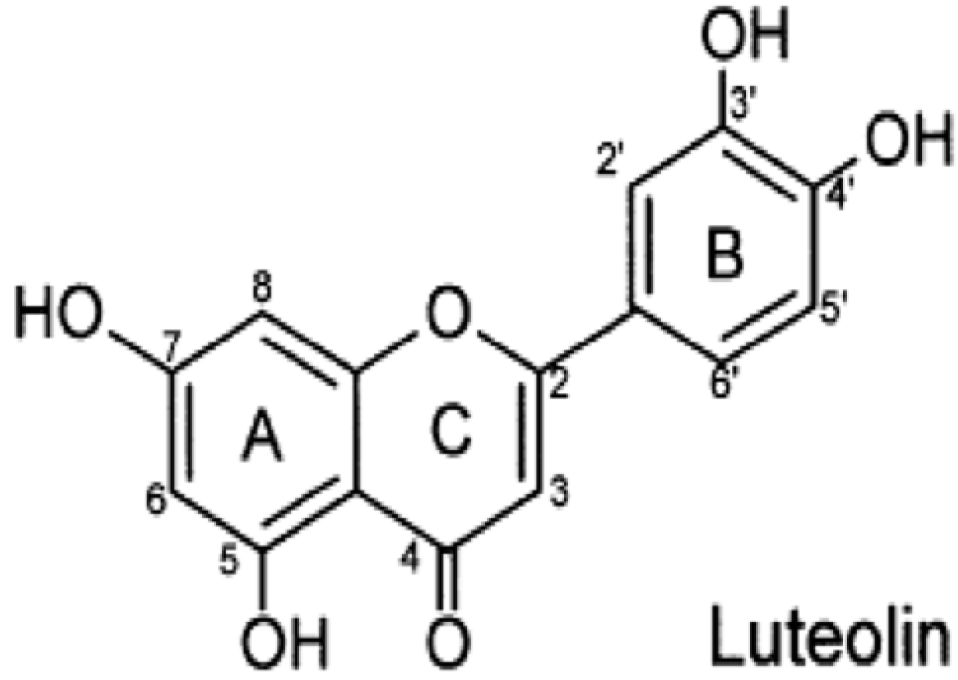
Luteolin Inhibits Inflammatory Responses via p38/MK2/TTP-mediated mRNA Stability

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
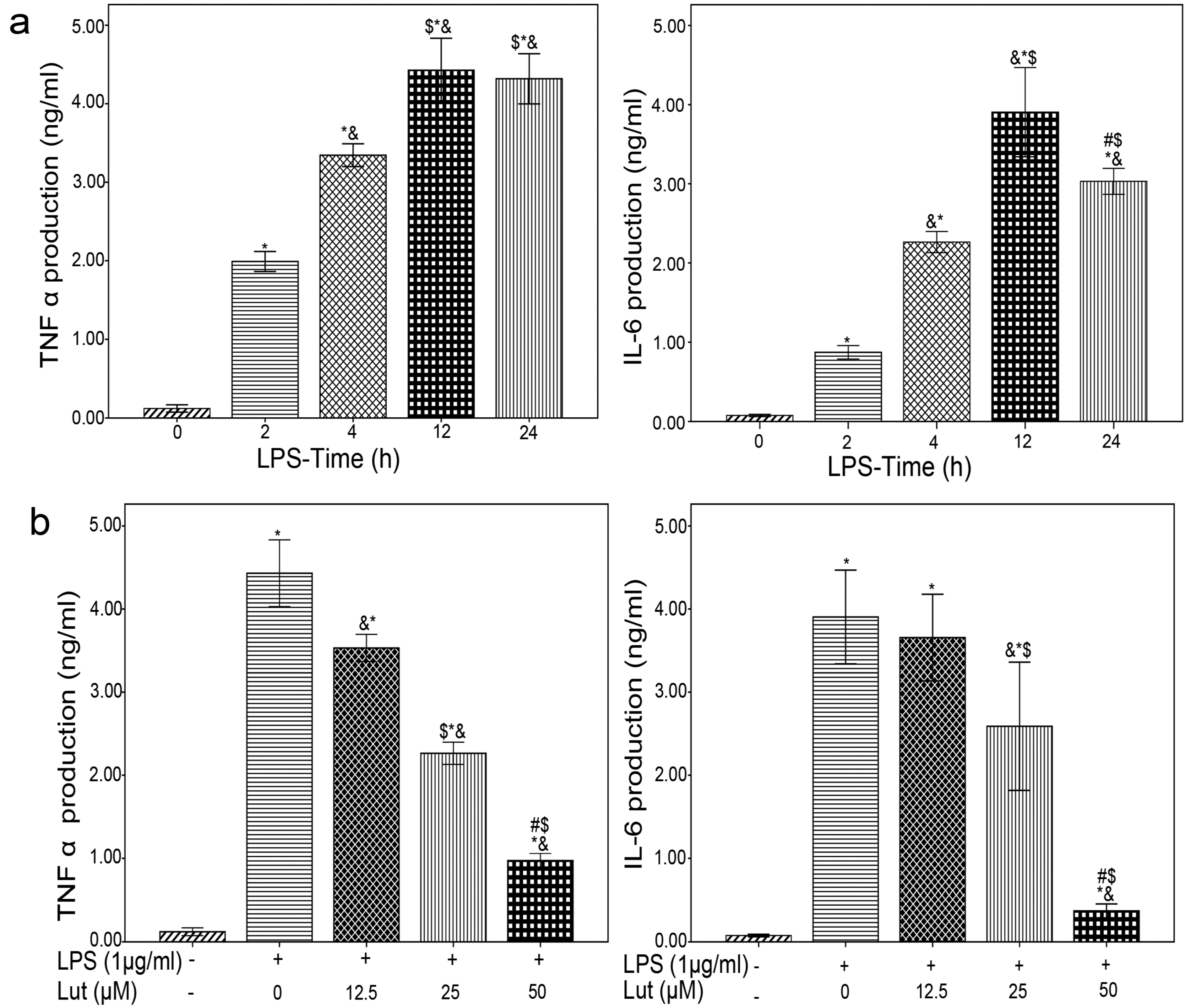
2.1. Lut Inhibits the Production of TNF-α and IL-6


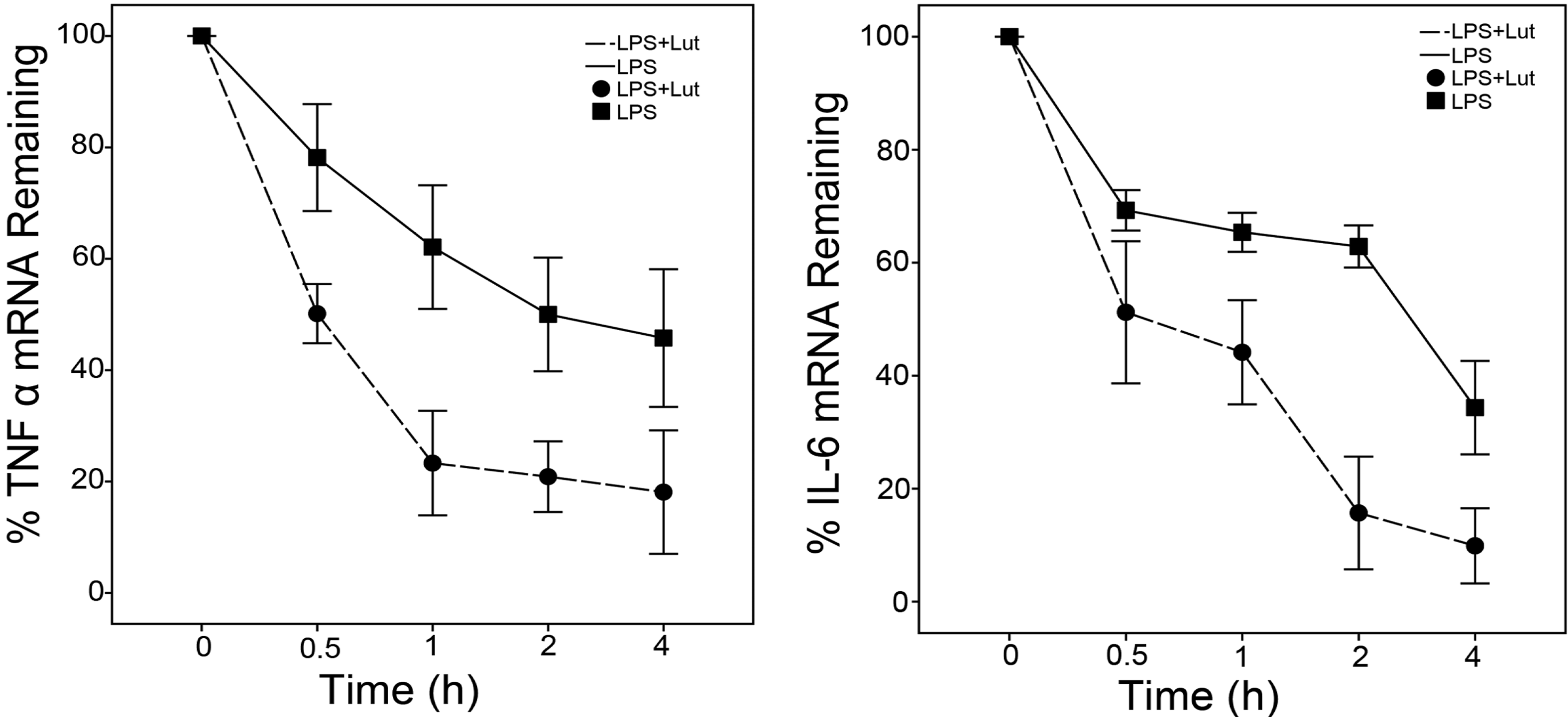
2.2. Lut Promotes the Degradation of TNF-α and IL-6 mRNAs

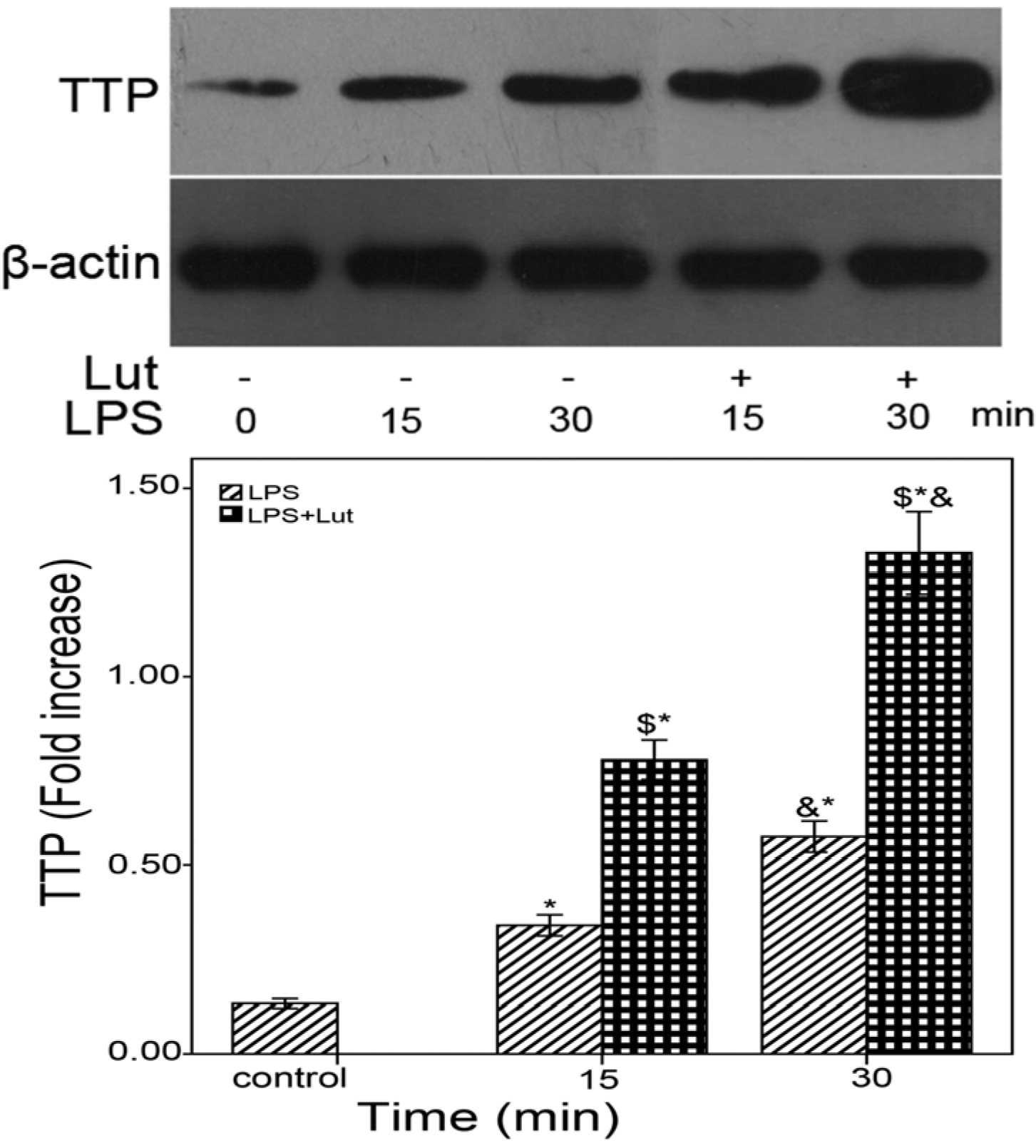
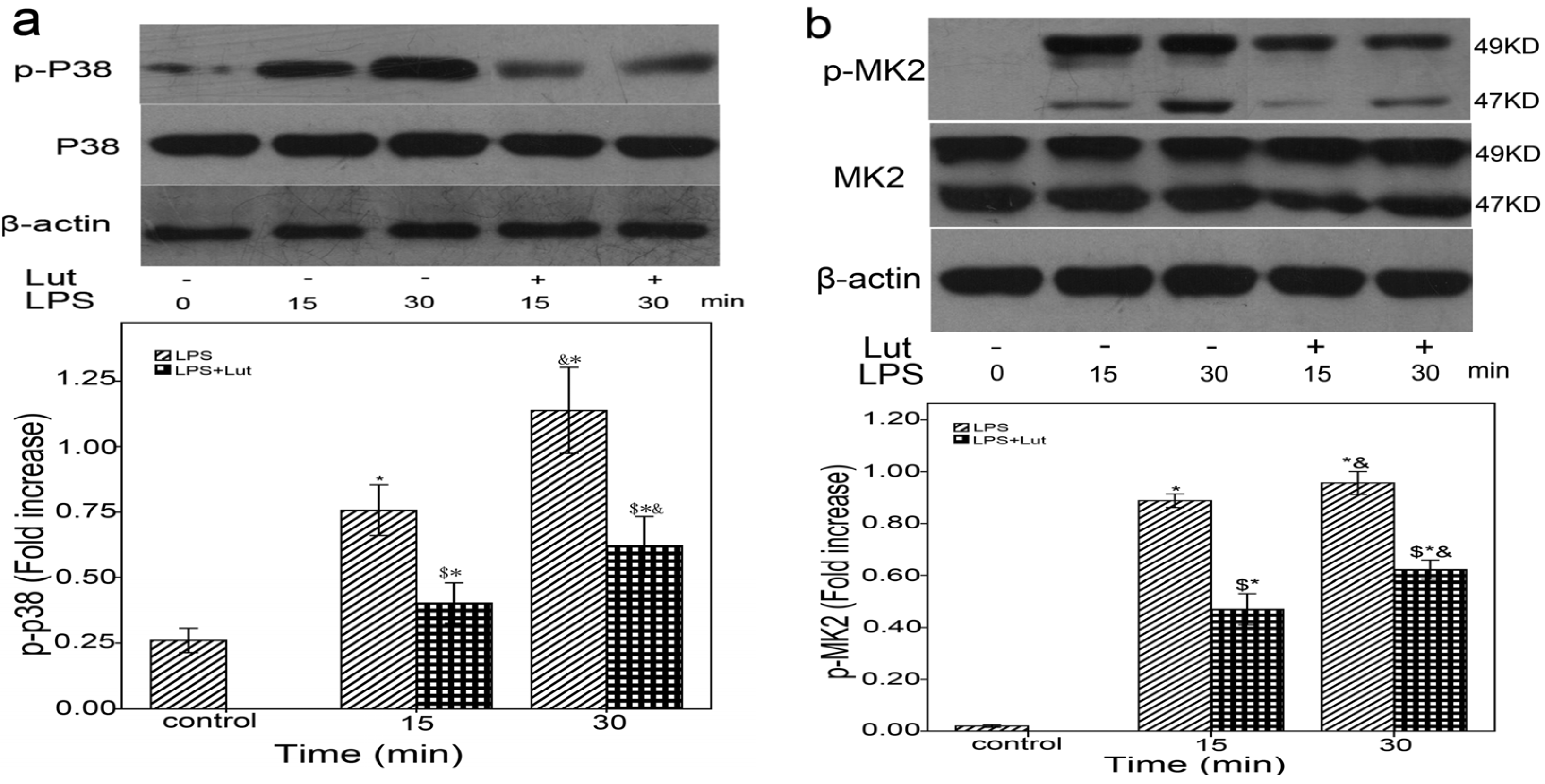
2.3. Lut Affects the mRNAs Stability of TNF-α and IL-6 via the p38/MK2/TTP Signaling Cascade


3. Experimental
3.1. Materials
3.2. BMM Cultures
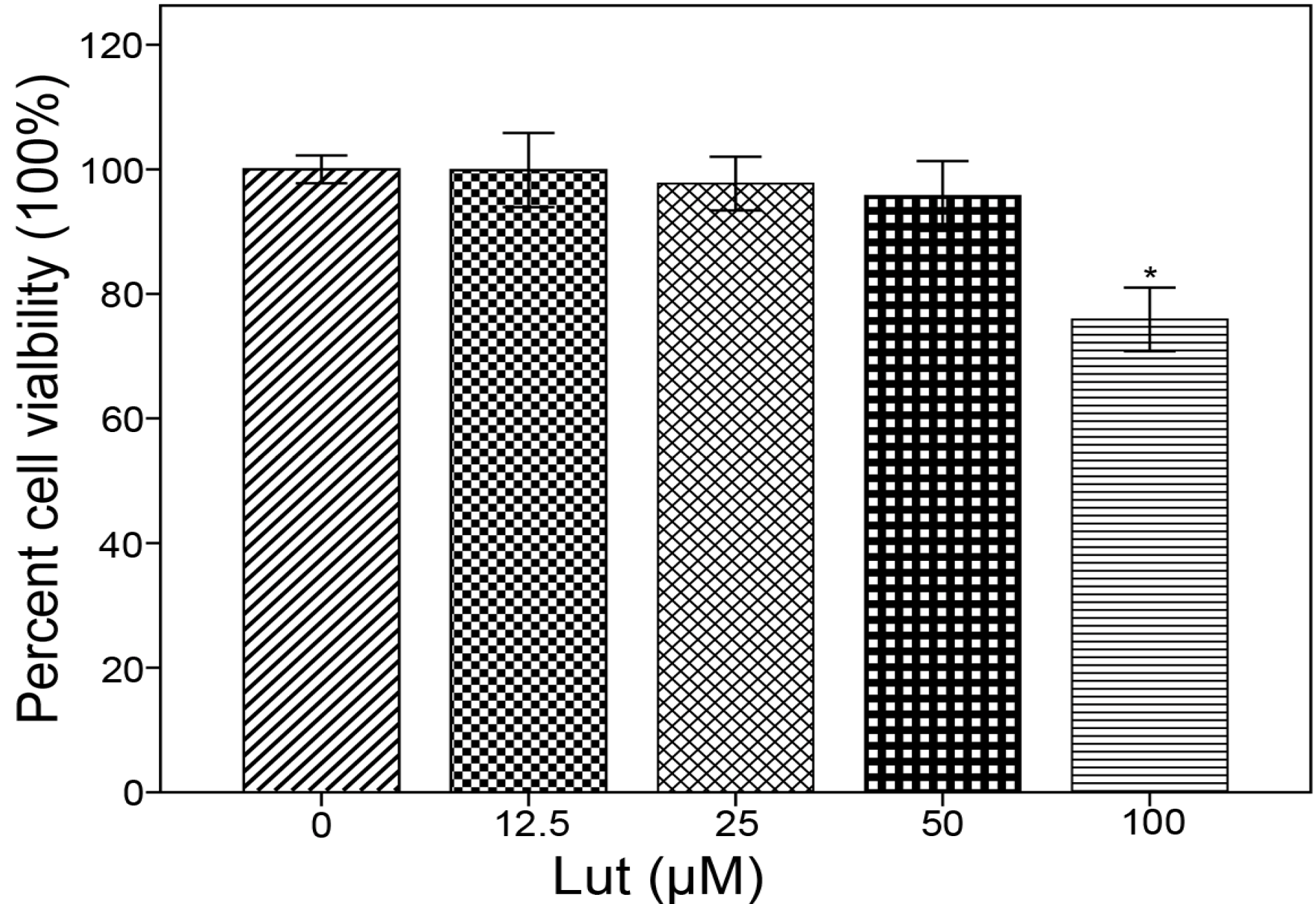
3.3. MTT Assay
3.4. ELISA
3.5. RNA Isolation and Reverse Transcription
3.6. Real-Time PCR
3.7. Western Blot
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- McCullough, M.L.; Peterson, J.J.; Patel, R.; Jacques, P.F.; Shah, R.; Dwyer, J.T. Flavonoid intake and cardiovascular disease mortality in a prospective cohort of US adults. Am. J. Clin. Nutr. 2012, 95, 454–464. [Google Scholar]
- Neuhouser, M.L. Dietary flavonoids and cancer risk: Evidence from human population studies. Nutr. Cancer 2004, 50, 1–7. [Google Scholar] [CrossRef]
- Kumazawa, Y.; Kawaguchi, K.; Takimoto, H. Immunomodulating effects of flavonoids on acute and chronic inflammatory responses caused by tumor necrosis factor alpha. Curr. Pharm. Des. 2006, 12, 4271–4279. [Google Scholar]
- Harris, G.K.; Qian, Y.; Leonard, S.S.; Sbarra, D.C; Shi, X. Luteolin and chrysin differentially inhibit cyclooxygenase-2 expression and scavenge reactive oxygen species but similarly inhibit prostaglandin-E2 formation in RAW 264.7 cells. J. Nutr. 2006, 136, 1517–1521. [Google Scholar]
- Park, C.M.; Jin, K.S; Lee, Y.W.; Song, Y.S. Luteolin and chicoric acid synergistically inhibited inflammatory responses via inactivation of PI3K-Akt pathway and impairment of NF-kappaB translocation in LPS stimulated RAW 264.7 cells. Eur. J. Pharmacol. 2011, 660, 454–459. [Google Scholar]
- Lang, Y.; Chen, D; Li, D.Y.; Zhu, M.Y.; Xu, T.D.; Zhang, T.; Qian, W.H.; Luo, Y.Y. Luteolin inhibited hydrogen peroxide-induced vascular smooth muscle cells proliferation and migration by suppressing the Src and Akt signalling pathways. J. Pharm. Pharmacol. 2012, 64, 597–603. [Google Scholar]
- Zhu, M.Y.; Chen, D.; Li, D.Y.; Ding, H.; Zhang, T.; Xu, T.D.; Zhang, Y.B. Luteolin inhibits angiotensin II-induced human umbilical vein endothelial cell proliferation and migration through downregulation of Src and Akt phosphorylation. Circ. J. 2013, 77, 772–779. [Google Scholar]
- Libby, P. Inflammation in atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar]
- Szekanecz, Z. Pro-inflammatory cytokines in atherosclerosis. Isr. Med. Assoc. J. 2008, 10, 529–530. [Google Scholar]
- Gotsman, I.; Stabholz, A.; Planer, D.; Pugatsch, T.; Lapidus, L.; Novikov, Y.; Masrawa, S.; Soskolne, A.; Lotan, C. Serum cytokine tumor necrosis factor-alpha and interleukin-6 associated with the severity of coronary artery disease: indicators of an active inflammatory burden? Isr. Med. Assoc. J. 2008, 10, 494–498. [Google Scholar]
- Wu, X.; Brewer, G. The regulation of mRNA stability in mammalian cells: 2.0. Gene 2012, 500, 10–21. [Google Scholar] [CrossRef]
- Winzen, R.; Gowrishankar, G.; Bollig, F.; Redich, N.; Resch, K.; Holtmann, H. Distinct domains of AU-rich elements exert different functions in mRNA destabilization and stabilization by p38 mitogen-activated protein kinase or HuR. Mol. Cell. Biol. 2004, 24, 4835–4847. [Google Scholar] [CrossRef]
- Chen, C.Y.; Shyu, A.B. AU-rich elements: Characterization and importance in mRNA degradation. Trends Biochem. Sci. 1995, 20, 465–470. [Google Scholar] [CrossRef]
- Shaw, G.; Kamen, R. A conserved AU sequence from the 3' untranslated region of GM-CSF mRNA mediates selective mRNA degradation. Cell 1986, 46, 659–667. [Google Scholar]
- Biswas, R.; Biswas, R.; Datta, S.; Gupta, J.D.; Novotny, M.; Tebo, J.; Hamilton, T.A. Regulation of chemokine mRNA stability by lipopolysaccharide and IL-10. J. Immunol. 2003, 170, 6202–6208. [Google Scholar]
- Carrick, D.M.; Lai, W.S.; Blackshear, P.J. The tandem CCCH zinc finger protein tristetraprolin and its relevance to cytokine mRNA turnover and arthritis. Arthritis Res. Ther. 2004, 6, 248–264. [Google Scholar] [CrossRef] [Green Version]
- Wilson, G.M.; Brewer, G. The search for trans-acting factors controlling messenger RNA decay. Prog. Nucleic. Acid Res. Mol. Biol. 1999, 62, 257–291. [Google Scholar]
- Dean, J.L.; Sully, G.; Clark, A.R.; Saklatvala, J. The involvement of AU-rich element-binding proteins in p38 mitogen-activated protein kinase pathway-mediated mRNA stabilisation. Cell Signal 2004, 16, 1113–1121. [Google Scholar]
- Brennan, C.M.; Steitz, J.A. HuR and mRNA stability. Cell Mol. Life Sci. 2001, 58, 266–277. [Google Scholar] [CrossRef]
- Shi, J.X.; Su, X.; Xu, J.; Zhang, W.Y.; Shi, Y. MK2 posttranscriptionally regulates TNF-alpha-induced expression of ICAM-1 and IL-8 via tristetraprolin in human pulmonary microvascular endothelial cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 302, 793–799. [Google Scholar] [CrossRef]
- Lai, W.S.; Carballo, E.; Strum, J.R.; Kennington, E.A.; Phillips, R.S.; Blackshear, P.J. Evidence that tristetraprolin binds to AU-rich elements and promotes the deadenylation and destabili-zation of tumor necrosis factor alpha mRNA. Mol. Cell Biol. 1999, 19, 4311–4323. [Google Scholar]
- Chen, C.Y.; Gherzi, R.; Ong, S.E.; Chan, E.L.; Raijmakers, R.; Pruijn, G.J.; Stoecklin, G.; Moroni, C.; Mann, M.; Karin, M. AU binding proteins recruit the exosome to degrade ARE-containing mRNAs. Cell 2001, 107, 451–464. [Google Scholar] [CrossRef]
- Mahtani, K.R.; Brook, M.; Dean, J.L.; Sully, G.; Saklatvala, J.; Clark, A.R. Mitogen-activated protein kinase p38 controls the expression and posttranslational modification of tristetraprolin, a regulator of tumor necrosis factor alpha mRNA stability. Mol. Cell. Biol. 2001, 21, 6461–6469. [Google Scholar] [CrossRef]
- Chrestensen, C.A.; Schroeder, M.J.; Shabanowitz, J.; Hunt, D.F.; Pelo, J.W.; Worthington, M.T.; Sturgill, T.W. MAPKAP kinase 2 phosphorylates tristetraprolin on in vivo sites including Ser178, a site required for 14–3-3 binding. J. Biol. Chem. 2004, 279, 10176–10184. [Google Scholar]
- Brook, M.; Tchen, C.R.; Santalucia, T.; McIlrath, J.; Arthur, J.S.; Saklatvala, J.; Clark, A.R. Posttranslational regulation of tristetraprolin subcellular localization and protein stability by p38 mitogen-activated protein kinase and extracellular signal-regulated kinase pathways. Mol. Cell Biol. 2006, 26, 2408–2418. [Google Scholar] [CrossRef]
- Hitti, E.; Brook, M.; Deppenmeier, S.; Gruber, A.D.; Radzioch, D.; Clark, A.R.; Blackshear, P.J.; Kotlyarov, A.; Gaestel, M. Mitogen-activated protein kinase-activated protein kinase 2 regulates tumor necrosis factor mRNA stability and translation mainly by altering tristetraprolin expression, stability, and binding to adenine/uridine-rich element. Mol. Cell Biol. 2006, 26, 2399–2407. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, M.; D'Silva, N.J.; Kirkwood, K.L. Tristetraprolin regulates interleukin-6 expression through p38 MAPK-dependent affinity changes with mRNA 3' untranslated region. J. Interferon Cytokine Res. 2011, 31, 629–637. [Google Scholar]
- Sun, L.; Stoecklin, G.; Van Way, S.; Hinkovska-Galcheva, V.; Guo, R.F.; Anderson, P.; Shanley, T.P. Tristetraprolin (TTP)-14–3-3 complex formation protects TTP from dephosphorylation by protein phosphatase 2a and stabilizes tumor necrosis factor-alpha mRNA. J. Biol. Chem. 2007, 282, 3766–3777. [Google Scholar]
- Ronkina, N.; Menon, M.B.; Schwermann, J.; Tiedje, C.; Hitti, E.; Kotlyarov, A.; Gaestel, M. MAPKAP kinases MK2 and MK3 in inflammation: Complex regulation of TNF biosynthesis via expression and phosphorylation of tristetraprolin. Biochem. Pharmacol. 2010, 80, 1915–1920. [Google Scholar] [CrossRef]
- O’Dea, K.P.; Dokpesi, J.O.; Tatham, K.C.; Wilson, M.R.; Takata, M. Regulation of monocyte subset proinflammatory responses within the lung microvasculature by the p38 MAPK/MK2 pathway. Am. J. Physiol. Lung Cell Mol. Physiol. 2011, 301, 812–821. [Google Scholar] [CrossRef]
- Zhu, L.H.; Bi, W.; Qi, R.B.; Wang, H.D.; Lu, D.X. Luteolin inhibits microglial inflammation and improves neuron survival against inflammation. Int. J. Neurosci. 2011, 121, 329–336. [Google Scholar] [CrossRef]
- Lee, J.K.; Kim, S.Y.; Kim, Y.S; Lee, W.H.; Hwang, D.H.; Lee, J.Y. Suppression of the TRIF-dependent signaling pathway of Toll-like receptors by luteolin. Biochem. Pharmacol. 2009, 77, 1391–1400. [Google Scholar]
- Xie, C.; Kang, J.; Li, Z.; Schauss, A.G.; Badger, T.M.; Nagarajan, S.; Wu, T.; Wu, X. The acai flavonoid velutin is a potent anti-inflammatory agent: blockade of LPS-mediated TNF-alpha and IL-6 production through inhibiting NF-kappaB activation and MAPK pathway. J. Nutr. Biochem. 2012, 23, 1184–1191. [Google Scholar] [CrossRef]
- Anderson, P. Post-transcriptional control of cytokine production. Nat. Immunol. 2008, 9, 353–359. [Google Scholar] [CrossRef]
- Patil, C.S.; Liu, M.; Zhao, W.; Coatney, D.D.; Li, F.; VanTubergen, E.A.; D'Silva, N.J.; Kirkwood, K.L. Targeting mRNA stability arrests inflammatory bone loss. Mol. Ther. 2008, 16, 1657–1664. [Google Scholar] [CrossRef]
- Joe, Y.; Kim, H.J.; Kim, S.; Chung, J.; Ko, M.S.; Lee, W.H.; Chang, K.C.; Park, J.W.; Chung, H.T. Tristetraprolin mediates anti-inflammatory effects of nicotine in lipopolysaccharide-stimulated macrophages. J. Biol. Chem. 2011, 286, 24735–24742. [Google Scholar]
- Yin, K.; Deng, X.; Mo, Z.C.; Zhao, G.J.; Jiang, J.; Cui, L.B.; Tan, C.Z.; Wen, G.B.; Fu, Y.; Tang, C.K. Tristetraprolin-dependent post-transcriptional regulation of inflammatory cytokine mRNA expression by apolipoprotein A-I: role of ATP-binding membrane cassette transporter A1 and signal transducer and activator of transcription 3. J. Biol. Chem. 2011, 286, 13834–13845. [Google Scholar] [CrossRef]
- Ronkina, N.; Kotlyarov, A.; Dittrich-Breiholz, O.; Kracht, M.; Hitti, E.; Milarski, K.; Askew, R.; Marusic, S.; Lin, L.L.; Gaestel, M. The mitogen-activated protein kinase (MAPK)-activated protein kinases MK2 and MK3 cooperate in stimulation of tumor necrosis factor biosynthesis and stabilization of p38 MAPK. Mol. Cell. Biol. 2007, 27, 170–181. [Google Scholar] [CrossRef]
- Clement, S.L.; Scheckel, C.; Stoecklin, G.; Lykke-Andersen, J. Phosphorylation of tristetraprolin by MK2 impairs AU-rich element mRNA decay by preventing deadenylase recruitment. Mol. Cell Biol. 2011, 31, 256–266. [Google Scholar] [CrossRef]
- Sandler, H.; Stoecklin, G. Control of mRNA decay by phosphorylation of tristetraprolin. Biochem. Soc. Trans. 2008, 36, 491–496. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, W.; Li, D.; Zong, Y.; Zhu, H.; Pan, D.; Xu, T.; Wang, T.; Wang, T. Luteolin Inhibits Inflammatory Responses via p38/MK2/TTP-mediated mRNA Stability. Molecules 2013, 18, 8083-8094. https://doi.org/10.3390/molecules18078083
Wu W, Li D, Zong Y, Zhu H, Pan D, Xu T, Wang T, Wang T. Luteolin Inhibits Inflammatory Responses via p38/MK2/TTP-mediated mRNA Stability. Molecules. 2013; 18(7):8083-8094. https://doi.org/10.3390/molecules18078083
Chicago/Turabian StyleWu, Wanling, Dongye Li, Yu Zong, Hong Zhu, Defeng Pan, Tongda Xu, Tao Wang, and Tingting Wang. 2013. "Luteolin Inhibits Inflammatory Responses via p38/MK2/TTP-mediated mRNA Stability" Molecules 18, no. 7: 8083-8094. https://doi.org/10.3390/molecules18078083