Discrimination of Multi-Origin Chinese Herbal Medicines Using Gas Chromatography-Mass Spectrometry-Based Fatty Acid Profiling

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Validation of the GC-MS Method

{kind=link}
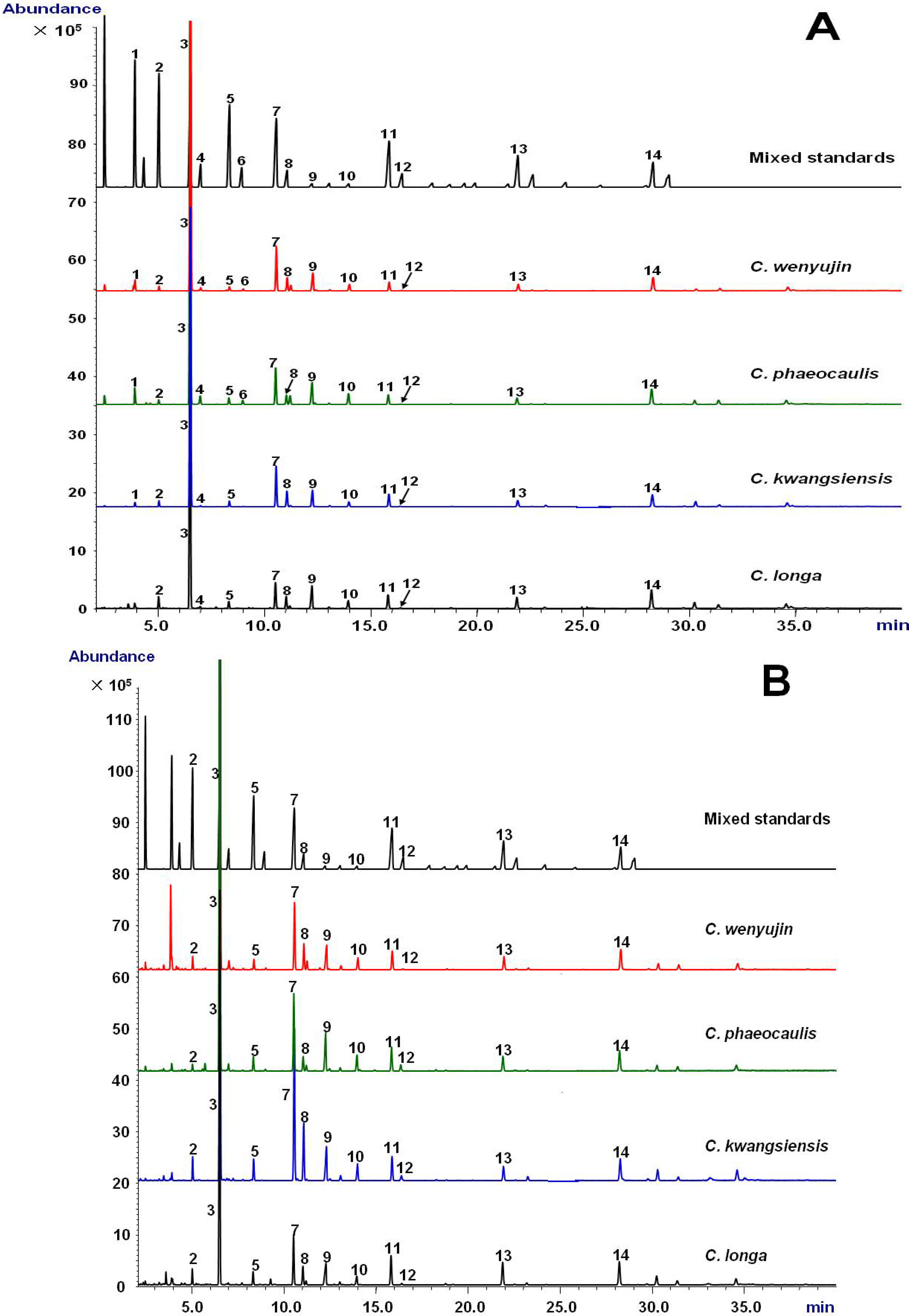{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FA | Precision | Stability | Repeatability | |||
|---|---|---|---|---|---|---|
| Content (%) | RSD (%) | Content (%) | RSD (%) | Content (%) | RSD (%) | |
| C15:0 | 9.16 ± 0.22 | 2.39 | 0.57 ± 0.02 | 2.95 | 0.57 ± 0.03 | 4.49 |
| C16:0 | 9.69 ± 0.15 | 1.52 | 25.01 ± 0.21 | 0.84 | 25.53 ± 0.32 | 1.27 |
| C17:0 | 9.85 ± 0.07 | 0.68 | 0.61 ± 0.01 | 1.95 | 0.58 ± 0.03 | 5.15 |
| C18:0 | 9.97 ± 0.05 | 0.46 | 4.23 ± 0.06 | 1.52 | 4.45 ± 0.39 | 8.77 |
| C18:1 n-9 | 10.14 ± 0.02 | 0.23 | 7.59 ± 0.08 | 1.07 | 7.71 ± 0.12 | 1.58 |
| C18:2 n-6 | 9.74 ± 0.04 | 0.43 | 39.39 ± 0.31 | 0.77 | 38.71 ± 0.46 | 1.18 |
| C18:3 n-3 | 9.46 ± 0.02 | 0.26 | 17.3 ± 0.59 | 3.4 | 17.03 ± 0.35 | 2.08 |
| C20:0 | 8.71 ± 0.08 | 0.86 | 1.54 ± 0.05 | 3.11 | 1.58 ± 0.08 | 4.92 |
| C20:1 | 10.87 ± 0.10 | 0.96 | 0.55 ± 0.02 | 2.77 | 0.52 ± 0.05 | 8.90 |
| C22:0 | 6.74 ± 0.22 | 3.24 | 1.17 ± 0.07 | 6.01 | 1.21 ± 0.04 | 3.7 |
| C24:0 | 5.66 ± 0.10 | 1.69 | 2.02 ± 0.11 | 5.38 | 2.09 ± 0.10 | 4.96 |
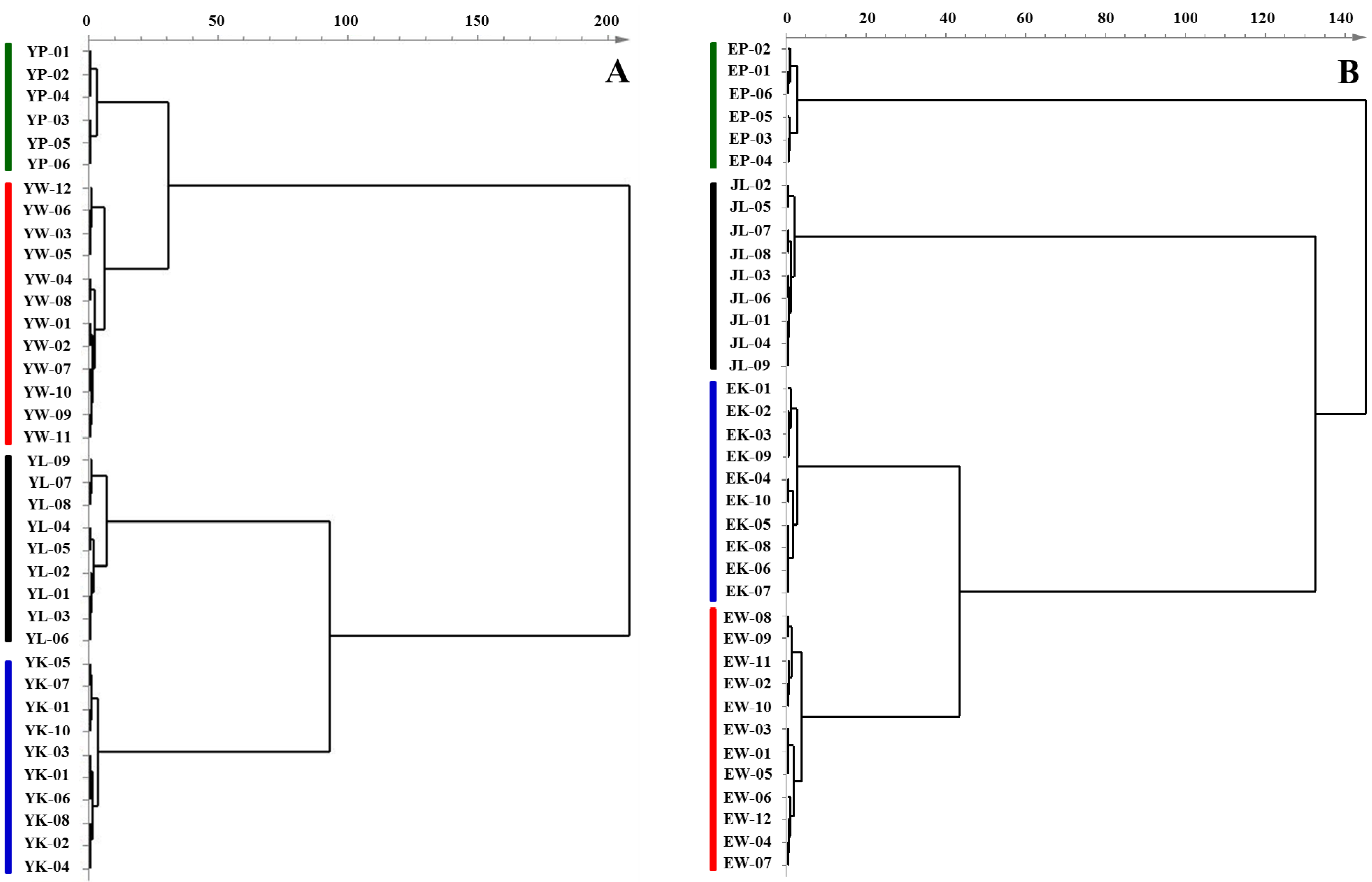
2.2. Fatty Acid Composition of Tuberous Roots and Rhizomes Derived from Four Curcuma Species
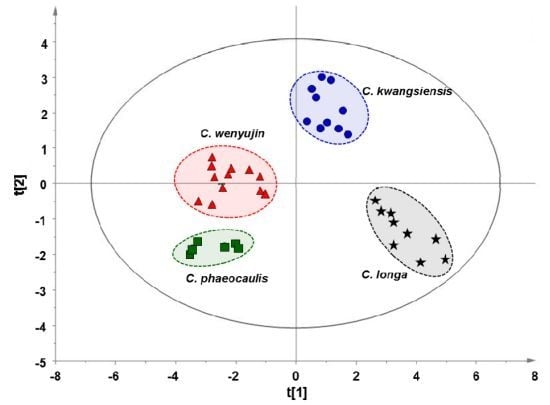
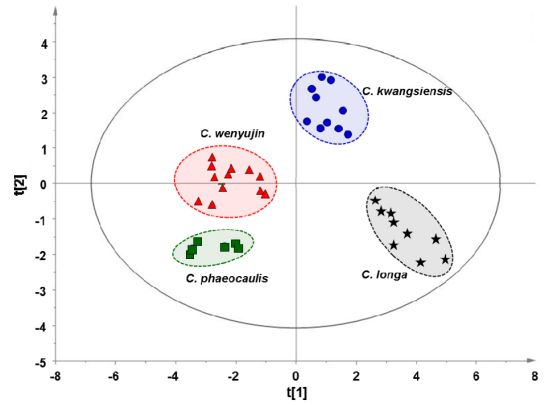
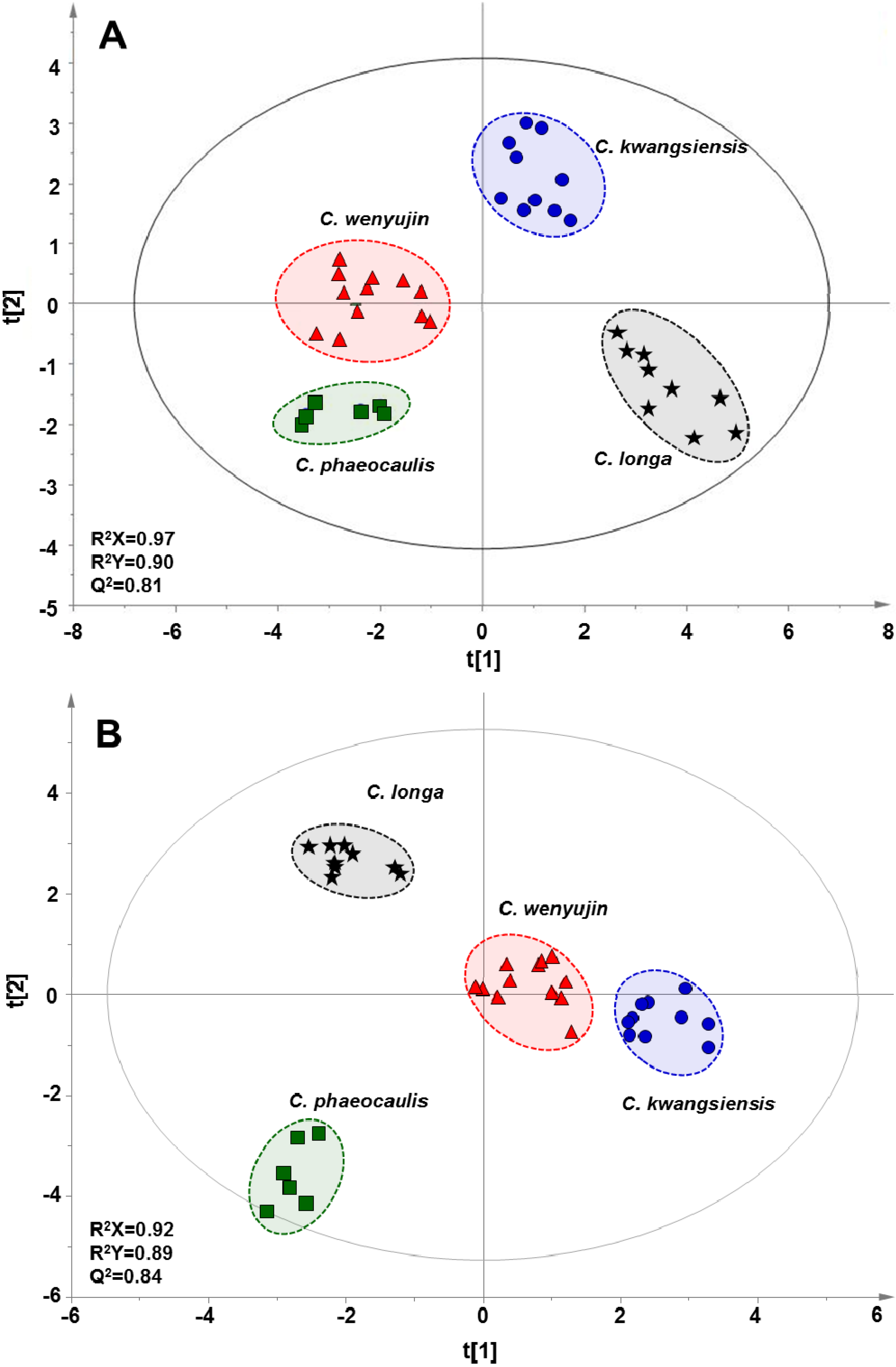
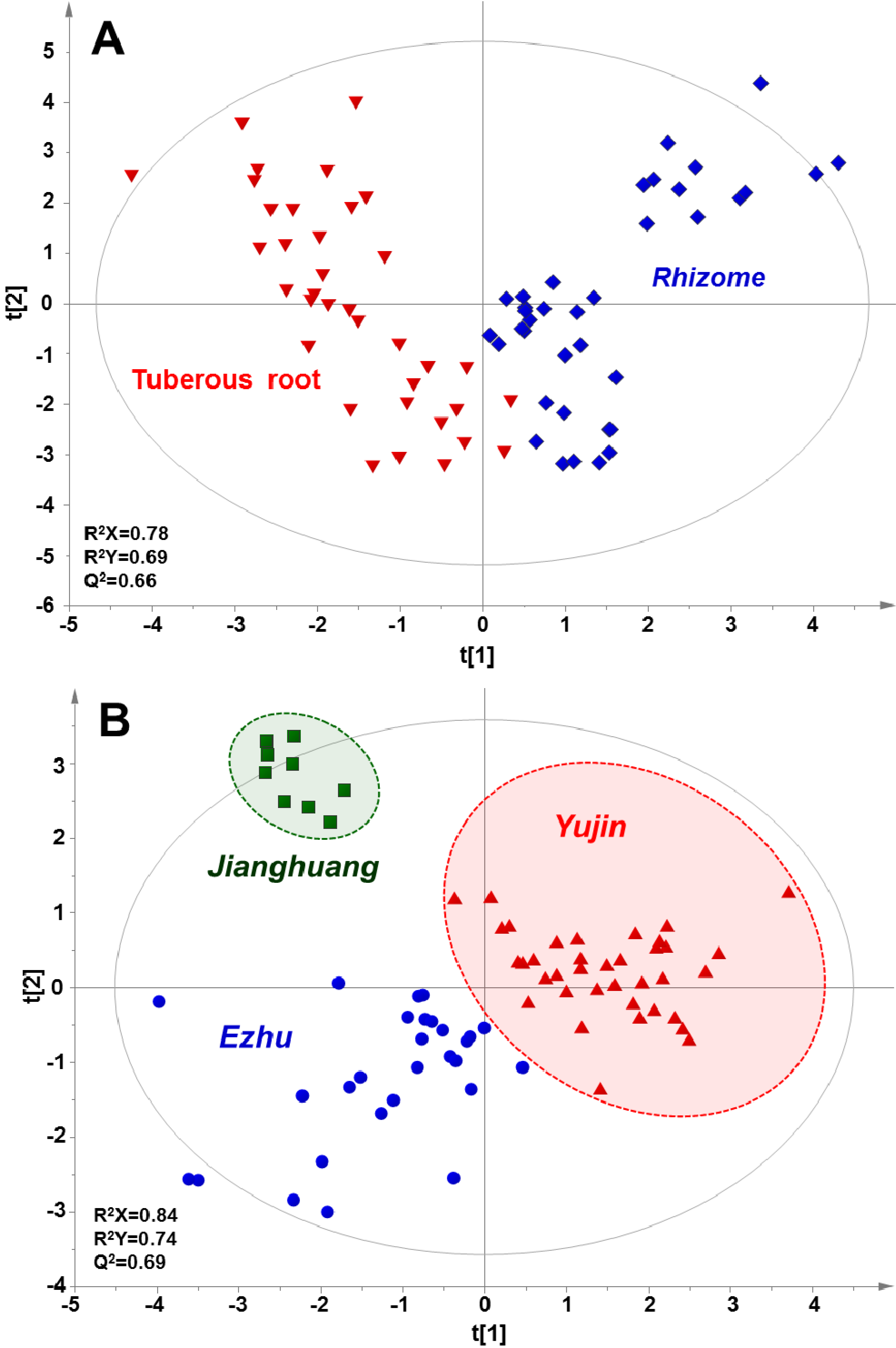
2.3. Multivariate Statistical Analysis

| Peak No. | Fatty Acid (%) | Tuberous Root | Rhizome | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Common Name | Symbol | CW ( n = 12) | CP ( n = 6) | CK ( n = 10) | CL ( n = 9) | CW ( n = 12) | CP( n = 6) | CK ( n = 10) | CL ( n = 9) | |
| 1 | Tetradecanoic acid | C14:0 | 0.66 ± 0.31 a | 0.83 ± 0.19 a | 0.32 ± 0.12 b | ND | ND | ND | ND | ND |
| 2 | Pentadecanoic acid | C15:0 | 0.32 ± 0.06 a | 0.28 ± 0.02 b | 0.46 ± 0.05 c | 0.76 ± 0.09 d | 0.56 ± 0.05 a | 0.26 ± 0.02 b | 0.68 ± 0.12 c | 0.77 ± 0.08 d |
| 3 | Palmitic acid | C16:0 | 28.32 ± 1.10 a | 29.40 ± 0.96 b | 27.09 ± 1.5 c | 24.55 ± 0.54 d | 25.45 ± 1.10 a | 25.82 ± 0.38 a | 25.55 ± 1.89 a | 22.88 ± 0.64 b |
| 4 | Palmitoleic acid | C16:1 n-7 | 2.85 ± 2.25 a | 3.62 ± 0.18 a | 0.58 ± 0.24 b | 0.53 ± 0.18 b | ND | 2.01 ± 0.35 | ND | ND |
| 5 | Heptadecanoic acid | C17:0 | 0.33 ± 0.04 a | 0.47 ± 0.04 b | 0.52 ± 0.07 b | 0.69 ± 0.11 c | 0.62 ± 0.07 a | 0.63 ± 0.05 a | 0.85 ± 0.16 b | 0.77 ± 0.05 b |
| 6 | Heptadecenoic acid | C17:1 n-7 | 0.62 ± 0.41 a | 1.55 ± 0.28 b | ND | ND | ND | 0.85 ± 0.36 | ND | ND |
| 7 | Stearic acid | C18:0 | 4.43 ± 0.88 a | 3.11 ± 0.76 b | 4.28 ± 0.59 a | 3.08 ± 0.43 b | 4.78 ± 0.58 a | 3.79 ± 0.17 b | 5.64 ± 0.53 c | 3.39 ± 0.17 d |
| 8 | Oleic acid | C18:1 n-9 | 5.49 ± 0.84 a | 3.02 ± 0.49 b | 10.78 ± 2.4 c | 4.57 ± 0.56 d | 8.92 ± 0.89 a | 4.08 ± 0.47 b | 10.83 ± 1.51 c | 6.28 ± 1.28 d |
| 9 | Linoleic acid | C18:2 n-6 | 37.29 ± 2.35 a | 35.66 ± 1.19 a | 40.10 ± 0.79 b | 42.78 ± 1.15 c | 39.73 ± 0.92 a | 38.26 ± 1.18 b | 38.61 ± 2.36 a,b | 42.61 ± 1.09 c |
| 10 | α-linolenic acid | C18:3 n-3 | 15.52 ± 0.93 a | 17.65 ± 1.0 b | 10.56 ± 2.62 c | 16.25 ± 0.71 d | 14.57 ± 1.47 a | 17.59 ± 0.65 b | 13.37 ± 2.42 a | 15.69 ± 1.20 c |
| 11 | Arachidic acid | C20:0 | 0.90 ± 0.17 a | 0.99 ± 0.21 a | 1.34 ± 0.58 a | 1.68 ± 0.23 b | 1.33 ± 0.21 a | 1.64 ± 0.21 b | 0.91 ± 0.32 c | 2.20 ± 0.37 d |
| 12 | Eicosenoic acid | C20:1 n-9 | 0.26 ± 0.06 a | 0.37 ± 0.13 a,b | 0.47 ± 0.12 b | 0.46 ± 0.06 c | 0.54 ± 0.08 a | 1.85 ± 0.08 b | 1.02 ± 0.58 c | 0.56 ± 0.10 a |
| 13 | Docosanoic acid | C22:0 | 0.81 ± 0.20 a | 0.86 ± 0.09 a,b | 1.06 ± 0.33 b | 1.63 ± 0.22 c | 1.18 ± 0.18 a | 1.34 ± 0.26 b | 0.81 ± 0.27 c | 2.14 ± 0.41 d |
| 14 | Lignoceric acid | C24:0 | 2.27 ± 0.40 a | 2.19 ± 0.21 a | 2.44 ± 0.73 a | 3.03 ± 0.36 b | 2.33 ± 0.39 a | 1.88 ± 0.67 b | 1.73 ± 0.51 b | 2.73 ± 0.46 c |
| SFA | 37.95 ± 1.36 a | 38.12 ± 1.26 a | 37.50 ± 0.81 a | 35.41 ± 0.81 b | 36.24 ± 1.34 a | 35.37 ± 0.74 | 36.18 ± 2.29 | 34.89 ± 1.12 b | ||
| MUFA | 9.21 ± 2.22 a | 8.58 ± 1.10 a | 11.83 ± 2.55 b | 5.56 ± 0.57 c | 9.45 ± 0.91 a | 8.79 ± 0.94 a | 11.85 ± 1.94 c | 6.81 ± 1.27 b | ||
| PUFA | 52.79 ± 2.73 a | 53.30 ± 1.61 a | 50.66 ± 2.57 b | 59.03 ± 0.96 c | 54.31 ± 1.45 a | 55.85 ± 1.23 b | 51.980 ± 1.27 c | 58.30 ± 1.15 d | ||
| n-6/n-3 | 2.41 ± 0.18 a | 2.0 3 ± 0.14 b | 4.05 ± 1.14 c | 2.64 ± 0.16 d | 2.75 ± 0.29 a | 2.18 ± 0.11 b | 2.99 ± 0.64 a | 2.73 ± 0.25 a | ||



3. Experimental
3.1. Herbal Materials and Chemicals
3.2. Sample Preparation
3.3. GC-MS Analysis
3.4. Data Processing
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Chinese Pharmacopoeia Committee. The Pharmacopoeia of the People’s Republic of China; Chemical Industry Press: Beijing, China, 2005; Volume 1. [Google Scholar]
- Chen, X.J.; Guo, B.L.; Li, S.P.; Zhang, Q.W.; Tu, P.F.; Wang, Y.T. Simultaneous determination of 15 flavonoids in Epimedium using pressurized liquid extraction and high-performance liquid chromatography. J. Chromatogr. A 2007, 1163, 96–104. [Google Scholar]
- Yang, F.Q.; Wang, Y.T.; Li, S.P. Simultaneous determination of 11 characteristic components in three species of Curcuma rhizomes using pressurized liquid extraction and high-performance liquid chromatography. J. Chromatogr. A 2006, 1134, 226–231. [Google Scholar]
- Chai, X.Y.; Li, S.L.; Li, P. Quality evaluation of Flos lonicerae through a simultaneous determination of seven saponins by HPLC with ELSD. J. Chromatogr. A 2005, 1070, 43–48. [Google Scholar]
- Chinese Pharmacopoeia Committee. The Pharmacopoeia of the People’s Republic of China; Chemical Industry Publiser: Beijing, China, 2010; Volume 1. [Google Scholar]
- Xia, Q.; Zhao, K.J.; Huang, Z.G.; Zhang, P.; Dong, T.T.; Li, S.P.; Tsim, K.W. Molecular genetic and chemical assessment of Rhizoma Curcumae in China. J. Agric. Food Chem. 2005, 53, 6019–6026. [Google Scholar] [CrossRef]
- Wu, X.; Tong, Y.; Shankar, K.; Baumgardner, J.N.; Kang, J.; Badeaux, J.; Badger, T.M.; Ronis, M.J. Lipid fatty acid profile analyses in liver and serum in rats with nonalcoholic steatohepatitis using improved gas chromatography-mass spectrometry methodology. J. Agric. Food Chem. 2011, 59, 747–754. [Google Scholar] [CrossRef]
- Zhao, C.; Mao, J.; Ai, J.; Shenwu, M.; Shi, T.; Zhang, D.; Wang, X.; Wang, Y.; Deng, Y. Integrated lipidomics and transcriptomic analysis of peripheral blood reveals significantly enriched pathways in type 2 diabetes mellitus. BMC Med. Genomics 2013, 6, S12. [Google Scholar] [CrossRef]
- Wang, D.C.; Sun, C.H.; Liu, L.Y.; Sun, X.H.; Jin, X.W.; Song, W.L.; Liu, X.Q.; Wan, X.L. Serum fatty acid profiles using GC-MS and multivariate statistical analysis: Potential biomarkers of Alzheimer’s disease. Neurobiol. Aging 2012, 33, 1057–1066. [Google Scholar] [CrossRef]
- Xiong, Y.H.; Xu, Y.; Yang, L.; Wang, Z.T. Gas chromatography-mass spectrometry-based profiling of serum fatty acids in acetaminophen-induced liver injured rats. J. Appl. Toxicol. 2012. [Google Scholar] [CrossRef]
- Ozbek, A.; Aktas, O. Identification of three strains of Mycobacterium species isolated from clinical samples using fatty acid methyl ester profiling. J. Int. Med. Res. 2003, 31, 133–140. [Google Scholar] [CrossRef]
- Whittaker, P. Evaluating the use of fatty acid profiles to differentiate human pathogenic and nonpathogenic listeria species. J. AOAC Int. 2012, 95, 1457–1459. [Google Scholar] [CrossRef]
- Zhang, X.J.; Huang, L.L.; Cai, X.J.; Li, P.; Wang, Y.T.; Wan, J.B. Fatty acid variability in three medicinal herbs of Panax species. Chem. Cent. J. 2013, 7, 12. [Google Scholar] [CrossRef]
- Taylor, J.; King, R.D.; Altmann, T.; Fiehn, O. Application of metabolomics to plant genotype discrimination using statistics and machine learning. Bioinformatics 2002, 18, S241–S248. [Google Scholar]
- Lan, F.Y. Pharmacological activity and clinical application of Yujin. Changchun Zhongyiyao Daxue Xuebao 2009, 25, 27–28. [Google Scholar]
- Araujo, C.C.; Leon, L.L. Biological activities of Curcuma longa L. Mem. Inst. Oswaldo Cruz 2001, 96, 723–728. [Google Scholar] [CrossRef]
- Zahid Ashraf, M.; Hussain, M.E.; Fahim, M. Antiatherosclerotic effects of dietary supplementations of garlic and turmeric: Restoration of endothelial function in rats. Life Sci. 2005, 77, 837–857. [Google Scholar] [CrossRef]
- Yang, F.Q.; Li, S.P.; Zhao, J.; Lao, S.C.; Wang, Y.T. Optimization of GC-MS conditions based on resolution and stability of analytes for simultaneous determination of nine sesquiterpenoids in three species of Curcuma rhizomes. J. Pharm. Biomed. Anal. 2007, 43, 73–82. [Google Scholar] [CrossRef]
- Zhang, J.S.; Guan, J.; Yang, F.Q.; Liu, H.G.; Cheng, X.J.; Li, S.P. Qualitative and quantitative analysis of four species of Curcuma rhizomes using twice development thin layer chromatography. J. Pharm. Biomed. Anal. 2008, 48, 1024–1028. [Google Scholar] [CrossRef]
- Lechtenberg, M.; Quandt, B.; Nahrstedt, A. Quantitative determination of curcuminoids in Curcuma rhizomes and rapid differentiation of Curcuma domestica Val. and Curcuma xanthorrhiza Roxb. by capillary electrophoresis. Phytochem. Anal. 2004, 15, 152–158. [Google Scholar] [CrossRef]
- Xiang, Z.; Wang, X.Q.; Cai, X.J.; Zeng, S. Metabolomics study on quality control and discrimination of three curcuma species based on gas chromatograph-mass spectrometry. Phytochem. Analysis 2011, 22, 411–418. [Google Scholar] [CrossRef]
- Wiklund, S. Multivariate Data Analysis for Omics; Umetrics: Umeå, Sweden, 2008. [Google Scholar]
- Wan, J.B.; Bai, X.; Cai, X.J.; Rao, Y.; Wang, Y.S.; Wang, Y.T. Chemical differentiation of Da-Cheng-Qi-Tang, A Chinese medicine formula, prepared by traditional and modern decoction methods using UPLC/Q-TOFMS-based metabolomics approach. J. Pharm. Biomed. Anal. 2013, 83, 34–42. [Google Scholar] [CrossRef]
- Araujo, P.; Nguyen, T.T.; Froyland, L.; Wang, J.; Kang, J.X. Evaluation of a rapid method for the quantitative analysis of fatty acids in various matrices. J. Chromatogr. A 2008, 1212, 106–113. [Google Scholar]
- Kang, J.X.; Wang, J. A simplified method for analysis of polyunsaturated fatty acids. BMC Biochem. 2005, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, Y.; Hao, F.; Zhou, X.; Han, X.; Tang, H.; Ji, L. Human serum metabonomic analysis reveals progression axes for glucose intolerance and insulin resistance statuses. J. Proteome Res. 2009, 8, 5188–5195. [Google Scholar] [CrossRef]
- Koh, Y.; Pasikanti, K.K.; Yap, C.W.; Chan, E.C. Comparative evaluation of software for retention time alignment of gas chromatography/time-of-flight mass spectrometry-based metabonomic data. J. Chromatogr. A 2010, 1217, 8308–8316. [Google Scholar]
- Sample Availability: Samples of the Curcuma plant materials are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, X.-J.; Qiu, J.-F.; Guo, L.-P.; Wang, Y.; Li, P.; Yang, F.-Q.; Su, H.; Wan, J.-B. Discrimination of Multi-Origin Chinese Herbal Medicines Using Gas Chromatography-Mass Spectrometry-Based Fatty Acid Profiling. Molecules 2013, 18, 15329-15343. https://doi.org/10.3390/molecules181215329
Zhang X-J, Qiu J-F, Guo L-P, Wang Y, Li P, Yang F-Q, Su H, Wan J-B. Discrimination of Multi-Origin Chinese Herbal Medicines Using Gas Chromatography-Mass Spectrometry-Based Fatty Acid Profiling. Molecules. 2013; 18(12):15329-15343. https://doi.org/10.3390/molecules181215329
Chicago/Turabian StyleZhang, Xiao-Jing, Jian-Feng Qiu, Lan-Ping Guo, Ying Wang, Peng Li, Feng-Qing Yang, Huanxing Su, and Jian-Bo Wan. 2013. "Discrimination of Multi-Origin Chinese Herbal Medicines Using Gas Chromatography-Mass Spectrometry-Based Fatty Acid Profiling" Molecules 18, no. 12: 15329-15343. https://doi.org/10.3390/molecules181215329