Cyclic versus Hemi-Bastadins. Pleiotropic Anti-Cancer Effects: from Apoptosis to Anti-Angiogenic and Anti-Migratory Effects

Abstract
:
1. Introduction

2. Results and Discussion
2.1. In Vitro Growth Inhibitory Concentrations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Carcinoma | Glioma | Melanoma | Mean ± SEM | |||
|---|---|---|---|---|---|---|---|
| MCF-7 (breast) | A549 (NSCLC) | Hs683 | U373 | B16F10 * | SKMEL28 | ||
| (oligodendroglioma) | (astroglioma) | ||||||
| 1 | 4 | 3 | 3 | 3 | 4 | 4 | 4.0 ± 0.2 |
| 2 | 8 | 7 | 4 | 7 | 5 | 7 | 6.0 ± 0.6 |
| 3 | 7 | 8 | 4 | 11 | 6 | 7 | 7.0 ± 0.9 |
| 4 | >100 | >100 | >100 | >100 | 75 | >100 | >96 |
| 5 | 94 | >100 | >100 | >100 | 86 | >100 | >97 |
| 6 | >100 | >100 | >100 | >100 | 45 | >100 | >91 |
| 7 | >100 | >100 | >100 | >100 | 63 | >100 | >94 |
| 8 | 68 | 68 | 70 | 73 | 58 | 76 | 69 ± 3 |
2.2. Quantitative Videomicroscopy Analyses

2.3. Cell Cycle Kinetics versus Pro-Apoptotic Features

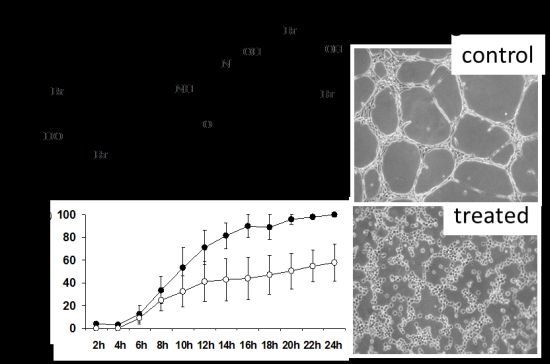
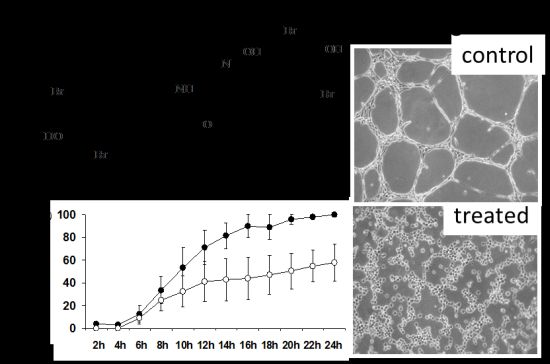
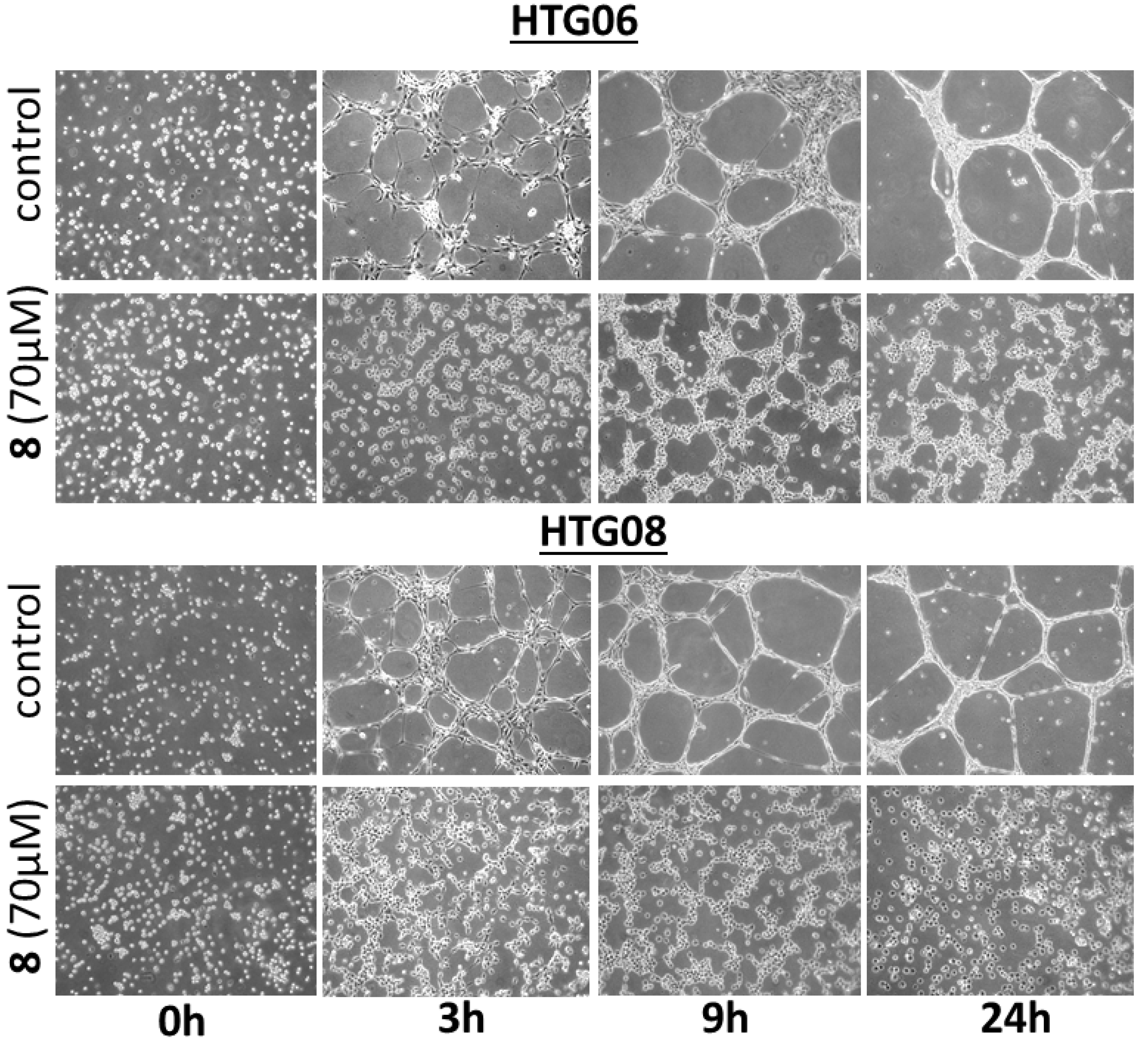
2.4. DBHB (8) Exhibits Anti-Angiogenic Activity

2.5. Direct Impact of Serum on the Anti-Migratory Properties of DBHB (8) in B16F10 Melanoma Cells, with a Potential Involvement of Albumin

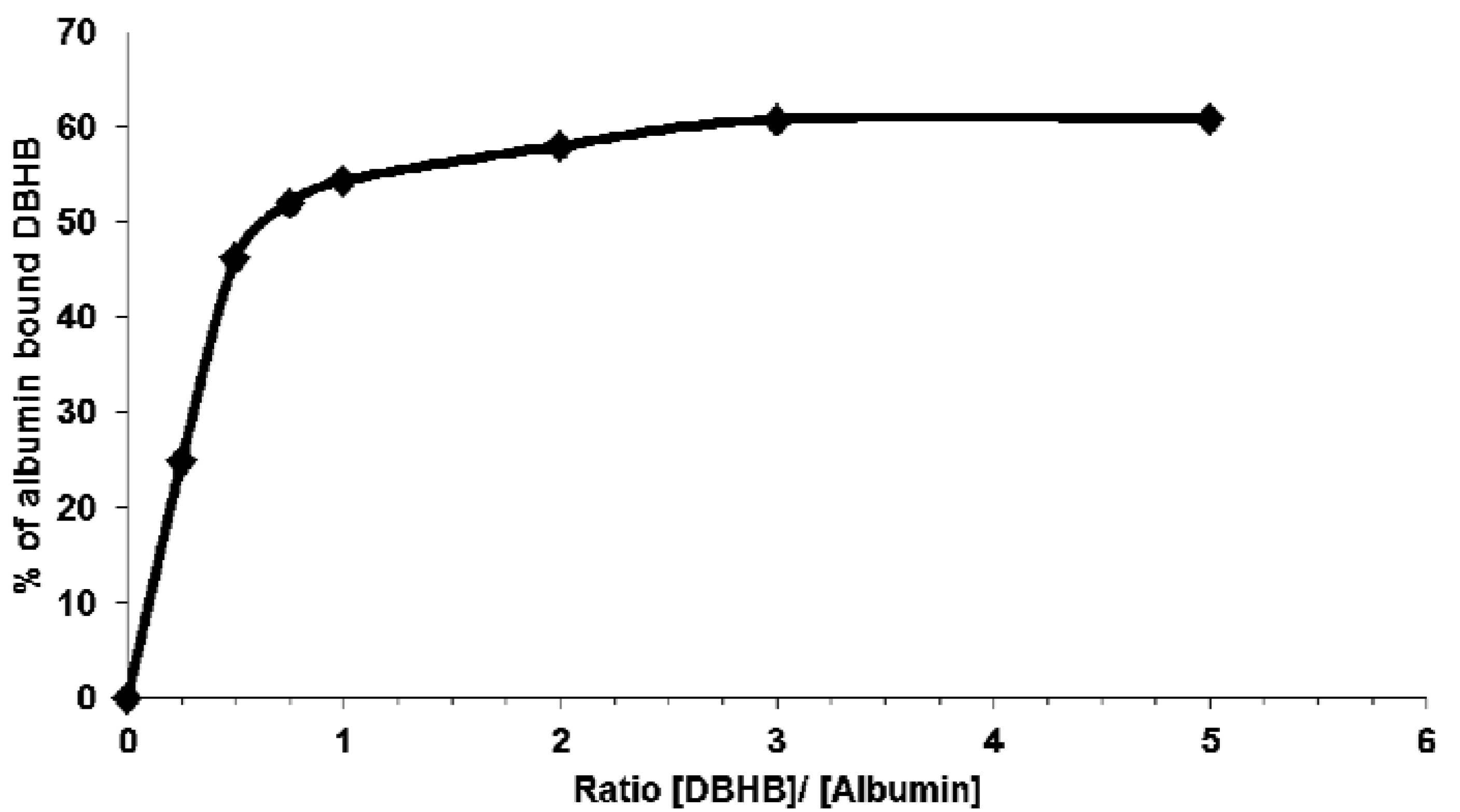
2.6. In Vivo Analyses of DBHB (8) Activity in B16F10 Melanoma-Bearing Mice
3. Experimental
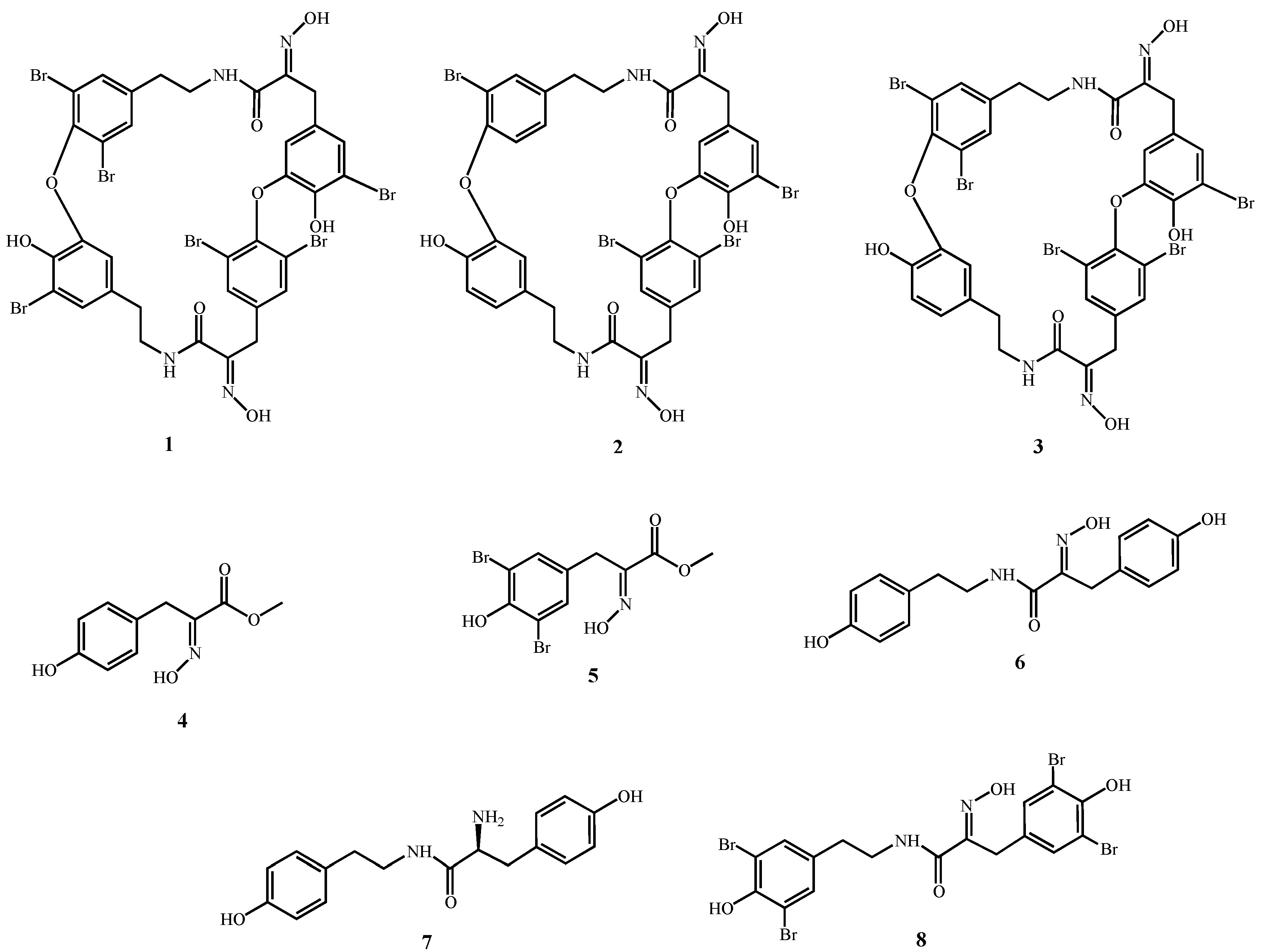
3.1. Sample Collection, Extraction and Purification of bastadin 6 (1), bastadin 9 (2) and bastadin 16 (3)
3.2. Chemical Syntheses
3.3. Determination of the In Vitro Growth Inhibitory Concentrations
3.4. Computer-Assisted Phase Contrast Microscopy (Quantitative Videomicroscopy)
3.5. Flow Cytometry
3.6. In Vitro Anti-Angiogenesis Analyses
3.7. Formulating DBHB (8) for in Vivo Analyses
3.8. The in Vivo Model of Lung Pseudometastases from Mouse B16F10 Melanoma
3.9. Scratch Wound Assay
3.10. Determination of DBHB (8) Affinity to Albumin
4. Conclusions
Acknowledgments
References
- Kazlauskas, R.; Lidgard, R.O.; Murphy, P.T.; Well, R.J. Brominated tyrosine-derived metabolites from the sponge Ianthella basta. Tetrahedron Lett. 1980, 21, 2277–2280. [Google Scholar] [CrossRef]
- Kazlauskas, R.; Lidgard, R.O.; Murphy, P.T.; Well, R.J.; Blount, J.F. Brominated tyrosine-derived metabolites from the sponge Ianthella basta. Aust. J. Chem. 1981, 34, 765–786. [Google Scholar] [CrossRef]
- Park, S.K.; Park, H.; Scheuer, P.J. Isolation and structure determination of a new bastadins from the marine sponge Ianthella basta. Bull. Korean Chem. Soc. 1994, 15, 534–537. [Google Scholar]
- Pettit, G.R.; Butler, M.S.; Bass, C.G.; Doubek, D.L.; Williams, M.D.; Schmidt, J.M.; Pettit, R.K.; Hooper, J.N.; Tackett, L.P.; Filiatrault, M.J. Antineoplastic agents, 326. The stereochemistry of bastadins 8, 10, and 12 from the Bismarck archipelago marine sponge Ianthella basta. J. Nat. Prod. 1995, 58, 680–688. [Google Scholar] [CrossRef]
- Coll, J.C.; Kearns, P.S.; Rideout, J.A.; Sankar, V. Bastadin 21, a novel isobastarane metabolite from the Great Barrier Reef marine sponge Ianthella quadrangulata. J. Nat. Prod. 2002, 65, 753–756. [Google Scholar] [CrossRef]
- Calcul, L.; Inman, W.D.; Morris, A.A.; Tenney, K.; Ratman, J.; McKerrow, J.H.; Valeriote, F.A.; Crews, P. Additional insights on the bastadins: Isolation of analogues from the sponge Ianthella cf. reticulata and exploration of the oxime configurations. J. Nat. Prod. 2010, 73, 365–372. [Google Scholar] [CrossRef]
- Carroll, A.R.; Kaiser, S.M.; Davis, R.A.; Moni, R.W.; Hooper, J.N.; Quinn, R.J. A bastadins with potent and selective delta-opioid receptor binding affinity from the Australian sponge Ianthella flabelliformis. J. Nat. Prod. 2010, 73, 1173–1176. [Google Scholar] [CrossRef]
- Couladouros, E.A.; Pitsinos, E.N.; Moutsos, V.I.; Sarakinos, G. A general method for the synthesis of bastaranes and isobastaranes: First total synthesis of bastadins 5, 10, 12, 16, 20, and 21. Chemistry 2004, 11, 406–421. [Google Scholar]
- Masuno, M.N.; Pessah, I.N.; Olmstead, M.M.; Molinski, T.F. Simplified cyclic analogues of bastadins-5. Structure-activity relationships for modulation of the Ryr1/FKBP12 Ca2+ channel complex. J. Med. Chem. 2006, 49, 4497–4511. [Google Scholar] [CrossRef]
- Zieminska, E.; Lazarewicz, JW.; Couladouros, E.A.; Moutsos, V.I.; Pitsinos, E.N. Open-chain half-bastadins mimic the effects of cyclic bastadins and calcium homeostasis in cultured neurons. Bioorg. Med. Chem. Lett. 2008, 18, 5734–5737. [Google Scholar]
- Ortlepp, S.; Sjögren, M.; Dahlström, M.; Weber, H.; Ebel, R.; Edrada, R.; Thoms, C.; Schupp, P.; Bohlin, L.; Proksch, P. Antifouling activity of bromotyrosine-derived sponge metabolites and synthetic analogues. Mar. Biotechnol. 2007, 9, 776–785. [Google Scholar] [CrossRef]
- Bayer, M.; Hellio, C.; Maréchal, J.P.; Frank, W.; Lin, W.; Weber, H.; Proksch, P. Antifouling bastadins congeners target mussel phenoloxidase and complex copper(II) ions. Mar. Biotechnol. 2011, 13, 1148–1158. [Google Scholar] [CrossRef]
- Chen, L.; Molinski, T.F.; Pessah, I.N. Bastadin 10 stabilizes the open conformation of the ryanodine-sensitive Ca2+ channel in an FKBP12-dependent manner. J. Biol. Chem. 1999, 274, 32603–32612. [Google Scholar] [CrossRef]
- Masuno, M.N.; Hoepker, A.C.; Pessah, I.N.; Molinski, T.F. 1-O-Sulfatobastadins-1 and -2 from Lanthella basta (Pallas). Antagonists of the RyR1-FKBP12 Ca2+ channel. Mar. Drugs 2004, 2, 176–184. [Google Scholar] [CrossRef]
- Zieminska, E.; Stafiej, A.; Pitsinos, E.N.; Couladouros, E.A.; Moutsos, V.; Kozlowska, H.; Toczylowska, B.; Lazarewicz, J.W. Synthetic bastadins modify the activity of ryanodine receptors in cultured cerebellar granule cells. Neurosignals 2006, 15, 283–292. [Google Scholar] [CrossRef]
- Reddy, A.V.; Ravinder, K.; Narasimhulu, M.; Sridevi, A.; Satyanarayana, N.; Kondapi, A.K.; Venkateswarlu, Y. New anticancer bastadins alkaloids from the sponge Dendrilla cactos. Bioorg. Med. Chem. 2006, 14, 4452–4457. [Google Scholar] [CrossRef]
- Greve, H.; Kehraus, S.; Krick, A.; Kelter, G.; Maier, A.; Fiebig, H.H.; Wright, A.D.; König, G.M. Cytotoxic bastadin 24 from the Australian sponge Ianthella quadrangulata. J. Nat. Prod. 2008, 71, 309–312. [Google Scholar] [CrossRef]
- Kotoku, N.; Tsujita, H.; Hiramatsu, A.; Mori, C.; Koizumi, N.; Kobayashi, M. Efficient total synthesis of bastadins 6, an anti-angiogenic brominated tyrosine-derived metabolite from marine sponge. Tetrahedron 2005, 61, 7211–7218. [Google Scholar]
- Kotoku, N.; Hiramatsu, A.; Tsujita, H.; Hirakawa, Y.; Sanagawa, M.; Aoki, S.; Kobayashi, M. Structure-activity relationship study of bastadins 6, an anti-angiogenic brominated-tyrosine derived metabolite from marine sponge. Arch. Pharm. 2008, 341, 568–577. [Google Scholar] [CrossRef]
- Aoki, S.; Cho, S.H.; Hiramatsu, A.; Kotoku, N.; Kobayashi, M. Bastadins, Cyclic tetramers of brominated-tyrosine derivatives, Selectively inhibit the proliferation of endothelial cells. J. Nat. Med. 2006, 60, 231–235. [Google Scholar] [CrossRef]
- Aoki, S.; Cho, S.H.; Ono, M.; Kuwano, T.; Nakao, S.; Kuwano, M.; Nakagawa, S.; Gao, J.Q.; Mayumi, T.; Shibuya, M.; Kobayashi, M. Bastadin 6, a spongean brominated tyrosine derivative, inhibits tumor angiogenesis by inducing selective apoptosis to endothelial cells. Anticancer Drugs 2006, 17, 269–278. [Google Scholar] [CrossRef]
- Chen, K.G.; Sikic, B.I. Molecular pathways: Regulation and therapeutic implications of multidrug resistance. Clin. Cancer Res. 2012, 18, 1863–1869. [Google Scholar] [CrossRef]
- Lefranc, F.; Sadeghi, N.; Camby, I.; Metens, T.; De Witte, O.; Kiss, R. Present and potential future issues in glioblastoma treatment. Expert Rev. Anticancer Ther. 2006, 6, 719–732. [Google Scholar] [CrossRef]
- Maira, F.; Catania, A.; Candido, S.; Russo, A.E.; McCubrey, J.A.; Libra, M.; Malaponte, G.; Fenga, C. Molecular targeted therapy in melanoma: A way to reverse resistance to conventional drugs. Curr. Drug Deliv. 2012, 9, 17–29. [Google Scholar] [CrossRef]
- Fennell, D.A.; Swanton, C. Unlocking Pandora’s box: personalizing cancer cell death in non-small cell lung cancer. EPMA J. 2012, 3, 6. [Google Scholar] [CrossRef]
- Bruyère, C.; Lonez, C.; Duray, A.; Cludts, S.; Ruysschaert, J.M.; Saussez, S.; Yeaton, P.; Kiss, R.; Mijatovic, T. Considering temozolomide as a novel potential treatment for esophageal cancer. Cancer 2011, 117, 2004–2016. [Google Scholar] [CrossRef]
- Nagaprashantha, L.; Vartak, N.; Awasthi, S.; Awasthi, S.; Singhal, S.S. Novel anti-cancer compounds for developing combinatorial therapies to target anoïkis-resistant tumors. Pharm. Res. 2012, 29, 621–636. [Google Scholar] [CrossRef]
- Okayama, H. Cell cycle control by floatage signaling. Cell Signal. 2012, 24, 1599–1609. [Google Scholar] [CrossRef]
- Mathieu, V.; Mijatovic, T.; Van Damme, M.; Kiss, R. Gastrin exerts pleitropic effects on human melanoma cell biology. Neoplasia 2005, 7, 930–943. [Google Scholar] [CrossRef]
- Mégalizzi, V.; Mathieu, V.; Mijatovic, T.; Gailly, P.; Debeir, O.; De Neve, N.; Van Damme, M.; Bontempi, G.; Haibe-Kains, B.; Decaestecker, C.; et al. 4-IBP, A sigma1 receptor agonist, Decreases the migration of human cancer cells, Including glioblastoma cells, in vitro and sensitizes them in vitro and in vivo to cytotoxic insults of proapoptotic and proautophagic drugs. Neoplasia 2007, 9, 358–369. [Google Scholar] [CrossRef]
- Tabatabai, G.; Tonn, J.C.; Stupp, R.; Weller, M. The role of integrins in glioma biology and anti-glioma therapies. Curr. Pharm. Des. 2011, 17, 2402–2410. [Google Scholar] [CrossRef]
- Svensen, N.; Walton, J.G.; Bradley, M. Peptides for cell-selective drug delivery. Trends Pharmacol. Sci. 2012, 33, 186–192. [Google Scholar] [CrossRef]
- Mathieu, A.; Remmelink, M.; D’Haene, N.; Penant, S.; Gaussin, J.F.; Van Ginckel, R.; Darro, F.; Kiss, R.; Salmon, I. Development of a chemoresistant orthotopic human non-small cell lung carcinoma models in nude mice: Analyses of tumor heterogeneity in relation to the immunohistochemical levels of expression of ciclooxygenase-2, ornitine decarboxylase, lung-related resistance protein, prostaglandine-E synthetase, and glutathione-S-transferase-alpha (GST)-alpha, GST-mu, and GST-pi. Cancer 2004, 101, 1908–1918. [Google Scholar] [CrossRef]
- Van Goietsenoven, G.; Hutton, J.; Becker, J.P.; Lallemand, B.; Robert, F.; Lefranc, F.; Pirker, C.; Vandenbussche, G.; Van Antwerpen, P.; Evidente, A.; et al. Targeting of eEF1A with Amaryllidaceae isocarbostyrils as a strategy to combat melanomas. FASEB J. 2010, 24, 4575–4584. [Google Scholar] [CrossRef]
- Ingrassia, L.; Lefranc, F.; Dewelle, J.; Pottier, L.; Mathieu, V.; Spiegl-Kreinecker, S.; Sauvage, S.; El Yazidi, M.; Dehoux, M.; Berger, W.; et al. Structure-activity relationship analysis of novel derivatives of narciclasine (an Amaryllidaceae isocarbostyril derivative) as potential anticancer agents. J. Med. Chem. 2009, 52, 1100–1104. [Google Scholar] [CrossRef]
- Dumont, P.; Ingrassia, L.; Rouzeau, S.; Ribaucour, F.; Thomas, S.; Roland, I.; Darro, F.; Lefranc, F.; Kiss, R. The amaryllidaceae isocarbostyril narciclasine induces apoptosis by activation of the death receptor and/or mitochondrial pathways in cancer cells but not in normal fibroblasts. Neoplasia 2007, 9, 766–776. [Google Scholar] [CrossRef]
- Goffin, E.; Lamoral-Theys, D.; Tajedinne, N.; de Tullio, P.; Mondin, L.; Lefranc, F.; Gailly, P.; Rogister, B.; Kiss, R.; Pirotte, B. N-Aryl-N’-(chroman-4-yl)ureas and thioureas display in vitro anticancer activity and selectivity on apoptosis-resistant glioblastoma cells: Screening, synthesis of simplified derivatives, and structure-activity relationship analysis. Eur. J. Med. Chem. 2012, 54, 834–844. [Google Scholar] [CrossRef]
- Lallemand, B.; Masi, M.; Maddau, L.; De Lorenzi, M.; Dam, R.; Cimmino, A.; Moreno Y Banuls, L.; Andolfi, A.; Kiss, R.; Mathieu, V.; et al. Evaluation of in vitro anticancer activity of sphaeropsidins A-C, fungal rearranged pimarane diterpenes, and semisynthetic derivatives. Phytochem. Lett. 2012, 5, 770–775. [Google Scholar] [CrossRef]
- Delbrouck, C.; Doyen, I.; Belot, N.; Decaestecker, C.; Ghanooni, R.; de Lavareille, A.; Kaltner, H.; Choufani, G.; Danguy, A.; Vandenhoven, G.; et al. Galectin-1 is overexpressed in nasal polyps under budesonide and inhibits eosinophil migration. Lab. Invest. 2002, 82, 147–158. [Google Scholar]
- Guzman, R.E.; Bolanos, P.; Delgado, A.; Rojas, H.; DiPolo, R.; Caputo, C.; Jaffe, E.H. Depolymerization and rearrangement of actin filaments during exocytosis in rat peritoneal mast cells: involvement of ryanodine-sensitive calcium stores. Pflugers Arch. 2007, 454, 131–141. [Google Scholar] [CrossRef]
- Bose, D.D.; Thomas, D.W. The actin cytoskeleton differentially regulates NG115–401L cell ryanodine receptor and inositol 1,4,5-triphosphate receptor induced calcium signaling pathways. Biochem. Biophys. Res. Commun. 2009, 379, 594–599. [Google Scholar] [CrossRef]
- Lefranc, F.; Mijatovic, T.; Mathieu, V.; Rorive, S.; Decaestecker, C.; Debeir, O.; Brotchi, J.; Van Ham, P.; Salmon, I.; Kiss, R. Characterization of gastrin-induced proangiogenic effects in vivo in orthotopic U373 experimental human glioblastomas and in vitro in human umbilical vein endothelial cells. Clin. Cancer Res. 2004, 10, 8250–8265. [Google Scholar] [CrossRef]
- Mackrill, J.J. Ryanodine receptor calcium channels and their partners as drug targets. Biochem. Pharmacol. 2010, 79, 1535–1543. [Google Scholar] [CrossRef]
- Mathieu, V.; Le Mercier, M.; De Neve, N.; Sauvage, S.; Gras, T.; Roland, I.; Lefranc, F.; Kiss, R. Galectin-1 knockdown increases sensitivity to temozlomide in a B16F10 mouse metastatic melanoma model. J. Invest. Dermatol. 2007, 127, 2399–2410. [Google Scholar] [CrossRef]
- Wauthoz, N.; Deleuze, P.; Hecq, J.; Roland, I.; Saussez, S.; Adanja, I.; Debeir, O.; Decaestecker, C.; Mathieu, V.; Kiss, R.; et al. In vivo assessment of temozolomide local delivery for lung cancer inhalation therapy. Eur. J. Pharm. Sci. 2010, 39, 402–411. [Google Scholar] [CrossRef]
- Wauthoz, N.; Deleuze, P.; Saumet, A.; Duret, C.; Kiss, R.; Amighi, K. Temozolomide-based dry powder formulations for lung tumor-related inhalation treatment. Pharm. Res. 2011, 28, 762–775. [Google Scholar] [CrossRef]
- Black, A.; Morris, D. Personalized medicine in metastatic non-small-cell lung cancer: Promising targets and current clinical trials. Curr. Oncol. 2012, 19, S73–S85. [Google Scholar]
- Ganguli, A.; Wiegand, P.; Gao, X.; Carter, J.A.; Botterman, M.F.; Ray, S. The impact of second-line agents on patients’ health-related quality of life in the treatment for non-small cell lung cancer: A systematic review. Qual. Life Res. 2012, in press.
- Nakata, A.; Gotoh, N. Recent understanding of the molecular mechanisms for the efficacy and resistance of EGF receptor-specific tyrosine kinase inhibitors in non-small cell lung cancer. Expert Opin. Ther. Targets 2012, 16, 771–781. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds 1–8 are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mathieu, V.; Wauthoz, N.; Lefranc, F.; Niemann, H.; Amighi, K.; Kiss, R.; Proksch, P. Cyclic versus Hemi-Bastadins. Pleiotropic Anti-Cancer Effects: from Apoptosis to Anti-Angiogenic and Anti-Migratory Effects. Molecules 2013, 18, 3543-3561. https://doi.org/10.3390/molecules18033543
Mathieu V, Wauthoz N, Lefranc F, Niemann H, Amighi K, Kiss R, Proksch P. Cyclic versus Hemi-Bastadins. Pleiotropic Anti-Cancer Effects: from Apoptosis to Anti-Angiogenic and Anti-Migratory Effects. Molecules. 2013; 18(3):3543-3561. https://doi.org/10.3390/molecules18033543
Chicago/Turabian StyleMathieu, Véronique, Nathalie Wauthoz, Florence Lefranc, Hendrik Niemann, Karim Amighi, Robert Kiss, and Peter Proksch. 2013. "Cyclic versus Hemi-Bastadins. Pleiotropic Anti-Cancer Effects: from Apoptosis to Anti-Angiogenic and Anti-Migratory Effects" Molecules 18, no. 3: 3543-3561. https://doi.org/10.3390/molecules18033543