Dendrimers as Potential Therapeutic Tools in HIV Inhibition

Abstract
:1. Introduction
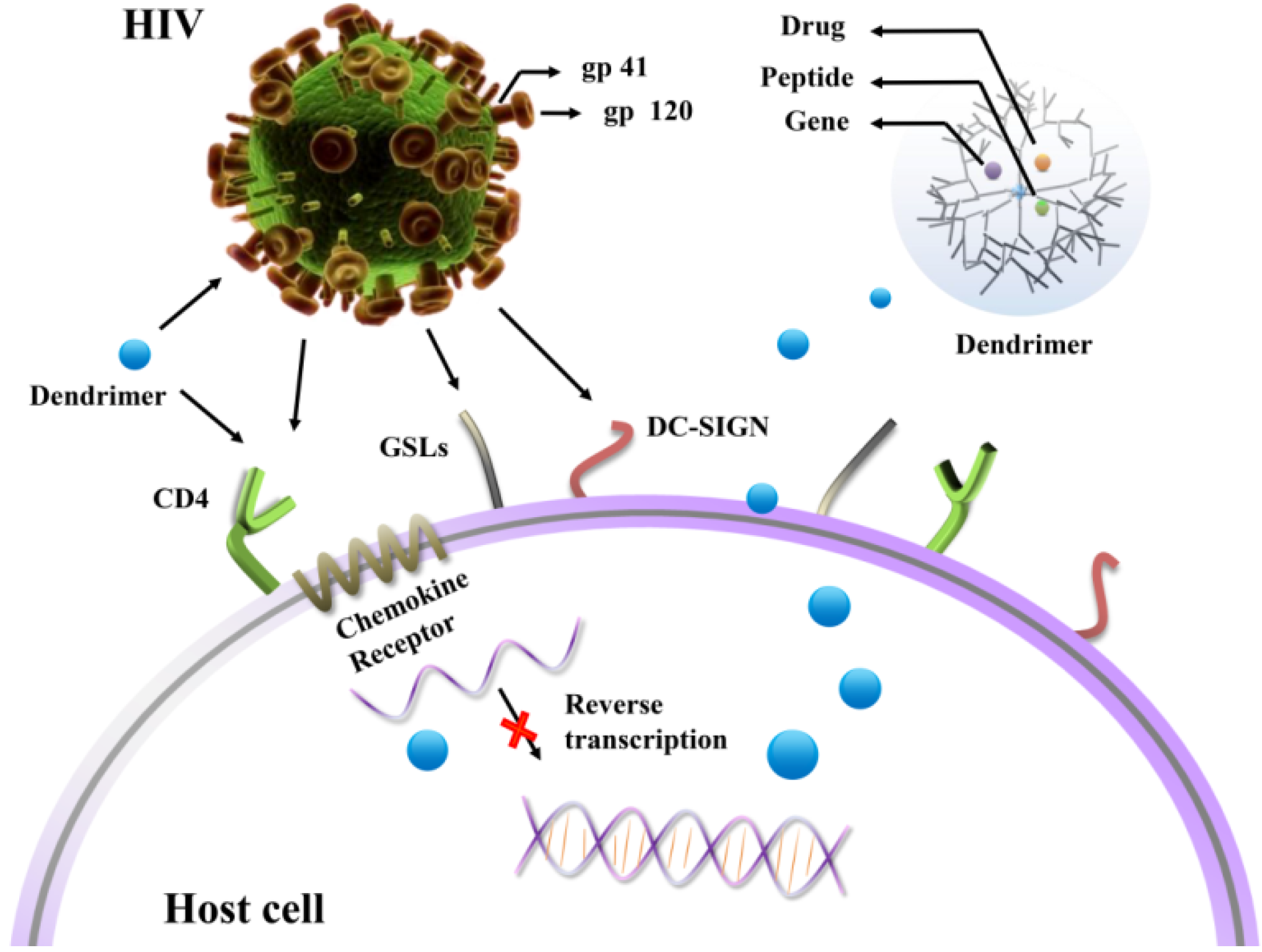
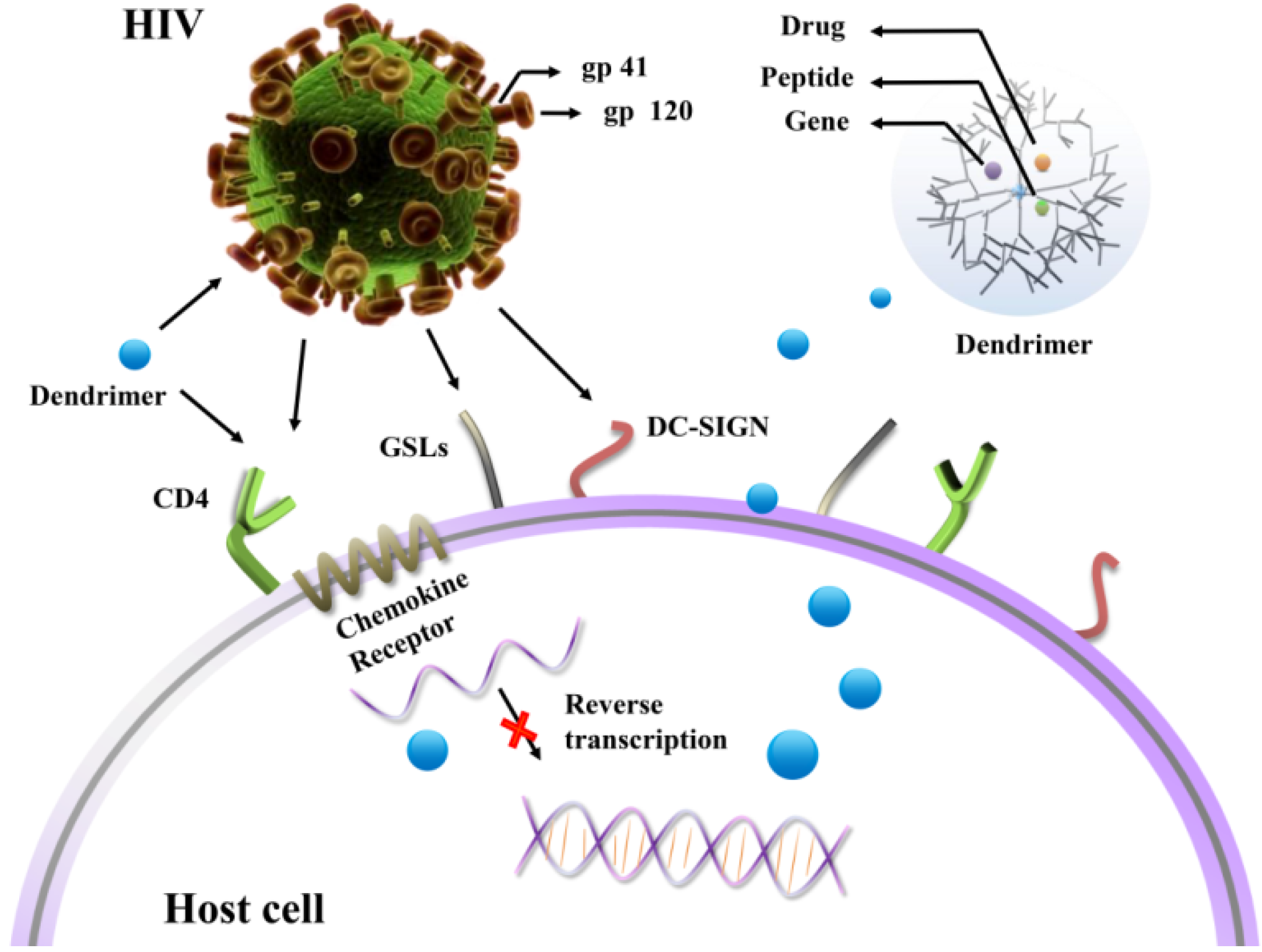
2. Functionalized Dendrimers as Potential Anti-HIV Agents

{kind=link}
{kind=link}
| Code name and reference | Dendrimer classification | Functional end groups | Combination sites | Stage of inhibition |
|---|---|---|---|---|
| BRI2932 (SPL2923) [7] | PAMAM G4 |  | gp120 | attachment and replication |
| BRI6195 (SPL6195) [7] | PAMAM G4 |  | gp120 | attachment |
| SPL7013 [8,9] | Polylysine dendrimer |  | gp120 | attachmentand replication |
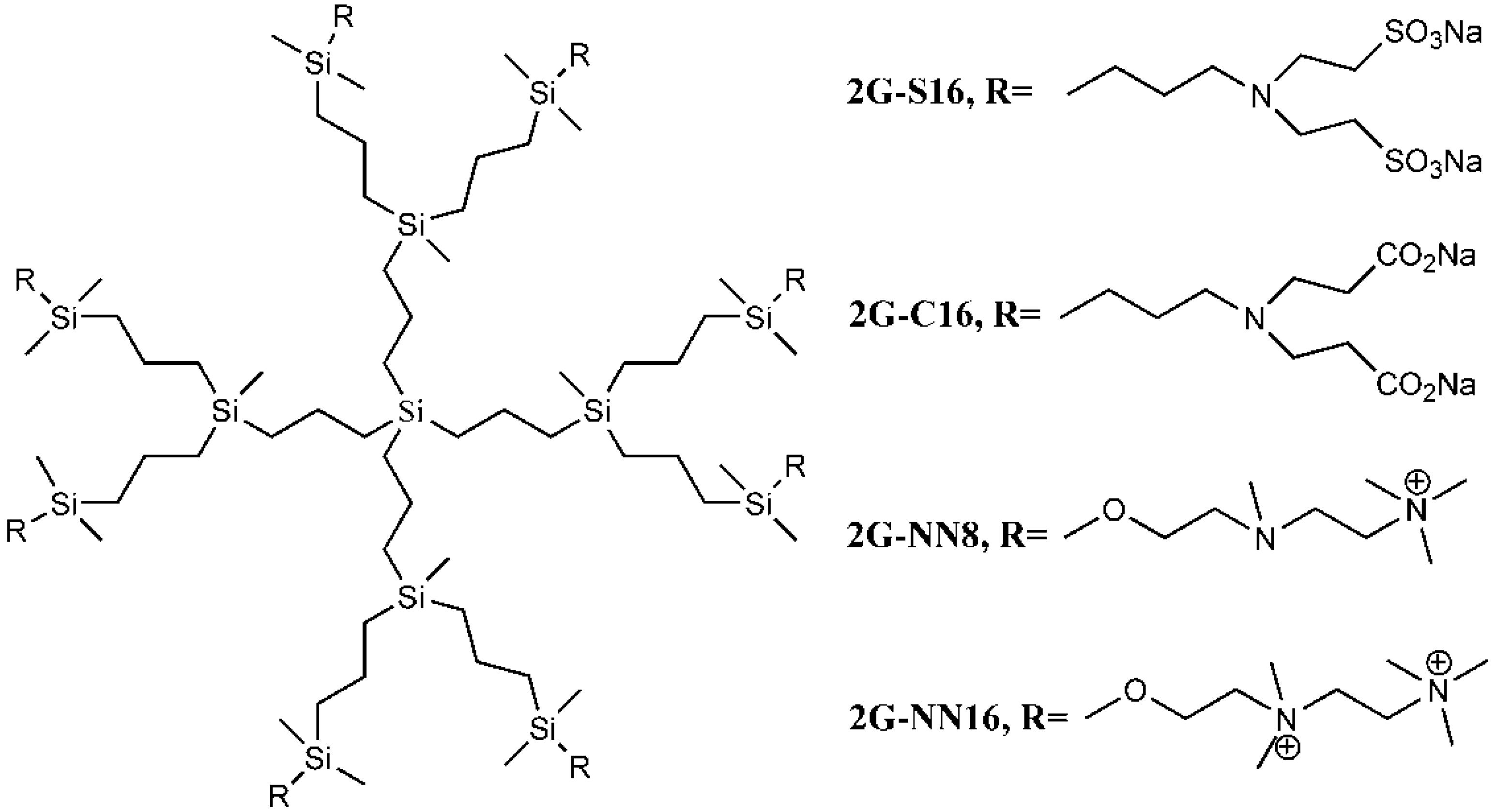
| 2G-S16 [10,11] | Carbosilane Dendrimer |  | gp120/CD4 | attachment |
| 2G-C16 [11] | Carbosilane Dendrimer |  | N/A | N/A |
| PS Gal 64mer [12] | Polypropylenimine tetrahexaconta- amine dendrimer |  | gp120 | attachment |
| PLDG3 and SCSLD3 [13,14] | Polylysine and amphiphilic lysine dendrimer |  | gp120 | attachment |
| MVC-GBT [15] | Polypropylenimine dendrimer |  | gp120 | attachment |
| MVC-3SL [15] | Polypropylenimine dendrimer |  | gp120 | attachment |
| [G1]-CO2Na [16] | GATG dendrimer |  | C-terminal domain of capsid | assembly |
| PAMAM [17,18] | PAMAM | -NH2 | trans-acting responsive (TAR) RNA | replication |
2.1. Prevention of HIV Attachment to the Surface of Host Cells
2.1.1. CD4 Receptors

2.1.2. GSLs
2.1.3. DC-SIGN Receptors
2.2. Prevention of Replication of HIV in Host Cells
2.2.1. Polyanionic Dendrimers
2.2.2. Polycationic Dendrimers
3. Dendrimers Acting as Potential Vectors for Drugs and Genes
3.1. Dendrimers as Carriers for Chemical Drugs
3.2. Dendrimers as Carriers for Peptides
3.3. Dendrimers as Carriers for Genes
3.3.1. ODNs
3.3.2. siRNA
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- UNAIDS. World Aids Day Report; UNAIDS BerlinGeneva: Geneva, Swizerland, 2012. [Google Scholar]
- De Clercq, E. Antiviral therapy for human immunodeficiency virus infections. Clin. Microbiol. Rev. 1995, 8, 200–239. [Google Scholar]
- Fréchet, J.M.; Tomalia, D.A. Dendrimers and Other Dendritic Polymers; Wiley: New York, NY, USA, 2001. [Google Scholar]
- Jiang, Y.-H.; Emau, P.; Cairns, J.S.; Flanary, L.; Morton, W.R.; McCarthy, T.D.; Tsai, C.-C. SPL7013 gel as a topical microbicide for prevention of vaginal transmission of SHIV89. 6P in macaques. AIDS Res. Hum. Retroviruses 2005, 21, 207–213. [Google Scholar] [CrossRef]
- Svenson, S.; Tomalia, D.A. Dendrimers in biomedical applications—Reflections on the field. Adv. Drug Deliv. Rev. 2005, 57, 2106–2129. [Google Scholar] [CrossRef]
- D'Emanuele, A.; Attwood, D. Dendrimer-Drug interactions. Adv. Drug. Deliv. Rev. 2005, 57, 2147–2162. [Google Scholar] [CrossRef]
- Witvrouw, M.; Fikkert, V.; Pluymers, W.; Matthews, B.; Mardel, K.; Schols, D.; Raff, J.; Debyser, Z.; De Clercq, E.; Holan, G. Polyanionic (i.e., polysulfonate) dendrimers can inhibit the replication of human immunodeficiency virus by interfering with both virus adsorption and later steps (reverse transcriptase/integrase) in the virus replicative cycle. Mol. Pharmacol. 2000, 58, 1100–1108. [Google Scholar]
- Tyssen, D.; Henderson, S.A.; Johnson, A.; Sterjovski, J.; Moore, K.; La, J.; Zanin, M.; Sonza, S.; Karellas, P.; Giannis, M.P. Structure activity relationship of dendrimer microbicides with dual action antiviral activity. PLoS One 2010, 5, e12309. [Google Scholar] [CrossRef]
- Telwatte, S.; Moore, K.; Johnson, A.; Tyssen, D.; Sterjovski, J.; Aldunate, M.; Gorry, P.R.; Ramsland, P.A.; Lewis, G.R.; Paull, J.R. Virucidal activity of the dendrimer microbicide SPL7013 against HIV-1. Antiviral Res. 2011, 90, 195–199. [Google Scholar] [CrossRef]
- Chonco, L.; Pion, M.; Vacas, E.; Rasines, B.; Maly, M.; Serramía, M.; López-Fernández, L.; De la Mata, J.; Alvarez, S.; Gómez, R. Carbosilane dendrimer nanotechnology outlines of the broad HIV blocker profile. J. Control Release 2012, 161, 949–958. [Google Scholar] [CrossRef]
- Galan, M.; Sanchez-Rodriguez, J.; Cangiotti, M.; Garcia-Gallego, S.; Jimenez, J.; Gomez, R.; Ottaviani, M.; Munoz-Fernandez, M.; de la Mata, F. Antiviral Properties Against HIV of Water Soluble Copper Carbosilane Dendrimers and their EPR Characterization. Curr. Med. Chem. 2012, 19, 4984–4994. [Google Scholar] [CrossRef]
- Kensinger, R.D.; Catalone, B.J.; Krebs, F.C.; Wigdahl, B.; Schengrund, C.-L. Novel polysulfated galactose-derivatized dendrimers as binding antagonists of human immunodeficiency virus type 1 infection. Antimicrob. Agents Chemother. 2004, 48, 1614–1623. [Google Scholar] [CrossRef]
- Han, S.; Yoshida, D.; Kanamoto, T.; Nakashima, H.; Uryu, T.; Yoshida, T. Sulfated oligosaccharide cluster with polylysine core scaffold as a new anti-HIV dendrimer. Carbohydr. Polym. 2010, 80, 1111–1115. [Google Scholar] [CrossRef]
- Han, S.; Kanamoto, T.; Nakashima, H.; Yoshida, T. Synthesis of a new amphiphilic glycodendrimer with antiviral functionality. Carbohydr. Polym. 2012, 90, 1061–1068. [Google Scholar] [CrossRef]
- Rosa Borges, A.; Wieczorek, L.; Johnson, B.; Benesi, A.J.; Brown, B.K.; Kensinger, R.D.; Krebs, F.C.; Wigdahl, B.; Blumenthal, R.; Puri, A. Multivalent dendrimeric compounds containing carbohydrates expressed on immune cells inhibit infection by primary isolates of HIV-1. Virology 2010, 408, 80–88. [Google Scholar] [CrossRef]
- Doménech, R.; Abian, O.; Bocanegra, R.; Correa, J.; Sousa-Herves, A.; Riguera, R.; Mateu, M.G.; Fernandez-Megia, E.; Velázquez-Campoy, A. n.; Neira, J.L. Dendrimers as potential inhibitors of the dimerization of the capsid protein of HIV-1. Biomacromolecules 2010, 11, 2069–2078. [Google Scholar] [CrossRef]
- Zhao, H.; Li, J.; Xi, F.; Jiang, L. Polyamidoamine dendrimers inhibit binding of Tat peptide to TAR RNA. FEBS Lett. 2004, 563, 241–245. [Google Scholar] [CrossRef]
- Wang, W.; Guo, Z.; Chen, Y.; Liu, T.; Jiang, L. Influence of Generation 2–5 of PAMAM Dendrimer on the Inhibition of Tat Peptide/TAR RNA Binding in HIV-1 Transcription. Chem. Biol. Drug. Des. 2006, 68, 314–318. [Google Scholar] [CrossRef]
- Wyatt, R.; Sodroski, J. The HIV-1 envelope glycoproteins: Fusogens, antigens, and immunogens. Science 1998, 280, 1884–1888. [Google Scholar] [CrossRef]
- McCarthy, T.D.; Karellas, P.; Henderson, S.A.; Giannis, M.; O'Keefe, D.F.; Heery, G.; Paull, J.R.; Matthews, B.R.; Holan, G. Dendrimers as drugs: Discovery and preclinical and clinical development of dendrimer-based microbicides for HIV and STI prevention. Mol. Pharm. 2005, 2, 312–318. [Google Scholar] [CrossRef]
- Bernstein, D.; Stanberry, L.; Sacks, S.; Ayisi, N.; Gong, Y.; Ireland, J.; Mumper, R.; Holan, G.; Matthews, B.; McCarthy, T. Evaluations of unformulated and formulated dendrimer-based microbicide candidates in mouse and guinea pig models of genital herpes. Antimicrob. Agents Chemother. 2003, 47, 3784–3788. [Google Scholar] [CrossRef]
- Dezzutti, C.S.; James, V.N.; Ramos, A.; Sullivan, S.T.; Siddig, A.; Bush, T.J.; Grohskopf, L.A.; Paxton, L.; Subbarao, S.; Hart, C.E. In vitro comparison of topical microbicides for prevention of human immunodeficiency virus type 1 transmission. Antimicrob. Agents Chemother. 2004, 48, 3834–3844. [Google Scholar] [CrossRef]
- Rupp, R.; Rosenthal, S.L.; Stanberry, L.R. VivaGel™(SPL7013 Gel): A candidate dendrimer–microbicide for the prevention of HIV and HSV infection. Int. J. Nanomed. 2007, 2, 561. [Google Scholar]
- Cohen, C.R.; Brown, J.; Moscicki, A.-B.; Bukusi, E.A.; Paull, J.R.; Price, C.F.; Shiboski, S. A phase I randomized placebo controlled trial of the safety of 3% SPL7013 Gel (VivaGel®) in healthy young women administered twice daily for 14 days. PLoS One 2011, 6, e16258. [Google Scholar]
- Price, C.F.; Tyssen, D.; Sonza, S.; Davie, A.; Evans, S.; Lewis, G.R.; Xia, S.; Spelman, T.; Hodsman, P.; Moench, T.R. SPL7013 Gel (VivaGel®) Retains Potent HIV-1 and HSV-2 Inhibitory Activity following Vaginal Administration in Humans. PLoS One 2011, 6, e24095. [Google Scholar] [CrossRef]
- Harouse, J.M.; Collman, R.G.; Gonzalez-Scarano, F. Human immunodeficiency virus type 1 infection of SK-N-MC cells: domains of gp120 involved in entry into a CD4-negative, galactosyl ceramide/3'sulfo-galactosyl ceramide-positive cell line. J. Virol. 1995, 69, 7383–7390. [Google Scholar]
- Yahi, N.; Baghdiguian, S.; Moreau, H.; Fantini, J. Galactosyl ceramide (or a closely related molecule) is the receptor for human immunodeficiency virus type 1 on human colon epithelial HT29 cells. J. Virol. 1992, 66, 4848–4854. [Google Scholar]
- Popik, W.; Alce, T.M.; Au, W.-C. Human immunodeficiency virus type 1 uses lipid raft-colocalized CD4 and chemokine receptors for productive entry into CD4+ T cells. J. Virol. 2002, 76, 4709–4722. [Google Scholar] [CrossRef]
- Bhat, S.; Spitalnik, S.L.; Gonzalez-Scarano, F.; Silberberg, D.H. Galactosyl ceramide or a derivative is an essential component of the neural receptor for human immunodeficiency virus type 1 envelope glycoprotein gp120. Proc. Natl. Acad. Sci. USA 1991, 88, 7131–7134. [Google Scholar] [CrossRef]
- Pérez-Anes, A.; Stefaniu, C.; Moog, C.; Majoral, J.-P.; Blanzat, M.; Turrin, C.-O.; Caminade, A.-M.; Rico-Lattes, I. Multivalent catanionic GalCer analogs derived from first generation dendrimeric phosphonic acids. Bioorg. Med. Chem. 2010, 18, 242–248. [Google Scholar] [CrossRef]
- Lundquist, J.J. The cluster glycoside effect. Chem Rev. 2002, 102, 555–578. [Google Scholar] [CrossRef]
- Katsuraya, K.; Ikushima, N.; Takahashi, N.; Shoji, T.; Nakashima, H.; Yamamoto, N.; Yoshida, T.; Uryu, T. Synthesis of sulfated alkyl malto-and laminara-oligosaccharides with potent inhibitory effects on AIDS virus infection. Carbohydr. Res. 1994, 260, 51–61. [Google Scholar] [CrossRef]
- Hug, P.; Lin, H.-M.J.; Korte, T.; Xiao, X.; Dimitrov, D.S.; Wang, J.M.; Puri, A.; Blumenthal, R. Glycosphingolipids promote entry of a broad range of human immunodeficiency virus type 1 isolates into cell lines expressing CD4, CXCR4, and/or CCR5. J. Virol. 2000, 74, 6377–6385. [Google Scholar]
- Puri, A.; Hug, P.; Jernigan, K.; Barchi, J.; Kim, H.-Y.; Hamilton, J.; Wiels, J.; Murray, G.J.; Brady, R.O.; Blumenthal, R. The neutral glycosphingolipid globotriaosylceramide promotes fusion mediated by a CD4-dependent CXCR4-utilizing HIV type 1 envelope glycoprotein. Proc. Natl. Acad. Sci. USA 1998, 95, 14435–14440. [Google Scholar] [CrossRef]
- Puri, A.; Hug, P.; Jernigan, K.; Rose, P.; Blumenthal, R. Role of glycosphingolipids in HIV-1 entry: requirement of globotriosylceramide (Gb3) in CD4/CXCR4-dependent fusion. Biosci. Rep. 1999, 19, 317–325. [Google Scholar] [CrossRef]
- Geijtenbeek, T.B.; van Kooyk, Y. Pathogens target DC-SIGN to influence their fate DC-SIGN functions as a pathogen receptor with broad specificity. APMIS 2003, 111, 698–714. [Google Scholar] [CrossRef]
- Kooyk, Y.V.; Appelmelk, B.; Geijtenbeek, T.B. A fatal attraction:Mycobacterium tuberculosis and HIV-1 target DC-SIGN to escape immune surveillance. Trends Mol. Med. 2003, 9, 153–159. [Google Scholar] [CrossRef]
- Lozach, P.-Y.; Amara, A.; Bartosch, B.; Virelizier, J.-L.; Arenzana-Seisdedos, F.; Cosset, F.-L.; Altmeyer, R. C-type lectins L-SIGN and DC-SIGN capture and transmit infectious hepatitis C virus pseudotype particles. J. Biol. Chem. 2004, 279, 32035–32045. [Google Scholar] [CrossRef]
- Doores, K.J.; Bonomelli, C.; Harvey, D.J.; Vasiljevic, S.; Dwek, R.A.; Burton, D.R.; Crispin, M.; Scanlan, C.N. Envelope glycans of immunodeficiency virions are almost entirely oligomannose antigens. Proc. Natl. Acad. Sci. USA 2010, 107, 13800–13805. [Google Scholar] [CrossRef]
- Christopher, G. Selection of a synthetic glycan oligomer from a library of DNA-templated fragments against DC-SIGN and inhibition of HIV gp120 binding to dendritic cells. Chem. Commun. (Camb.) 2011, 47, 9321–9323. [Google Scholar] [CrossRef]
- Hamy, F.; Brondani, V.; Flörsheimer, A.; Stark, W.; Blommers, M.J.; Klimkait, T. A new class of HIV-1 Tat antagonist acting through Tat-TAR inhibition. Biochemistry 1998, 37, 5086–5095. [Google Scholar]
- Dayton, A.I.; Sodroski, J.G.; Rosen, C.A.; Goh, W.C.; Haseltine, W.A. The trans-activator gene of the human T cell lymphotropic virus type III is required for replication. Cell 1986, 44, 941–947. [Google Scholar] [CrossRef]
- Zhao, H.; Dai, D.; Li, J.; Chen, Y.; Jiang, L. Quantitative study of HIV-1 Tat peptide and TAR RNA interaction inhibited by poly (allylamine hydrochloride). Biochem. Biophys. Res. Commun. 2003, 312, 351–354. [Google Scholar] [CrossRef]
- Asaftei, S.; De Clercq, E. “Viologen” Dendrimers as Antiviral Agents: The effect of charge number and distance. J. Med. Chem. 2010, 53, 3480–3488. [Google Scholar] [CrossRef]
- McGann, K.A.; Collman, R.; Kolson, D.L.; Gonzalez-Scarano, F.; Coukos, G.; Coutifaris, C.; Strauss, J.F.; Nathanson, N. Human immunodeficiency virus type 1 causes productive infection of macrophages in primary placental cell cultures. J. Infect. Dis. 1994, 169, 746–753. [Google Scholar] [CrossRef]
- Auger, M.; Ross, J.; Lewis, C.; McGee, J. The Natural Immune System: The Macrophage; Oxford University Press: New York, NY, USA, 1992. [Google Scholar]
- Gordon, S. Mononuclear phagocytes in immune defence. Immunology 2001, 6, 147–162. [Google Scholar]
- Lee, Y.C.; Townsend, R.; Hardy, M.R.; Lönngren, J.; Arnarp, J.; Haraldsson, M.; Lönn, H. Binding of synthetic oligosaccharides to the hepatic Gal/GalNAc lectin. Dependence on fine structural features. J. Biol. Chem. 1983, 258, 199–202. [Google Scholar]
- Bhadra, D.; Yadav, A.; Bhadra, S.; Jain, N. Glycodendrimeric nanoparticulate carriers of primaquine phosphate for liver targeting. Int. J. Pharm. 2005, 295, 221–233. [Google Scholar] [CrossRef]
- Dutta, T.; Agashe, H.B.; Garg, M.; Balasubramanium, P.; Kabra, M.; Jain, N.K. Poly (propyleneimine) dendrimer based nanocontainers for targeting of efavirenz to human monocytes/macrophages in vitro: Research Paper. J. Drug Target. 2007, 15, 89–98. [Google Scholar] [CrossRef]
- Dutta, T.; Jain, N.K. Targeting potential and anti-HIV activity of lamivudine loaded mannosylated poly (propyleneimine) dendrimer. Biochim. Biophys. Acta. 2007, 1770, 681–686. [Google Scholar]
- Vinogradov, S.V.; Poluektova, L.Y.; Makarov, E.; Gerson, T.; Senanayake, M.T. Nano-NRTIs: efficient inhibitors of HIV type-1 in macrophages with a reduced mitochondrial toxicity. Antivir. Chem. Chemother. 2010, 21, 1. [Google Scholar] [CrossRef]
- Buckley, D.L.; Corson, T.W.; Aberle, N.; Crews, C.M. HIV protease-mediated activation of sterically capped proteasome inhibitors and substrates. J. Am. Chem. Soc. 2010, 133, 698–700. [Google Scholar]
- Pion, M.; Serramia, M.J.; Diaz, L.; Bryszewska, M.; Gallart, T.; García, F.; Gómez, R.; de la Mata, F.J.; Muñoz-Fernandez, M.Á. Phenotype and functional analysis of human monocytes-derived dendritic cells loaded with a carbosilane dendrimer. Biomaterials 2010, 31, 8749–8758. [Google Scholar] [CrossRef]
- Ionov, M.; Ciepluch, K.; Klajnert, B.; Glińska, S.; Gomez-Ramirez, R.; de la Mata, F.J.; Munoz-Fernandez, M.A.; Bryszewska, M. Complexation of HIV derived peptides with carbosilane dendrimers. Colloids Surf. B Biointerfaces. 2012, 101, 236–242. [Google Scholar]
- Rinaldo, C. Dendritic cell-based human immunodeficiency virus vaccine. J. Intern. Med. 2009, 265, 138–158. [Google Scholar] [CrossRef]
- Rayburn, E.; Wang, W.; Zhang, R.; Wang, H. Antisense approaches in drug discovery and development. In Prog Drug Res.; Springer: Basel, Swizerland, 2005. [Google Scholar]
- Chonco, L.; Bermejo-Martín, J.F.; Ortega, P.; Shcharbin, D.; Pedziwiatr, E.; Klajnert, B.; de la Mata, F.J.; Eritja, R.; Gómez, R.; Bryszewska, M. Water-soluble carbosilane dendrimers protect phosphorothioate oligonucleotides from binding to serum proteins. Org. Biomol. Chem. 2007, 5, 1886–1893. [Google Scholar] [CrossRef] [Green Version]
- Gonzalo, T.; Clemente, M.I.; Chonco, L.; Weber, N.D.; Díaz, L.; Serramía, M.J.; Gras, R.; Ortega, P.; de la Mata, F.J.; Gómez, R. Gene Therapy in HIV-Infected Cells to Decrease Viral Impact by Using an Alternative Delivery Method. ChemMedChem. 2010, 5, 921–929. [Google Scholar] [CrossRef]
- De Las Cuevas, N.; Garcia-Gallego, S.; Rasines, B.; de la Mata, F.; G Guijarro, L.; Munoz-Fernandez, M.; Gomez, R. In Vitro Studies of Water-Stable Cationic Carbosilane Dendrimers As Delivery Vehicles for Gene Therapy Against HIV and Hepatocarcinoma. Curr Med. Chem. 2012, 19, 5052–5061. [Google Scholar] [CrossRef]
- Pedziwiatr-Werbicka, E.; Shcharbin, D.; Maly, J.; Maly, M.; Zaborski, M.; Gabara, B.; Ortega, P.; de la Mata, F.J.; Gomez, R.; Munoz-Fernandez, M.A. Carbosilane Dendrimers are a Non-Viral Delivery System for Antisense Oligonucleotides: Characterization of Dendriplexes. J. Biomed. Nanotechnol. 2012, 8, 57–73. [Google Scholar]
- Shcharbin, D.; Pedziwiatr, E.; Nowacka, O.; Kumar, M.; Zaborski, M.; Ortega, P.; Javier de la Mata, F.; Gómez, R.; Muñoz-Fernandez, M.A.; Bryszewska, M. Carbosilane dendrimers NN8 and NN16 form a stable complex with siGAG1. Colloids Surf. B Biointerfaces. 2011, 83, 388–391. [Google Scholar] [CrossRef]
- Weber, N.; Ortega, P.; Clemente, M.I.; Shcharbin, D.; Bryszewska, M.; de la Mata, F.J.; Gómez, R.; Muñoz-Fernández, M. Characterization of carbosilane dendrimers as effective carriers of siRNA to HIV-infected lymphocytes. J. Control Release 2008, 132, 55–64. [Google Scholar] [CrossRef]
- Jiménez, J.L.; Clemente, M.I.; Weber, N.D.; Sanchez, J.; Ortega, P.; de la Mata, F.J.; Gómez, R.; García, D.; López-Fernández, L.A.; Muñoz-Fernández, M.Á. Carbosilane dendrimers to transfect human astrocytes with small interfering RNA targeting human immunodeficiency virus. BioDrugs 2010, 24, 331–343. [Google Scholar] [CrossRef]
- Szewczyk, M.; Drzewinska, J.; Dzmitruk, V.; Shcharbin, D.; Klajnert, B.; Appelhans, D.; Bryszewska, M. Stability of Dendriplexes Formed by Anti-HIV Genetic Material and Poly (propylene imine) Dendrimers in the Presence of Glucosaminoglycans. J. Phys. Chem. B 2012, 116, 14525–14532. [Google Scholar] [CrossRef]
- Zhou, J.; Neff, C.P.; Liu, X.; Zhang, J.; Li, H.; Smith, D.D.; Swiderski, P.; Aboellail, T.; Huang, Y.; Du, Q. Systemic administration of combinatorial dsiRNAs via nanoparticles efficiently suppresses HIV-1 infection in humanized mice. Mol. Ther. 2011, 19, 2228–2238. [Google Scholar] [CrossRef]
- Briz, V.; Serramia, M.; Madrid, R.; Hameau, A.; Caminade, A.-M.; Majoral, J.; Munoz-Fernandez, M. Validation of a Generation 4 Phosphorus-Containing Polycationic Dendrimer for Gene Delivery Against HIV-1. Curr. Med. Chem. 2012, 19, 5044–5051. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are not available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Peng, J.; Wu, Z.; Qi, X.; Chen, Y.; Li, X. Dendrimers as Potential Therapeutic Tools in HIV Inhibition. Molecules 2013, 18, 7912-7929. https://doi.org/10.3390/molecules18077912
Peng J, Wu Z, Qi X, Chen Y, Li X. Dendrimers as Potential Therapeutic Tools in HIV Inhibition. Molecules. 2013; 18(7):7912-7929. https://doi.org/10.3390/molecules18077912
Chicago/Turabian StylePeng, Jianqing, Zhenghong Wu, Xiaole Qi, Yi Chen, and Xiangbo Li. 2013. "Dendrimers as Potential Therapeutic Tools in HIV Inhibition" Molecules 18, no. 7: 7912-7929. https://doi.org/10.3390/molecules18077912