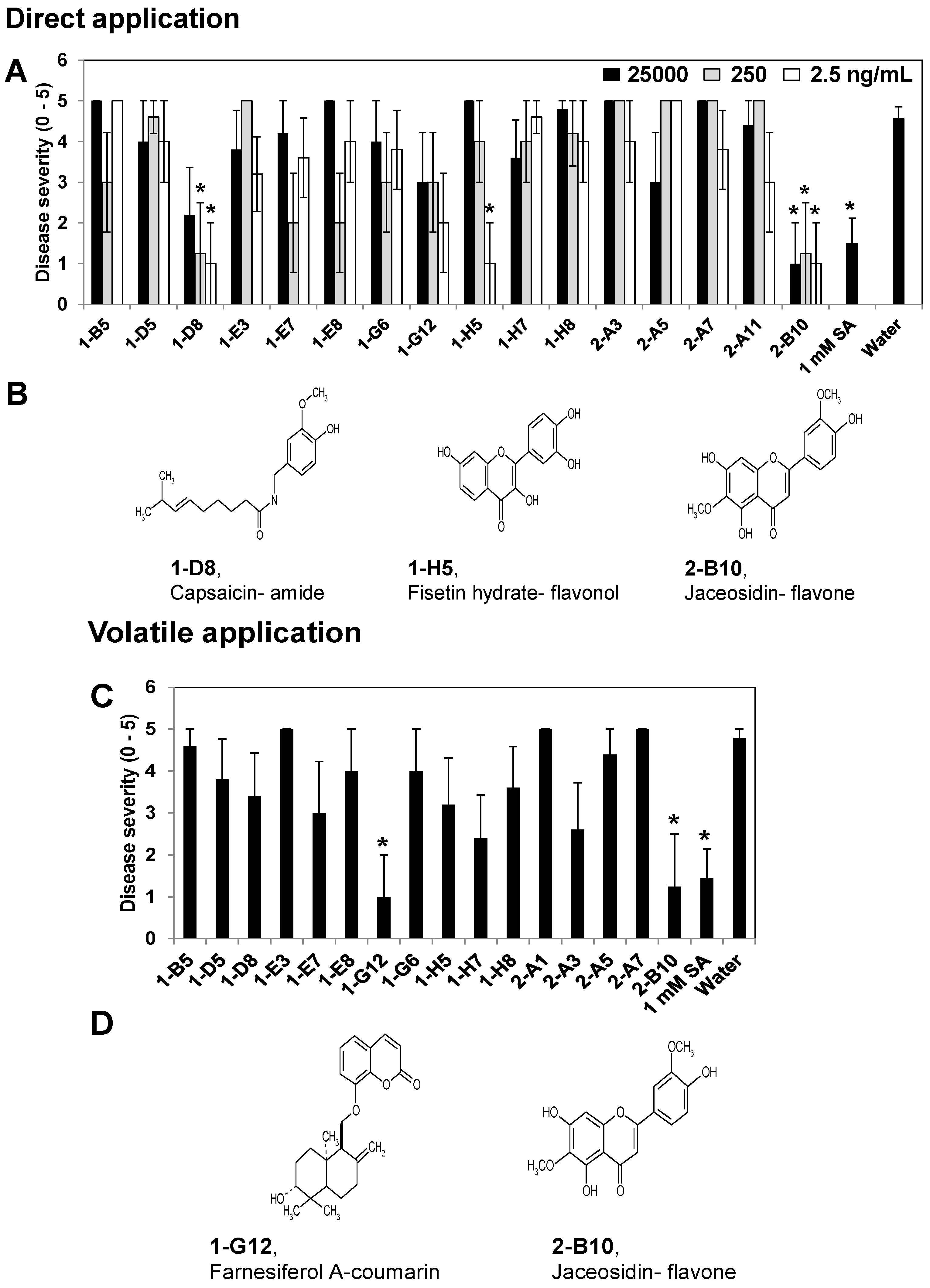
2.1. Screening Specific Compounds from Plant Extracts that Elicited Induced Resistance in the X-Plate System


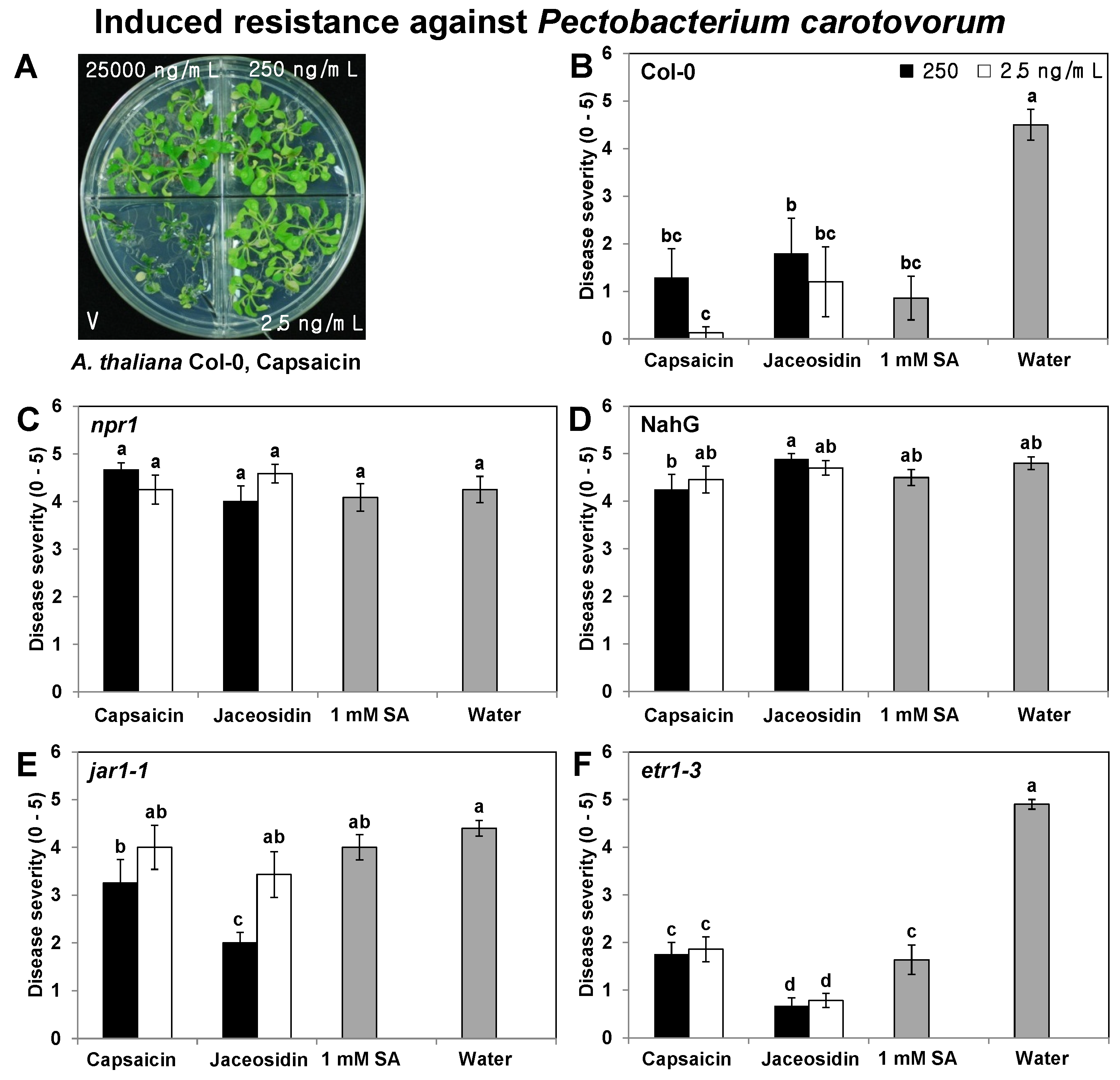
2.2. Elicitation of Induced Resistance by Capsaicin and Jaceosidin in Col-0, npr1, NahG, jar1-1, and etr1-3 Plants

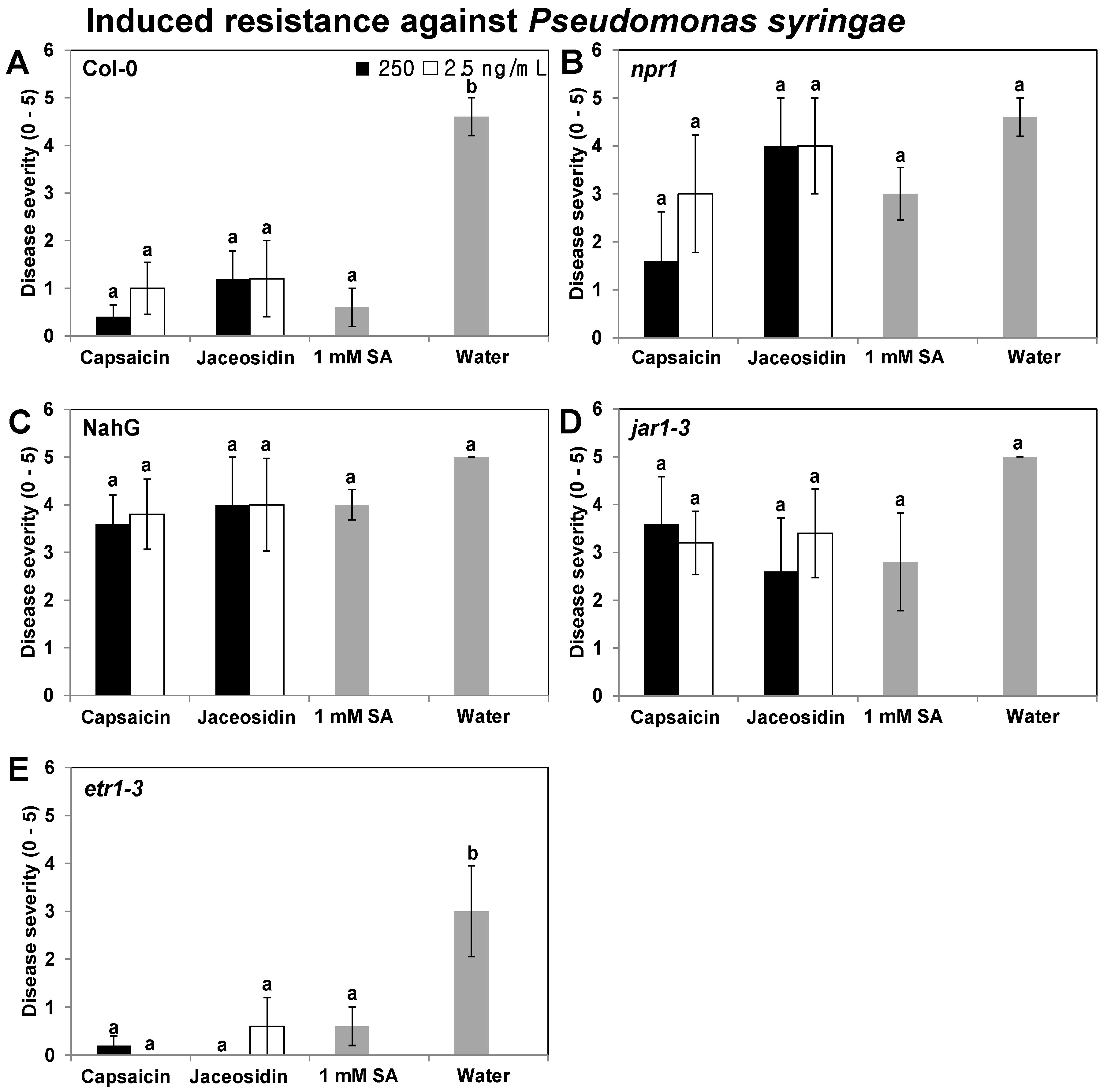
2.3. Elicitation of Induced Resistance in Col-0, NahG, npr1, jar1-1, and etr1-3 Plants by Capsaicin and Jaceosidin against the Biotrophic Pathogen P. syringae pv. tomato
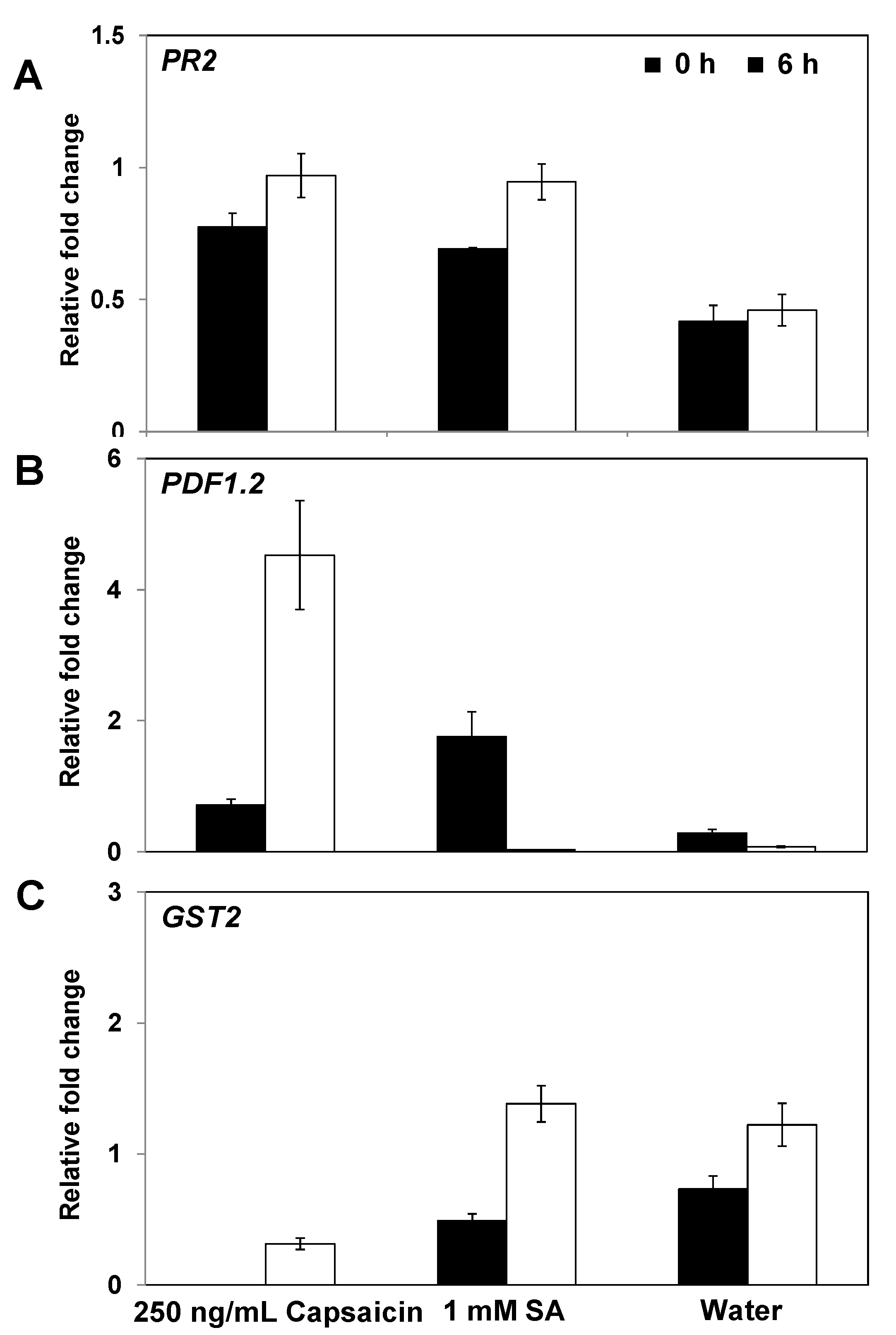
2.4. Expression of Defense-Related Genes in Response to Capsaicin


3. Experimental
3.1. Plant Preparation and Disease Assay
3.2. Plant Extraction and Identification
3.3. Direct Inhibition Assay
3.4. Induced Resistance in npr1, NahG, jar1-1, and etr1-3 Plants Using Specific Compounds
3.5. Quantitative RT-PCR
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Schneider, M.; Schweizer, P.; Meuwly, P.; Métraux, J. Systemic acquired resistance in plants. Int. Rev. Cytol. 1996, 168, 303–340. [Google Scholar] [CrossRef]
- Ross, A.F. Systemic acquired resistance induced by localized virus infections in plants. Virology 1961, 14, 340–358. [Google Scholar] [CrossRef]
- Hunt, M.D.; Ryals, J.A.; Reinhardt, D.D. Systemic acquired resistance signal transduction. Crit. Rev. Plant Sci. 1996, 15, 583–606. [Google Scholar]
- Ryals, J.A.; Neuenschwander, U.H.; Willits, M.G.; Molina, A.; Steiner, H.-Y.; Hunt, M.D. Systemic acquired resistance. Plant Cell 1996, 8, 1809. [Google Scholar]
- Métraux, J.; Ahl Goy, P.; Staub, T.; Speich, J.; Steinemann, A.; Ryals, J.; Ward, E. Induced systemic resistance in cucumber in response to 2,6-dichloro-isonicotinic acid and pathogens. Curr. Plant Sci. Biotechnol. Agric. 1991, 1, 432–439. [Google Scholar]
- Molina, A.; Hunt, M.D.; Ryals, J.A. Impaired fungicide activity in plants blocked in disease resistance signal transduction. Plant Cell 1998, 10, 1903–1914. [Google Scholar]
- Uknes, S.; Mauch-Mani, B.; Moyer, M.; Potter, S.; Williams, S.; Dincher, S.; Chandler, D.; Slusarenko, A.; Ward, E.; Ryals, J. Acquired resistance in Arabidopsis. Plant Cell 1992, 4, 645–656. [Google Scholar]
- Hammerschmidt, R. Induced disease resistance: How do induced plants stop pathogens? Physiol. Mol. Plant Pathol. 1999, 55, 77–84. [Google Scholar] [CrossRef]
- Ward, E.R.; Uknes, S.J.; Williams, S.C.; Dincher, S.S.; Wiederhold, D.L.; Alexander, D.C.; Ahl-Goy, P.; Métraux, J.-P.; Ryals, J.A. Coordinate gene activity in response to agents that induce systemic acquired resistance. Plant Cell 1991, 3, 1085–1094. [Google Scholar]
- Sanchez, L.; Courteaux, B.; Hubert, J.; Kauffmann, S.; Renault, J.-H.; Clément, C.; Baillieul, F.; Dorey, S. Rhamnolipids elicit defense responses and induce disease resistance against biotrophic, hemibiotrophic, and necrotrophic pathogens that require different signaling pathways in arabidopsis and highlight a central role for salicylic acid. Plant Physiol. 2012, 160, 1630–1641. [Google Scholar] [CrossRef]
- Makandar, R.; Nalam, V.; Chaturvedi, R.; Jeannotte, R.; Sparks, A.A.; Shah, J. Involvement of salicylate and jasmonate signaling pathways in arabidopsis interaction with Fusarium graminearum. Mol. Plant Microbe Interact. 2010, 23, 861–870. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Ryu, C.-M.; Zhang, S. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Leon-Reyes, A.; van der Ent, S.; van Wees, S.C. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef]
- Fawcett, C.; Spencer, D. Plant chemotherapy with natural products. Annu. Rev. Phytopathol. 1970, 8, 403–418. [Google Scholar] [CrossRef]
- Komatsu, K.; Nagayama, Y.; Tanaka, K.; Ling, Y.; Cai, S.-Q.; Omote, T.; Meselhy, M.R. Comparative study of chemical constituents of rhubarb from different origins. Chem. Pharm. Bull. 2006, 54, 1491–1499. [Google Scholar] [CrossRef]
- Izhaki, I. Emodin—A secondary metabolite with multiple ecological functions in higher plants. New Phytol. 2002, 155, 205–217. [Google Scholar] [CrossRef]
- Kim, Y.-M.; Lee, C.-H.; Kim, H.-G.; Lee, H.-S. Anthraquinones isolated from Cassia tora (Leguminosae) seed show an antifungal property against phytopathogenic fungi. J. Agric. Food Chem. 2004, 52, 6096–6100. [Google Scholar] [CrossRef]
- Mosch, J.; Klingauf, F.; Zeller, W. On the effect of plant extracts against fireblight (Erwinia amylovora). V Int. Workshop Fireblight ISHS Acta Horticult. 1989, 273, 355–362. [Google Scholar]
- Sasidharan, S.; Chen, Y.; Saravanan, D.; Sundram, K.; Latha, L.Y. Extraction, isolation and characterization of bioactive compounds from plants’ extracts. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 1–10. [Google Scholar]
- Noutoshi, Y.; Okazaki, M.; Kida, T.; Nishina, Y.; Morishita, Y.; Ogawa, T.; Suzuki, H.; Shibata, D.; Jikumaru, Y.; Hanada, A. Novel plant immune-priming compounds identified via high-throughput chemical screening target salicylic acid glucosyltransferases in Arabidopsis. Plant Cell 2012, 24, 3795–3804. [Google Scholar] [CrossRef]
- Anderson, J.P.; Thatcher, L.F.; Singh, K.B. Plant defence responses: Conservation between models and crops. Funct. Plant Biol. 2005, 32, 21–34. [Google Scholar] [CrossRef]
- Anderson, J.P.; Singh, K.B. Interactions of Arabidopsis and M. truncatula with the same Pathogens differ in dependence on ethylene and ethylene response factors. Plant Signal.Behav. 2011, 6, 551–552. [Google Scholar] [CrossRef]
- Laluk, K.; Mengiste, T. Necrotroph attacks on plants: Wanton destruction or covert extortion? The Arabidopsis book. Am. Soc. Plant Biol. 2010, 8, e0136. [Google Scholar]
- Vanneste, J.L. Fire Blight: The Disease and Its Causative Agent, Erwinia Amylovora; CABI: Boston, MA, USA, 2000. [Google Scholar]
- Bolton, M.D.; Thomma, B.P.; Nelson, B.D. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef]
- Bhat, K.; Masood, S.; Bhat, N.; Bhat, M.A.; Razvi, S.; Mir, M.; Akhtar, S.; Wani, N.; Habib, M. Current status of post harvest soft rot in vegetables: A review. Asian J. Plant Sci. 2010, 9, 200–208. [Google Scholar] [CrossRef]
- Sivasithamparam, K.; Barbetti, M.J.; Li, H. Recurring challenges from a necrotrophic fungal plant pathogen: A case study with Leptosphaeria maculans (causal agent of blackleg disease inBrassicas) in Western Australia. Ann. Bot. 2005, 96, 363–377. [Google Scholar] [CrossRef]
- Brkljacic, J.; Grotewold, E.; Scholl, R.; Mockler, T.; Garvin, D.F.; Vain, P.; Brutnell, T.; Sibout, R.; Bevan, M.; Budak, H. Brachypodium as a model for the grasses: Today and the future. Plant Physiol. 2011, 157, 3–13. [Google Scholar] [CrossRef]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Volatile C6-aldehydes and allo-ocimene activate defense genes and induce resistance against Botrytis cinerea in Arabidopsis thaliana. Plant Cell Physiol. 2005, 46, 1093–1102. [Google Scholar] [CrossRef]
- Seo, H.S.; Song, J.T.; Cheong, J.-J.; Lee, Y.-H.; Lee, Y.-W.; Hwang, I.; Lee, J.S.; Do Choi, Y. Jasmonic acid carboxyl methyltransferase: A key enzyme for jasmonate-regulated plant responses. Proc. Natl. Acad. Sci. USA 2001, 98, 4788–4793. [Google Scholar]
- Ozawa, R.; Arimura, G.-I.; Takabayashi, J.; Shimoda, T.; Nishioka, T. Involvement of jasmonate-and salicylate-related signaling pathways for the production of specific herbivore-induced volatiles in plants. Plant Cell Physiol. 2000, 41, 391–398. [Google Scholar] [CrossRef]
- Arimura, G.I.; Ozawa, R.; Nishioka, T.; Boland, W.; Koch, T.; Kühnemann, F.; Takabayashi, J. Herbivore-induced volatiles induce the emission of ethylene in neighboring lima bean plants. Plant J. 2002, 29, 87–98. [Google Scholar] [CrossRef]
- Farmer, E.E.; Ryan, C.A. Interplant communication: Airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves. Proc. Natl. Acad. Sci. USA 1990, 87, 7713–7716. [Google Scholar] [CrossRef]
- Achuo, E.; Audenaert, K.; Meziane, H.; Höfte, M. The salicylic acid-dependent defence pathway is effective against different pathogens in tomato and tobacco. Plant Pathol. 2004, 53, 65–72. [Google Scholar] [CrossRef]
- Lu, H.; Rate, D.N.; Song, J.T.; Greenberg, J.T. ACD6, a novel ankyrin protein, is a regulator and an effector of salicylic acid signaling in the Arabidopsis defense response. Plant Cell 2003, 15, 2408–2420. [Google Scholar] [CrossRef]
- Bate, N.J.; Rothstein, S.J. C6-volatiles derived from the lipoxygenase pathway induce a subset of defense-related genes. Plant J. 2001, 16, 561–569. [Google Scholar] [CrossRef]
- Kessler, A.; Halitschke, R.; Diezel, C.; Baldwin, I.T. Priming of plant defense responses in nature by airborne signaling between Artemisia tridentata and Nicotiana attenuata. Oecologia 2006, 148, 280–292. [Google Scholar] [CrossRef]
- Korea Chemical Bank. Available online: http://www.chembank.org/ (accessed on 14 August 2012).
- Arai, Y.; Watanabe, S.; Kimira, M.; Shimoi, K.; Mochizuki, R.; Kinae, N. Dietary intakes of flavonols, flavones and isoflavones by japanese women and the inverse correlation between quercetin intake and plasma LDL cholesterol concentration. J. Nutr. 2000, 130, 2243–2250. [Google Scholar]
- Hanneken, A.; Lin, F.-F.; Johnson, J.; Maher, P. Flavonoids protect human retinal pigment epithelial cells from oxidative-stress–induced death. Invest. Ophthalmol. Vis. Sci. 2006, 47, 3164–3177. [Google Scholar] [CrossRef]
- Higa, S.; Hirano, T.; Kotani, M.; Matsumoto, M.; Fujita, A.; Suemura, M.; Kawase, I.; Tanaka, T. Fisetin, a flavonol, inhibits TH2-type cytokine production by activated human basophils. J. Allergy Clin. Immunol. 2003, 111, 1299–1306. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Shen, S.-C.; Lee, W.-R.; Lin, H.-Y.; Ko, C.-H.; Shih, C.-M.; Yang, L.-L. Wogonin and fisetin induction of apoptosis through activation of easpase 3 cascade and alternative expression of p21 protein in hepatocellular carcinoma cells SK-HEP-1. Arch. Toxicol. 2002, 76, 351–359. [Google Scholar] [CrossRef]
- Khan, N.; Afaq, F.; Syed, D.N.; Mukhtar, H. Fisetin, a novel dietary flavonoid, causes Apoptosis and cell cycle arrest in human prostate cancer LNCaP cells. Carcinogenesis 2008, 29, 1049–1056. [Google Scholar] [CrossRef]
- Chen, F.-J.; Yang, Y.-G.; Zhao, D.-X.; Gui, Y.-L.; Guo, Z.-C. Advances in studies of species, habitats distribution and chemical composition of Snow lotuses (Saussurea) in China. Chin. Bull. Bot. 1999, 16, 561–566. [Google Scholar]
- Lee, H.-G.; Yu, K.; Oh, W.-K.; Baeg, T.-W.; Oh, H.-C.; Ahn, J.-S.; Jang, W.-C.; Kim, J.-W.; Lim, J.-S.; Choe, Y.-K. Inhibitory effect of jaceosidin isolated from Artemisiaargyi on the function of E6 and E7 oncoproteins of HPV 16. J. Ethnopharmacol. 2005, 98, 339–343. [Google Scholar] [CrossRef]
- Lee, C.-L.; Chiang, L.-C.; Cheng, L.-H.; Liaw, C.-C.; Abd El-Razek, M.H.; Chang, F.-R.; Wu, Y.-C. Influenza A (H1N1) antiviral and cytotoxic agents from Ferula assa-foetida. J. Nat. Prod. 2009, 72, 1568–1572. [Google Scholar] [CrossRef]
- Veloso, J.; Prego, C.; Varela, M.; Carballeira, R.; Bernal, A.; Merino, F.; Díaz, J. Properties of capsaicinoids for the control of fungi and oomycetes pathogenic to pepper. Plant Biol. 2013. [Google Scholar] [CrossRef]
- De Beer, E.J.; Sherwood, M.B. The paper-disc agar-plate method for the assay of antibiotic substances. J. Bacteriol. 1945, 50, 459–467. [Google Scholar]
- Smith, J.L.; de Moraes, C.M.; Mescher, M.C. Jasmonate-and salicylate-mediated plant defense responses to insect herbivores, pathogens and parasitic plants. Pest Manag. Sci. 2009, 65, 497–503. [Google Scholar] [CrossRef]
- Van Wees, S.C.; de Swart, E.A.; van Pelt, J.A.; van Loon, L.C.; Pieterse, C.M. Enhancement of induced disease resistance by simultaneous activation of salicylate-and jasmonate-dependent defense pathways in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2000, 97, 8711–8716. [Google Scholar]
- Spoel, S.H.; Johnson, J.S.; Dong, X. Regulation of tradeoffs between plant defenses against pathogens with different lifestyles. Proc. Natl. Acad. Sci. USA 2007, 104, 18842–18847. [Google Scholar] [CrossRef]
- Von Rad, U.; Mueller, M.J.; Durner, J. Evaluation of natural and synthetic stimulants of plant immunity by microarray technology. New Phytol. 2005, 165, 191–202. [Google Scholar] [CrossRef]
- Kunkel, B.N.; Brooks, D.M. Cross talk between signaling pathways in pathogen defense. Curr. Opin. Plant Biol. 2002, 5, 325–331. [Google Scholar] [CrossRef]
- Tsuda, K.; Sato, M.; Stoddard, T.; Glazebrook, J.; Katagiri, F. Network properties of robust immunity in plants. PLoS Genet. 2009, 5, e1000772. [Google Scholar] [CrossRef]
- Mengiste, T. Plant Immunity to necrotrophs. Annu. Rev. Phytopathol. 2012, 50, 267–294. [Google Scholar] [CrossRef]
- Gruner, K.; Griebel, T.; Návarová, H.; Attaran, E.; Zeier, J. Reprogramming of plants during systemic acquired resistance. Front. Plant Sci. 2013, 4, 252. [Google Scholar]
- Cichewicz, R.H.; Thorpe, P.A. The antimicrobial properties of chile peppers (Capsicum species) and their uses in mayan medicine. J. Ethnopharmacol. 1996, 52, 61–70. [Google Scholar] [CrossRef]
- Molina-Torres, J.; Garcı́a-Chávez, A.; Ramı́rez-Chávez, E. Antimicrobial properties of alkamides present in flavouring plants traditionally used in Mesoamerica: Affinin and capsaicin. J. Ethnopharmacol. 1999, 64, 241–248. [Google Scholar] [CrossRef]
- Kurita, S.; Kitagawa, E.; Kim, C.-H.; Momose, Y.; Iwahashi, H. Studies on the antimicrobial mechanisms of capsaicin using yeast DNA microarray. Biosci. Biotechnol. Biochem. 2002, 66, 532–536. [Google Scholar] [CrossRef]
- Xing, F.; Cheng, G.; Yi, K. Study on the antimicrobial activities of the capsaicin microcapsules. J. Appl. Polym. Sci. 2006, 102, 1318–1321. [Google Scholar] [CrossRef]
- Kraikruan, W.; Sangchote, S.; Sukprakarn, S. Effect of capsaicin on germination of Colletotrichum capsici conidia. Kasetsart J. Nat. Sci. 2008, 42, 417–422. [Google Scholar]
- Mauch, F.; Mauch-Mani, B.; Boller, T. Antifungal hydrolases in pea tissue. Plant Physiol. 1988, 88, 936–942. [Google Scholar] [CrossRef]
- Spencer, M.; Ryu, C.-M.; Yang, K.-Y.; Kim, Y.C.; Kloepper, J.W.; Anderson, A.J. Induced defence in tobacco by Pseudomonas chlororaphis strain O6 involves at least the ethylene pathway. Physiol. Mol. Plant Pathol. 2003, 63, 27–34. [Google Scholar] [CrossRef]
- Lee, B.; Farag, M.A.; Park, H.B.; Kloepper, J.W.; Lee, S.H.; Ryu, C.M. Induced resistance by a long-chain bacterial volatile: Elicitation of plant systemic defense by a C13 volatile produced by Paenibacillus polymyxa. PLoS One 2012, 7, e48744. [Google Scholar]
- Choi, J.-S.; Park, N.-J.; Lim, H.-K.; Ko, Y.-K.; Kim, Y.-S.; Ryu, S.-Y.; Hwang, I.-T. Plumbagin as a new natural herbicide candidate for Sicyon angulatus control agent with the target 8-amino-7-oxononanoate synthase. Pestic. Biochem. Physiol. 2012, 103, 166–172. [Google Scholar] [CrossRef]
- Bhattacharyya, J.; Carvalho, V. R. de. Epi-isoshinanolone from Plumbago scandens. Phytochemistry 1986, 25, 764–765. [Google Scholar] [CrossRef]
- Lee, B.; Park, Y.-S.; Yi, H.-S.; and Ryu, C.-M. Systemic induction of the small antibacterial compound in the leaf exudate during benzothiadiazole-elicited systemic acquired resistance in pepper. Plant Pathol. J. 2013, 29, 350–355. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the capsaicin, fiserin hydrate, jaceosidin, and farnesiferol A are available from the Dr. Shi Yong Ryu.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Song, G.C.; Ryu, S.Y.; Kim, Y.S.; Lee, J.Y.; Choi, J.S.; Ryu, C.-M. Elicitation of Induced Resistance against Pectobacterium carotovorum and Pseudomonas syringae by Specific Individual Compounds Derived from Native Korean Plant Species. Molecules 2013, 18, 12877-12895. https://doi.org/10.3390/molecules181012877
Song GC, Ryu SY, Kim YS, Lee JY, Choi JS, Ryu C-M. Elicitation of Induced Resistance against Pectobacterium carotovorum and Pseudomonas syringae by Specific Individual Compounds Derived from Native Korean Plant Species. Molecules. 2013; 18(10):12877-12895. https://doi.org/10.3390/molecules181012877
Chicago/Turabian StyleSong, Geun Cheol, Shi Yong Ryu, Young Sup Kim, Ji Young Lee, Jung Sup Choi, and Choong-Min Ryu. 2013. "Elicitation of Induced Resistance against Pectobacterium carotovorum and Pseudomonas syringae by Specific Individual Compounds Derived from Native Korean Plant Species" Molecules 18, no. 10: 12877-12895. https://doi.org/10.3390/molecules181012877