OSBP-Related Proteins: Liganding by Glycerophospholipids Opens New Insight into Their Function

Abstract
:1. Introduction
2. OSBP-Related Proteins and Their Liganding by Sterols

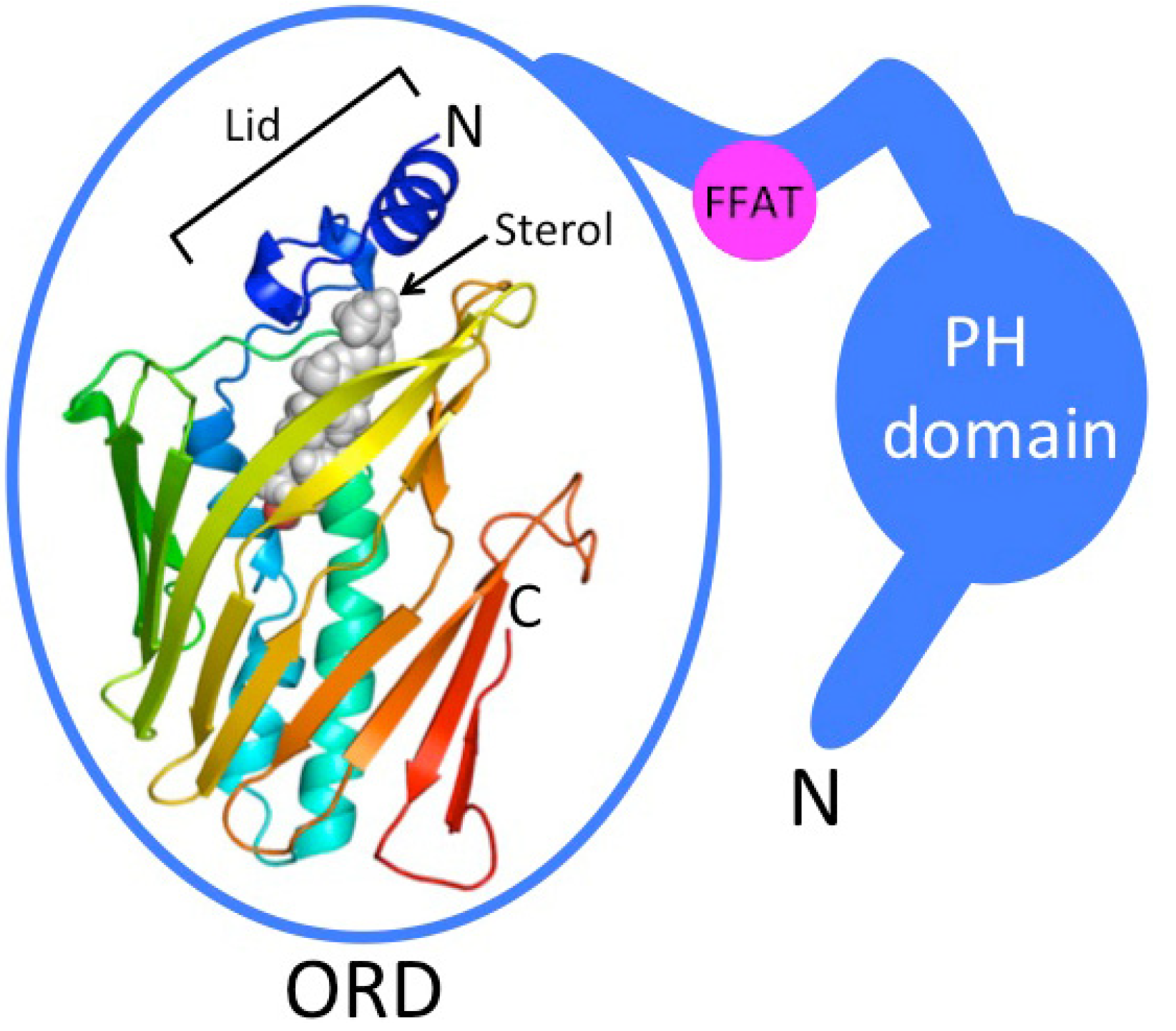
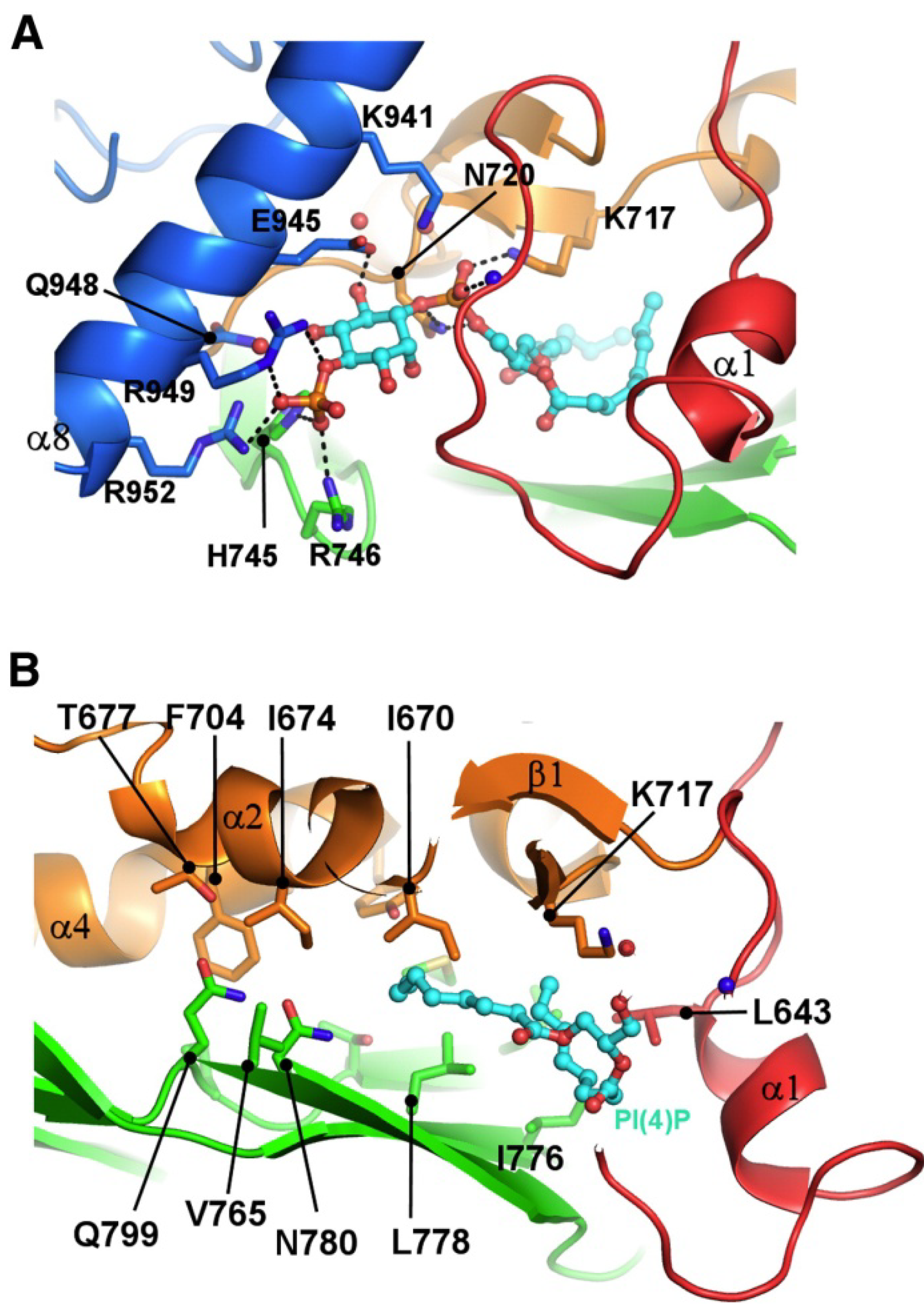
3. ORPs Can Bind Glycerophospholipids


{kind=link}
{kind=link}
{kind=link}
| Protein | ORD ligand(s) | Reference(s) |
|---|---|---|
| Homo sapiens | ||
| OSBP | 25OHC 1, 20OHC, 7KC 2, 22(R)OHC, 22(S)OHC, 7OHC, cholesterol + other oxysterols specified in [42] | [40,42,43,44] |
| Saccharomyces cerevisiae | ||
| Osh1p | Cholesterol? | [38] |
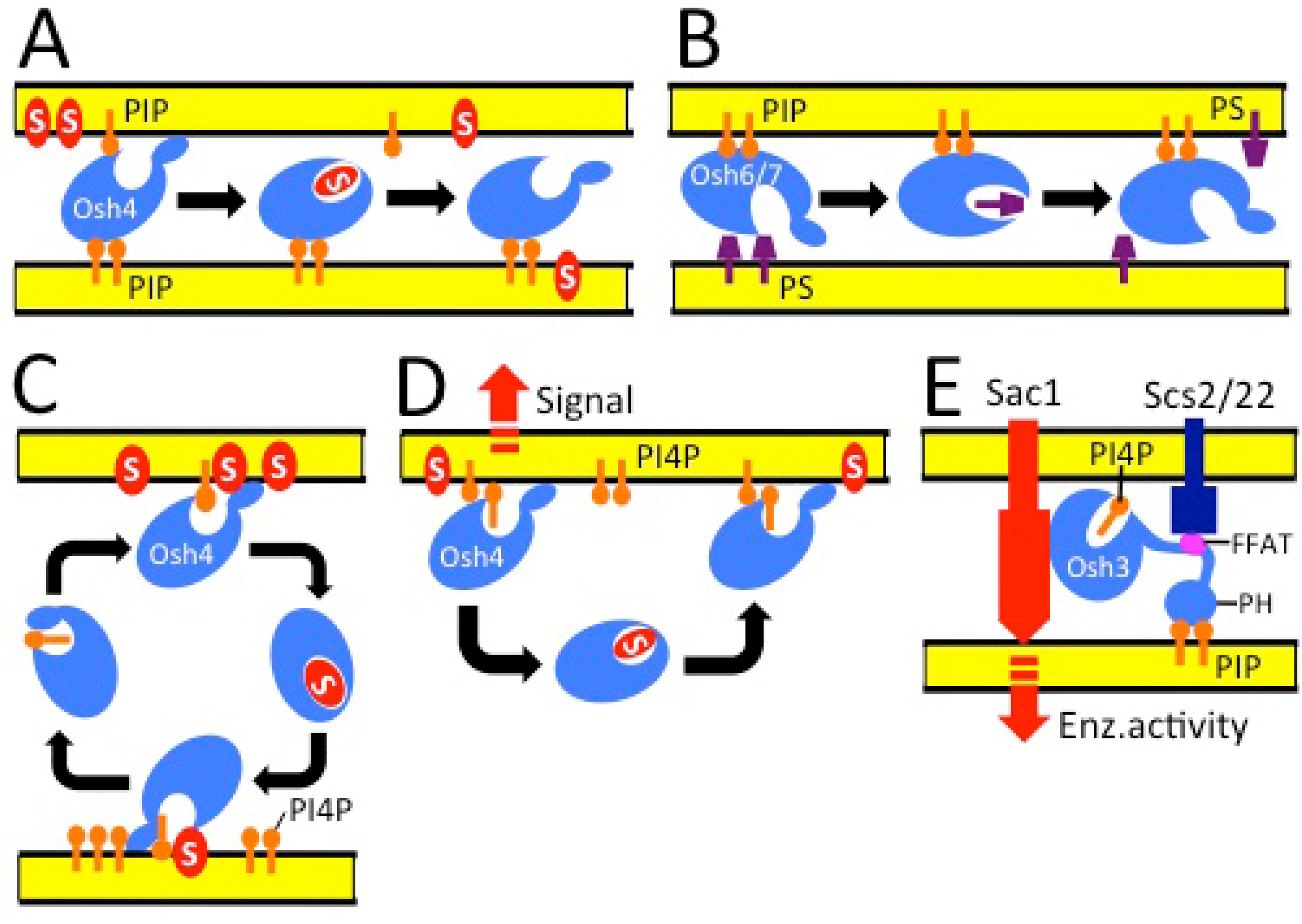
4. Function of ORPs at Membrane Contact Sites

5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell. Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Poulsen, L.R.; Lopez-Marques, R.L.; Palmgren, M.G. Flippases: Still more questions than answers. Cell. Mol. Life Sci. 2008, 65, 3119–3125. [Google Scholar]
- Sprong, H.; van der Sluijs, P.; van Meer, G. How proteins move lipids and lipids move proteins. Nat. Rev. Mol. Cell. Biol. 2001, 2, 504–513. [Google Scholar]
- Gillon, A.D.; Latham, C.F.; Miller, E.A. Vesicle-mediated ER export of proteins and lipids. Biochim Biophys Acta 2012, 1821, 1040–1049. [Google Scholar]
- Van Meer, G.; Sprong, H. Membrane lipids and vesicular traffic. Curr. Opin. Cell. Biol. 2004, 16, 373–378. [Google Scholar] [CrossRef]
- D’Angelo, G.; Vicinanza, M.; de Matteis, M.A. Lipid-transfer proteins in biosynthetic pathways. Curr. Opin. Cell. Biol. 2008, 20, 360–370. [Google Scholar] [CrossRef]
- Lev, S. Non-vesicular lipid transport by lipid-transfer proteins and beyond. Nat. Rev. Mol. Cell. Biol. 2010, 11, 739–750. [Google Scholar] [CrossRef]
- Elbaz, Y.; Schuldiner, M. Staying in touch: The molecular era of organelle contact sites. Trends Biochem. Sci. 2011, 36, 616–623. [Google Scholar] [CrossRef]
- Levine, T.; Loewen, C. Inter-organelle membrane contact sites: through a glass, darkly. Curr. Opin. Cell. Biol. 2006, 18, 371–378. [Google Scholar] [CrossRef]
- Stefan, C.J.; Manford, A.G.; Emr, S.D. ER-PM connections: Sites of information transfer and inter-organelle communication. Curr. Opin. Cell. Biol. 2013, 25, 434–442. [Google Scholar] [CrossRef]
- Toulmay, A.; Prinz, W.A. Lipid transfer and signaling at organelle contact sites: The tip of the iceberg. Curr. Opin. Cell. Biol. 2011, 23, 458–463. [Google Scholar] [CrossRef]
- Maxfield, F.R.; van Meer, G. Cholesterol, the central lipid of mammalian cells. Curr. Opin. Cell. Biol. 2010, 22, 422–429. [Google Scholar] [CrossRef]
- Ikonen, E. Cellular cholesterol trafficking and compartmentalization. Nat. Rev. Mol. Cell. Biol. 2008, 9, 125–138. [Google Scholar] [CrossRef]
- Mesmin, B.; Maxfield, F.R. Intracellular sterol dynamics. Biochim. Biophys. Acta 2009, 1791, 636–645. [Google Scholar] [CrossRef]
- Voelker, D.R. Bridging gaps in phospholipid transport. Trends Biochem Sci 2005, 30, 396–404. [Google Scholar] [CrossRef]
- Voelker, D.R. Genetic and biochemical analysis of non-vesicular lipid traffic. Annu. Rev. Biochem. 2009, 78, 827–856. [Google Scholar] [CrossRef]
- Björkhem, I.; Diczfalusy, U. Oxysterols: Friends, foes, or just fellow passengers? Arterioscle. Thromb. Vasc. Biol. 2002, 22, 734–742. [Google Scholar] [CrossRef]
- Russell, D.W. Oxysterol biosynthetic enzymes. Biochim. Biophys. Acta 2000, 1529, 126–135. [Google Scholar]
- Brown, A.J.; Jessup, W. Oxysterols and atherosclerosis. Atherosclerosis 1999, 142, 1–28. [Google Scholar] [CrossRef]
- Olkkonen, V.M.; Béaslas, O.; Nissilä, E. Oxysterols and their cellular effectors. Biomolecules 2012, 2, 76–103. [Google Scholar] [CrossRef]
- Lange, Y.; Ye, J.; Strebel, F. Movement of 25-hydroxycholesterol from the plasma membrane to the rough endoplasmic reticulum in cultured hepatoma cells. J. Lipid Res. 1995, 36, 1092–1097. [Google Scholar]
- Massey, J.B. Membrane and protein interactions of oxysterols. Curr. Opin. Lipidol. 2006, 17, 296–301. [Google Scholar] [CrossRef]
- Massey, J.B.; Pownall, H.J. Structures of biologically active oxysterols determine their differential effects on phospholipid membranes. Biochemistry 2006, 45, 10747–10758. [Google Scholar] [CrossRef]
- Olkkonen, V.M.; Hynynen, R. Interactions of oxysterols with membranes and proteins. Mol. Aspects Med. 2009, 30, 123–133. [Google Scholar] [CrossRef]
- Lordan, S.; Mackrill, J.J.; O’Brien, N.M. Oxysterols and mechanisms of apoptotic signaling: Implications in the pathology of degenerative diseases. J. Nutr. Biochem. 2009, 20, 321–336. [Google Scholar] [CrossRef]
- Olkkonen, V.M. Macrophage oxysterols and their binding proteins: Roles in atherosclerosis. Curr. Opin. Lipidol. 2012, 23, 462–470. [Google Scholar] [CrossRef]
- Shibata, N.; Glass, C.K. Macrophages, oxysterols and atherosclerosis. Circ. J. 2010, 74, 2045–2051. [Google Scholar] [CrossRef]
- Vejux, A.; Malvitte, L.; Lizard, G. Side effects of oxysterols: Cytotoxicity, oxidation, inflammation, and phospholipidosis. Braz J. Med. Biol. Res. 2008, 41, 545–556. [Google Scholar] [CrossRef]
- Wang, D.Q.; Afdhal, N.H. Good cholesterol, bad cholesterol: Role of oxysterols in biliary tract diseases. Gastroenterology 2001, 121, 216–218. [Google Scholar] [CrossRef]
- Javitt, N.B. Oxysterols: Novel biologic roles for the 21st century. Steroids 2008, 73, 149–157. [Google Scholar] [CrossRef]
- Anniss, A.M.; Apostolopoulos, J.; Dworkin, S.; Purton, L.E.; Sparrow, R.L. An oxysterol-binding protein family identified in the mouse. DNA Cell. Biol. 2002, 21, 571–580. [Google Scholar] [CrossRef]
- Beh, C.T.; Cool, L.; Phillips, J.; Rine, J. Overlapping functions of the yeast oxysterol-binding protein homologues. Genetics 2001, 157, 1117–1140. [Google Scholar]
- Im, Y.J.; Raychaudhuri, S.; Prinz, W.A.; Hurley, J.H. Structural mechanism for sterol sensing and transport by OSBP-related proteins. Nature 2005, 437, 154–158. [Google Scholar]
- Jaworski, C.J.; Moreira, E.; Li, A.; Lee, R.; Rodriguez, I.R. A family of 12 human genes containing oxysterol-binding domains. Genomics 2001, 78, 185–196. [Google Scholar] [CrossRef]
- Lehto, M.; Laitinen, S.; Chinetti, G.; Johansson, M.; Ehnholm, C.; Staels, B.; Ikonen, E.; Olkkonen, V.M. The OSBP-related protein family in humans. J. Lipid Res. 2001, 42, 1203–1213. [Google Scholar]
- Olkkonen, V.M.; Li, S. Oxysterol-binding proteins: Sterol and phosphoinositide sensors coordinating transport, signaling and metabolism. Prog. Lipid Res. 2013, 52, 529–538. [Google Scholar] [CrossRef]
- Raychaudhuri, S.; Im, Y.J.; Hurley, J.H.; Prinz, W.A. Nonvesicular sterol movement from plasma membrane to ER requires oxysterol-binding protein-related proteins and phosphoinositides. J. Cell. Biol. 2006, 173, 107–119. [Google Scholar] [CrossRef]
- Schulz, T.A.; Choi, M.G.; Raychaudhuri, S.; Mears, J.A.; Ghirlando, R.; Hinshaw, J.E.; Prinz, W.A. Lipid-regulated sterol transfer between closely apposed membranes by oxysterol-binding protein homologues. J. Cell. Biol. 2009, 187, 889–903. [Google Scholar] [CrossRef]
- Georgiev, A.G.; Sullivan, D.P.; Kersting, M.C.; Dittman, J.S.; Beh, C.T.; Menon, A.K. Osh proteins regulate membrane sterol organization but are not required for sterol movement between the ER and PM. Traffic 2011, 12, 1341–1355. [Google Scholar] [CrossRef]
- Dawson, P.A.; Ridgway, N.D.; Slaughter, C.A.; Brown, M.S.; Goldstein, J.L. cDNA cloning and expression of oxysterol-binding protein, an oligomer with a potential leucine zipper. J. Biol. Chem. 1989, 264, 16798–16803. [Google Scholar]
- Ridgway, N.D.; Dawson, P.A.; Ho, Y.K.; Brown, M.S.; Goldstein, J.L. Translocation of oxysterol binding protein to Golgi apparatus triggered by ligand binding. J. Cell. Biol. 1992, 116, 307–319. [Google Scholar] [CrossRef]
- Taylor, F.R.; Saucier, S.E.; Shown, E.P.; Parish, E.J.; Kandutsch, A.A. Correlation between oxysterol binding to a cytosolic binding protein and potency in the repression of hydroxymethylglutaryl coenzyme A reductase. J. Biol. Chem. 1984, 259, 12382–12387. [Google Scholar]
- Wang, C.; JeBailey, L.; Ridgway, N.D. Oxysterol-binding-protein (OSBP)-related protein 4 binds 25-hydroxycholesterol and interacts with vimentin intermediate filaments. Biochem. J. 2002, 361, 461–472. [Google Scholar] [CrossRef]
- Wang, P.Y.; Weng, J.; Lee, S.; Anderson, R.G. N-terminus controls sterol binding while c-terminus regulates scaffolding function of OSBP. J. Biol. Chem. 2007, 283, 8034–8045. [Google Scholar]
- Hynynen, R.; Suchanek, M.; Spandl, J.; Back, N.; Thiele, C.; Olkkonen, V.M. OSBP-related protein 2 is a sterol receptor on lipid droplets that regulates the metabolism of neutral lipids. J. Lipid Res. 2009, 50, 1305–1315. [Google Scholar] [CrossRef]
- Suchanek, M.; Hynynen, R.; Wohlfahrt, G.; Lehto, M.; Johansson, M.; Saarinen, H.; Radzikowska, A.; Thiele, C.; Olkkonen, V.M. The mammalian OSBP-related proteins (ORP) bind 25-hydroxycholesterol in an evolutionarily conserved pocket. Biochem. J. 2007, 405, 473–480. [Google Scholar] [CrossRef]
- Nissilä, E.; Ohsaki, Y.; Weber-Boyvat, M.; Perttilä, J.; Ikonen, E.; Olkkonen, V.M. ORP10, a cholesterol binding protein associated with microtubules, regulates apolipoprotein B-100 secretion. Biochim. Biophys. Acta 2012, 1821, 1472–1484. [Google Scholar] [CrossRef]
- Vihervaara, T.; Uronen, R.L.; Wohlfahrt, G.; Björkhem, I.; Ikonen, E.; Olkkonen, V.M. Sterol binding by OSBP-related protein 1L regulates late endosome motility and function. Cell. Mol. Life Sci. 2011, 68, 537–551. [Google Scholar] [CrossRef]
- Du, X.; Kumar, J.; Ferguson, C.; Schulz, T.A.; Ong, Y.S.; Hong, W.; Prinz, W.A.; Parton, R.G.; Brown, A.J.; Yang, H. A role for oxysterol-binding protein-related protein 5 in endosomal cholesterol trafficking. J. Cell. Biol. 2011, 192, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Ngo, M.; Ridgway, N.D. Oxysterol binding protein-related Protein 9 (ORP9) is a cholesterol transfer protein that regulates Golgi structure and function. Mol. Biol. Cell. 2009, 20, 1388–1399. [Google Scholar] [CrossRef]
- De Saint-Jean, M.; Delfosse, V.; Douguet, D.; Chicanne, G.; Payrastre, B.; Bourguet, W.; Antonny, B.; Drin, G. Osh4p exchanges sterols for phosphatidylinositol 4-phosphate between lipid bilayers. J. Cell. Biol. 2011, 195, 965–978. [Google Scholar] [CrossRef]
- Mesmin, B.; Antonny, B.; Drin, G. Insights into the mechanisms of sterol transport between organelles. Cell. Mol. Life Sci. 2013, 70, 3405–3421. [Google Scholar] [CrossRef]
- Fang, M.; Kearns, B.G.; Gedvilaite, A.; Kagiwada, S.; Kearns, M.; Fung, M.K.; Bankaitis, V.A. Kes1p shares homology with human oxysterol binding protein and participates in a novel regulatory pathway for yeast Golgi-derived transport vesicle biogenesis. Embo. J. 1996, 15, 6447–6459. [Google Scholar]
- Li, X.; Rivas, M.P.; Fang, M.; Marchena, J.; Mehrotra, B.; Chaudhary, A.; Feng, L.; Prestwich, G.D.; Bankaitis, V.A. Analysis of oxysterol binding protein homologue Kes1p function in regulation of Sec14p-dependent protein transport from the yeast Golgi complex. J. Cell. Biol. 2002, 157, 63–77. [Google Scholar] [CrossRef]
- Mousley, C.J.; Yuan, P.; Gaur, N.A.; Trettin, K.D.; Nile, A.H.; Deminoff, S.J.; Dewar, B.J.; Wolpert, M.; Macdonald, J.M.; Herman, P.K.; et al. A Sterol-Binding Protein Integrates Endosomal Lipid Metabolism with TOR Signaling and Nitrogen Sensing. Cell 2012, 148, 702–715. [Google Scholar] [CrossRef]
- Alfaro, G.; Johansen, J.; Dighe, S.A.; Duamel, G.; Kozminski, K.G.; Beh, C.T. The sterol-binding protein Kes1/Osh4p is a regulator of polarized exocytosis. Traffic 2011, 12, 1521–1536. [Google Scholar] [CrossRef]
- Leblanc, M.A.; Fairn, G.D.; Russo, S.B.; Czyz, O.; Zaremberg, V.; Cowart, L.A.; McMaster, C.R. The yeast oxysterol binding protein kes1 maintains sphingolipid levels. PLoS One 2013, 8, e60485. [Google Scholar]
- Tong, J.; Yang, H.; Eom, S.H.; Im, Y.J. Structure of osh3 reveals a conserved mode of phosphoinositide binding in oxysterol-binding proteins. Structure 2013, 21, 1203–1213. [Google Scholar] [CrossRef]
- Maeda, K.; Anand, K.; Chiapparino, A.; Kumar, A.; Poletto, M.; Kaksonen, M.; Gavin, A.C. Interactome map uncovers phosphatidylserine transport by oxysterol-binding proteins. Nature 2013. [Google Scholar] [CrossRef]
- Wang, P.; Duan, W.; Munn, A.L.; Yang, H. Molecular characterization of Osh6p, an oxysterol binding protein homolog in the yeast Saccharomyces cerevisiae. FEBS J. 2005, 272, 4703–4715. [Google Scholar] [CrossRef]
- Wyles, J.P.; Perry, R.J.; Ridgway, N.D. Characterization of the sterol-binding domain of oxysterol-binding protein (OSBP)-related protein 4 reveals a novel role in vimentin organization. Exp. Cell. Res. 2007, 313, 1426–1437. [Google Scholar] [CrossRef]
- Yan, D.; Mäyränpää, M.I.; Wong, J.; Perttilä, J.; Lehto, M.; Jauhiainen, M.; Kovanen, P.T.; Ehnholm, C.; Brown, A.J.; Olkkonen, V.M. OSBP-related protein 8 (ORP8) suppresses ABCA1 expression and cholesterol efflux from macrophages. J. Biol. Chem. 2008, 283, 332–340. [Google Scholar]
- Zhou, T.; Li, S.; Zhong, W.; Vihervaara, T.; Béaslas, O.; Perttilä, J.; Luo, W.; Jiang, Y.; Lehto, M.; Olkkonen, V.M.; Yan, D. OSBP-Related Protein 8 (ORP8) Regulates Plasma and Liver Tissue Lipid Levels and Interacts with the Nucleoporin Nup62. PLoS One 2011, 6, e21078. [Google Scholar] [CrossRef]
- Kvam, E.; Goldfarb, D.S. Nvj1p is the outer-nuclear-membrane receptor for oxysterol-binding protein homolog Osh1p in Saccharomyces cerevisiae. J. Cell. Sci. 2004, 117, 4959–4968. [Google Scholar] [CrossRef]
- Levine, T.P.; Munro, S. Dual targeting of Osh1p, a yeast homologue of oxysterol-binding protein, to both the Golgi and the nucleus-vacuole junction. Mol. Biol. Cell. 2001, 12, 1633–1644. [Google Scholar] [CrossRef]
- Olkkonen, V.M.; Levine, T.P. Oxysterol binding proteins: in more than one place at one time? Biochem. Cell Biol. 2004, 82, 87–98. [Google Scholar] [CrossRef]
- Stefan, C.J.; Manford, A.G.; Baird, D.; Yamada-Hanff, J.; Mao, Y.; Emr, S.D. Osh proteins regulate phosphoinositide metabolism at ER-plasma membrane contact sites. Cell 2011, 144, 389–401. [Google Scholar] [CrossRef]
- Pichler, H.; Gaigg, B.; Hrastnik, C.; Achleitner, G.; Kohlwein, S.D.; Zellnig, G.; Perktold, A.; Daum, G. A subfraction of the yeast endoplasmic reticulum associates with the plasma membrane and has a high capacity to synthesize lipids. Eur. J. Biochem. 2001, 268, 2351–2361. [Google Scholar] [CrossRef]
- Hanada, K.; Kumagai, K.; Tomishige, N.; Yamaji, T. CERT-mediated trafficking of ceramide. Biochim. Biophys. Acta 2009, 1791, 684–691. [Google Scholar]
- Johansson, M.; Lehto, M.; Tanhuanpää, K.; Cover, T.L.; Olkkonen, V.M. The oxysterol-binding protein homologue ORP1L interacts with Rab7 and alters functional properties of late endocytic compartments. Mol. Biol. Cell. 2005, 16, 5480–5492. [Google Scholar] [CrossRef]
- Rocha, N.; Kuijl, C.; van der Kant, R.; Janssen, L.; Houben, D.; Janssen, H.; Zwart, W.; Neefjes, J. Cholesterol sensor ORP1L contacts the ER protein VAP to control Rab7-RILP-p150 Glued and late endosome positioning. J. Cell. Biol. 2009, 185, 1209–1225. [Google Scholar] [CrossRef]
- Johansson, M.; Rocha, N.; Zwart, W.; Jordens, I.; Janssen, L.; Kuijl, C.; Olkkonen, V.M.; Neefjes, J. Activation of endosomal dynein motors by stepwise assembly of Rab7-RILP-p150Glued, ORP1L, and the receptor betalll spectrin. J. Cell. Biol. 2007, 176, 459–471. [Google Scholar] [CrossRef]
- Van der Kant, R.; Fish, A.; Janssen, L.; Janssen, H.; Krom, S.; Ho, N.; Brummelkamp, T.; Carette, J.; Rocha, N.; Neefjes, J. Late endosomal transport and tethering are coupled processes controlled by RILP and the cholesterol sensor ORP1L. J. Cell. Sci. 2013, 126, 3462–3474. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Olkkonen, V.M. OSBP-Related Proteins: Liganding by Glycerophospholipids Opens New Insight into Their Function. Molecules 2013, 18, 13666-13679. https://doi.org/10.3390/molecules181113666
Olkkonen VM. OSBP-Related Proteins: Liganding by Glycerophospholipids Opens New Insight into Their Function. Molecules. 2013; 18(11):13666-13679. https://doi.org/10.3390/molecules181113666
Chicago/Turabian StyleOlkkonen, Vesa M. 2013. "OSBP-Related Proteins: Liganding by Glycerophospholipids Opens New Insight into Their Function" Molecules 18, no. 11: 13666-13679. https://doi.org/10.3390/molecules181113666