Volatile Organic Compounds Obtained by in Vitro Callus Cultivation of Plectranthus ornatus Codd. (Lamiaceae)

Abstract
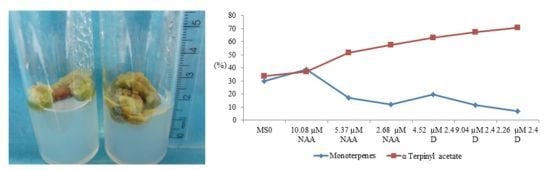
:
1. Introduction
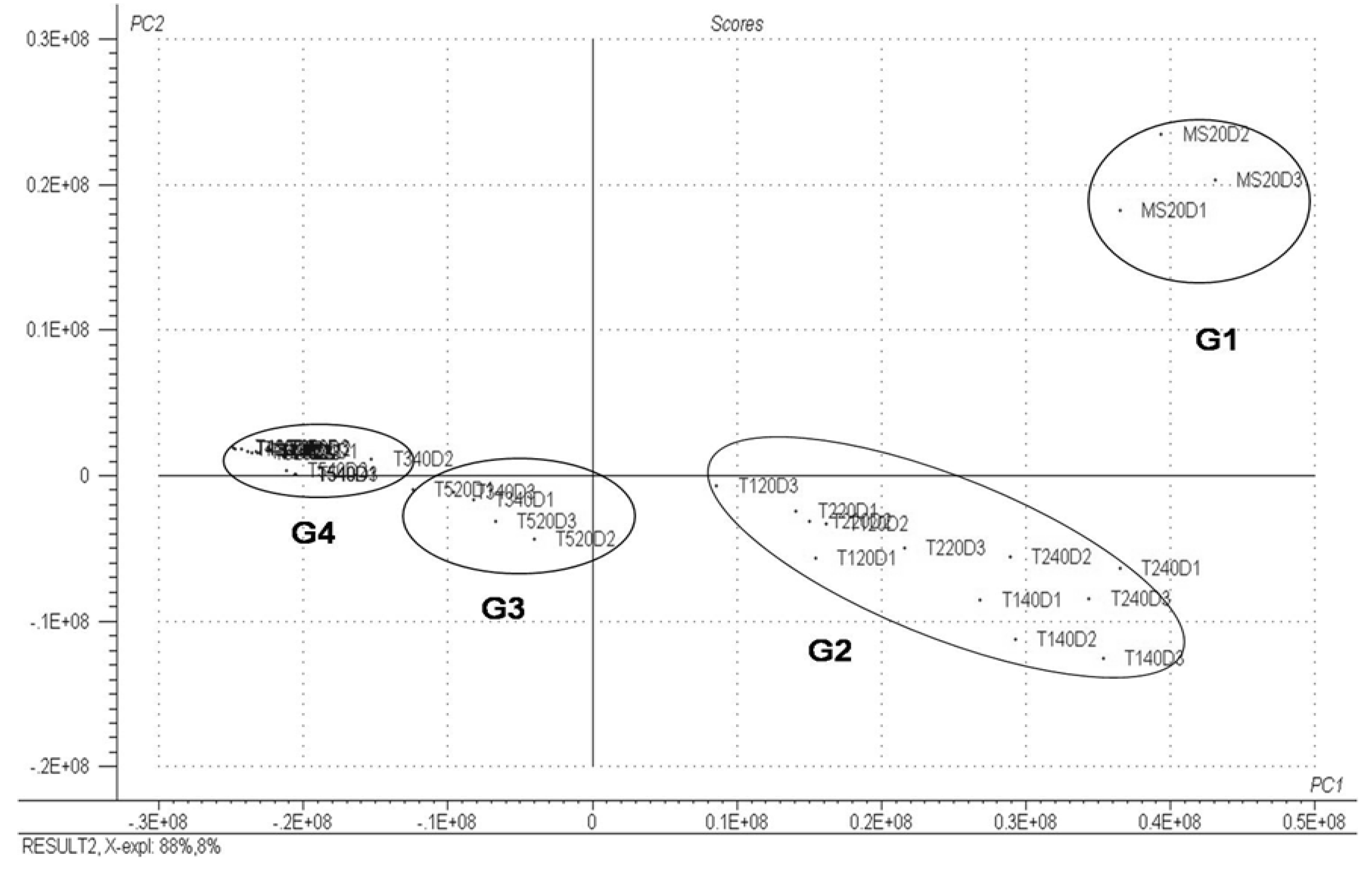
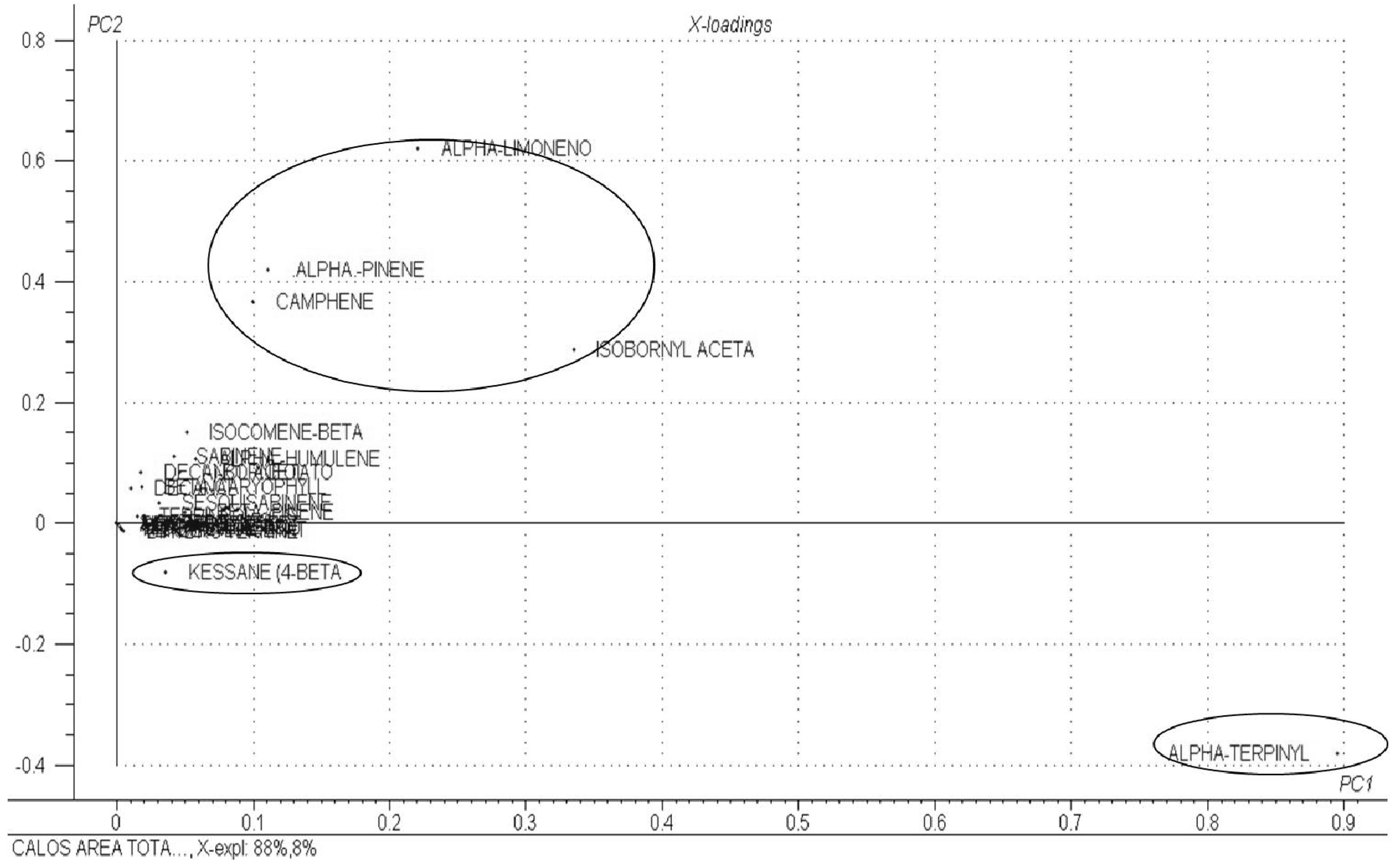
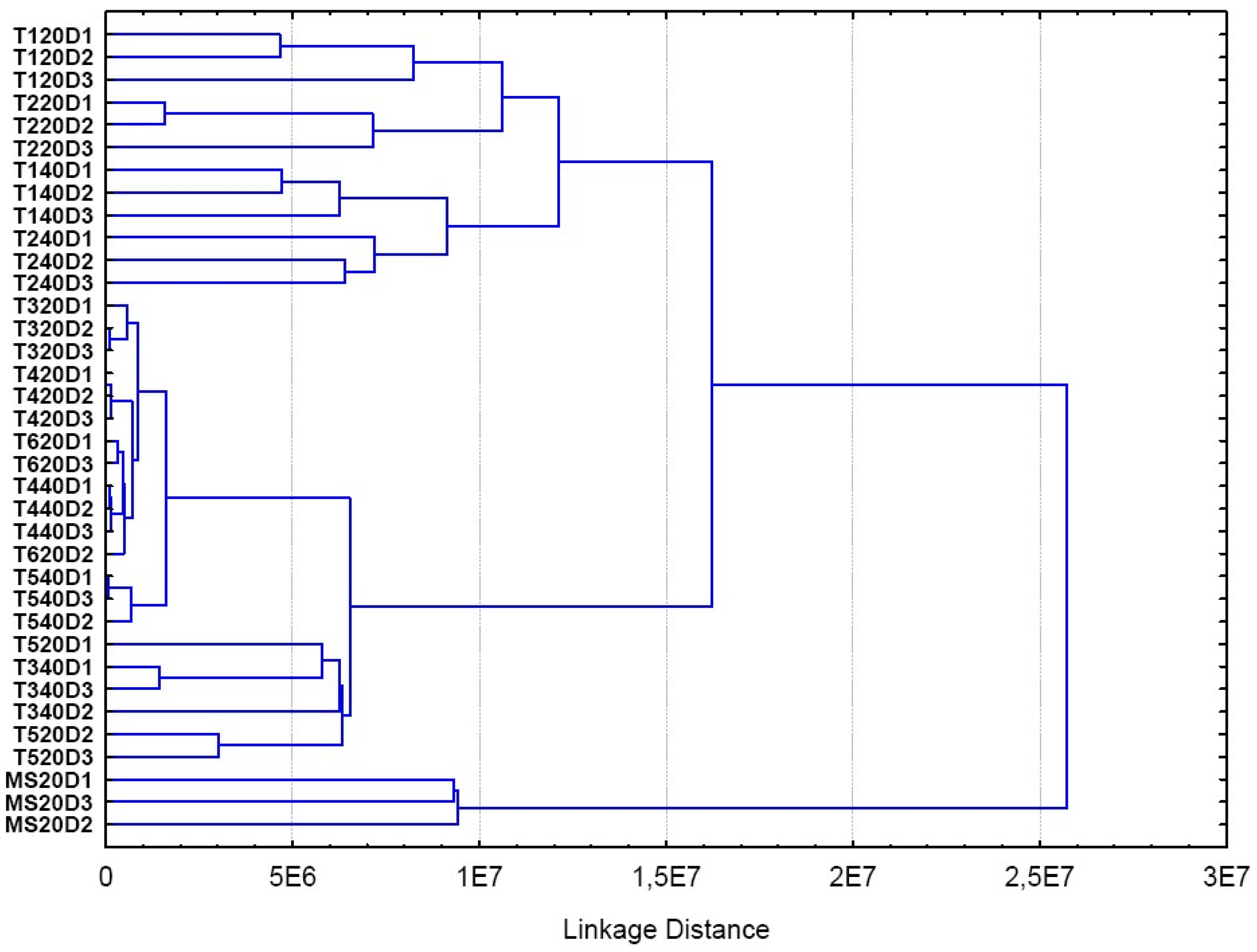
2. Results and Discussion

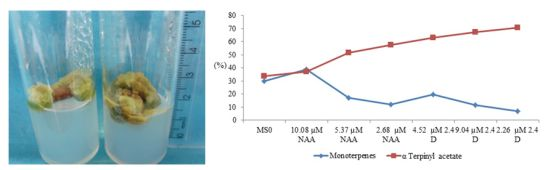{kind=link}
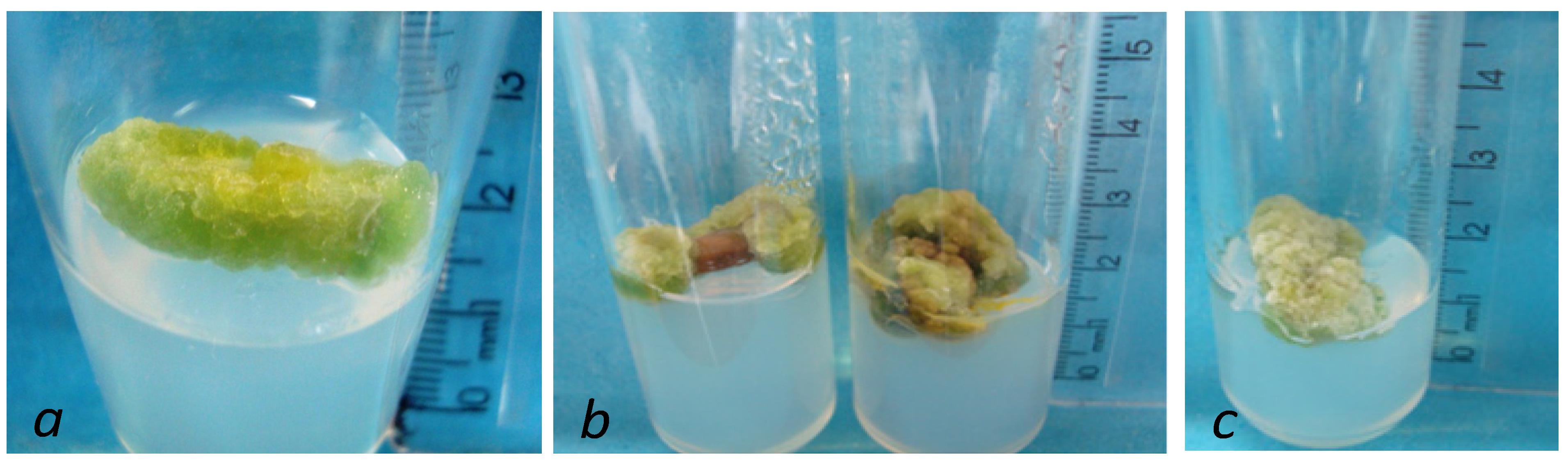{kind=link}
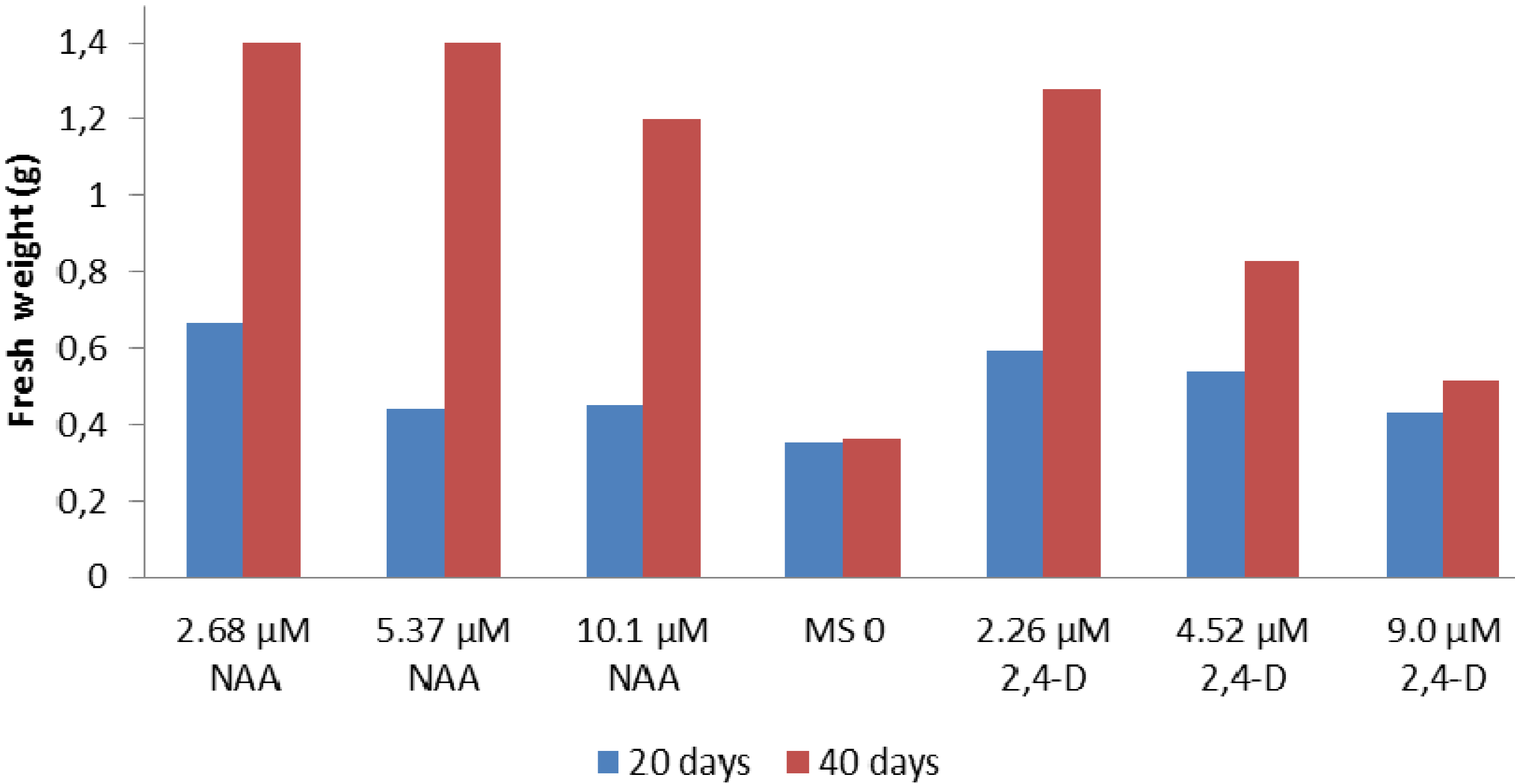{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NAA (µM) | BAP (µM) | |||||
|---|---|---|---|---|---|---|
| 4.5 | 9.0 | 18.0 | ||||
| Original average | Processed average | Original average | Processed average | Original average | Processed average | |
| EB | ||||||
| 0.0 | 90.8 | 77.2aA * | 100.0 | 90.0aA | 100.0 | 90.0aA |
| 5.4 | 100.0 | 90.0aA | 96.4 | 84.2aA | 87.5 | 78.7aA |
| 10.1 | 100.0 | 90.0aA | 100.0 | 90.0aA | 100.0 | 90.0aA |
| 21.5 | 100.0 | 90.0aA | 93.7 | 82.5aA | 100.0 | 90.0aA |
| EA | ||||||
| 0.0 | 30.8 | 32.2abA | 41.5 | 40.6aA | 12.5 | 11.2bA |
| 5.4 | 33.3 | 31.9abA | 50.0 | 45.0aA | 8.3 | 8.7bA |
| 10.1 | 50.0 | 44.9aA | 25.0 | 26.2aA | 34.8 | 36.0aA |
| 21.5 | 32.0 | 34.5aA | 31.2 | 37.5aA | 13.7 | 18.7aA |
| NS | ||||||
| 0.0 | 1.4 | 1.5aAB | 2.2 | 1.7aAB | 1.8 | 1.6aA |
| 5.4 | 1.3 | 1.5aB | 3.1 | 2.0aA | 1.3 | 1.5aA |
| 10.1 | 3.1 | 1.9aA | 1.0 | 1.4bB | 0.9 | 1.3bB |
| 21.5 | 0.8 | 1.3aB | 1.2 | 1.4aB | 0.9 | 1.3aB |
| NL | ||||||
| 0.0 | 5.2 | 2.4bA | 12.3 | 3.6aB | 7.1 | 2.8bA |
| 5.4 | 3.8 | 2.1bA | 22.0 | 4.7aA | 5.4 | 2.5bAB |
| 10.1 | 6.8 | 2.7aA | 3.9 | 2.2abC | 2.7 | 1.8bBC |
| 21.5 | 3.3 | 2.0aA | 3.4 | 2.1aC | 0.5 | 1.1aC |

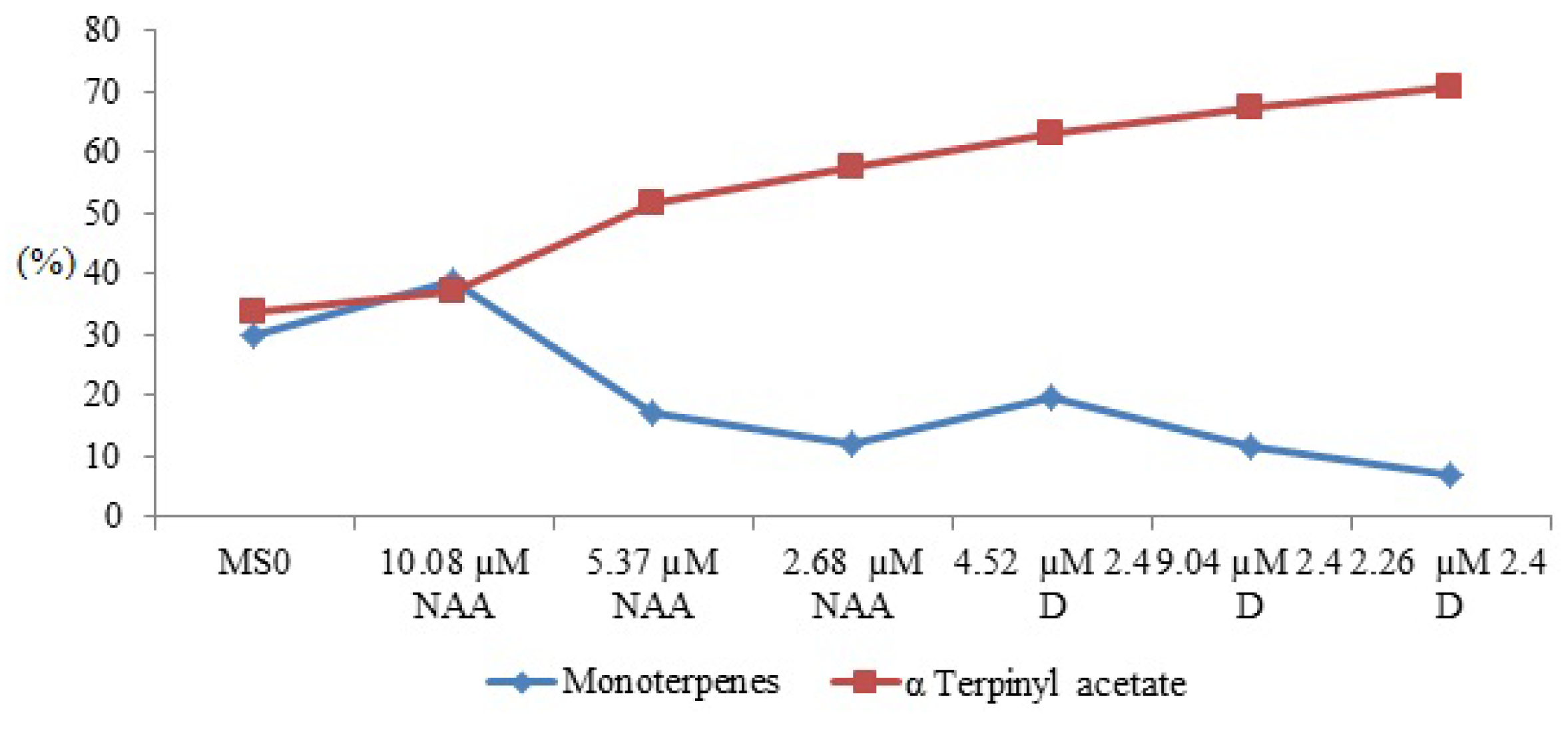
| Peak no. | Compound | RIexp | RIlit | MS0a | NAA (µM) | 2,4-D (µM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2.68a | 5.37a | 10.1a | 2.68b | 5.37b | 10.1b | 2.26a | 4.52a | 9.0a | 2.26b | 4.52b | |||||
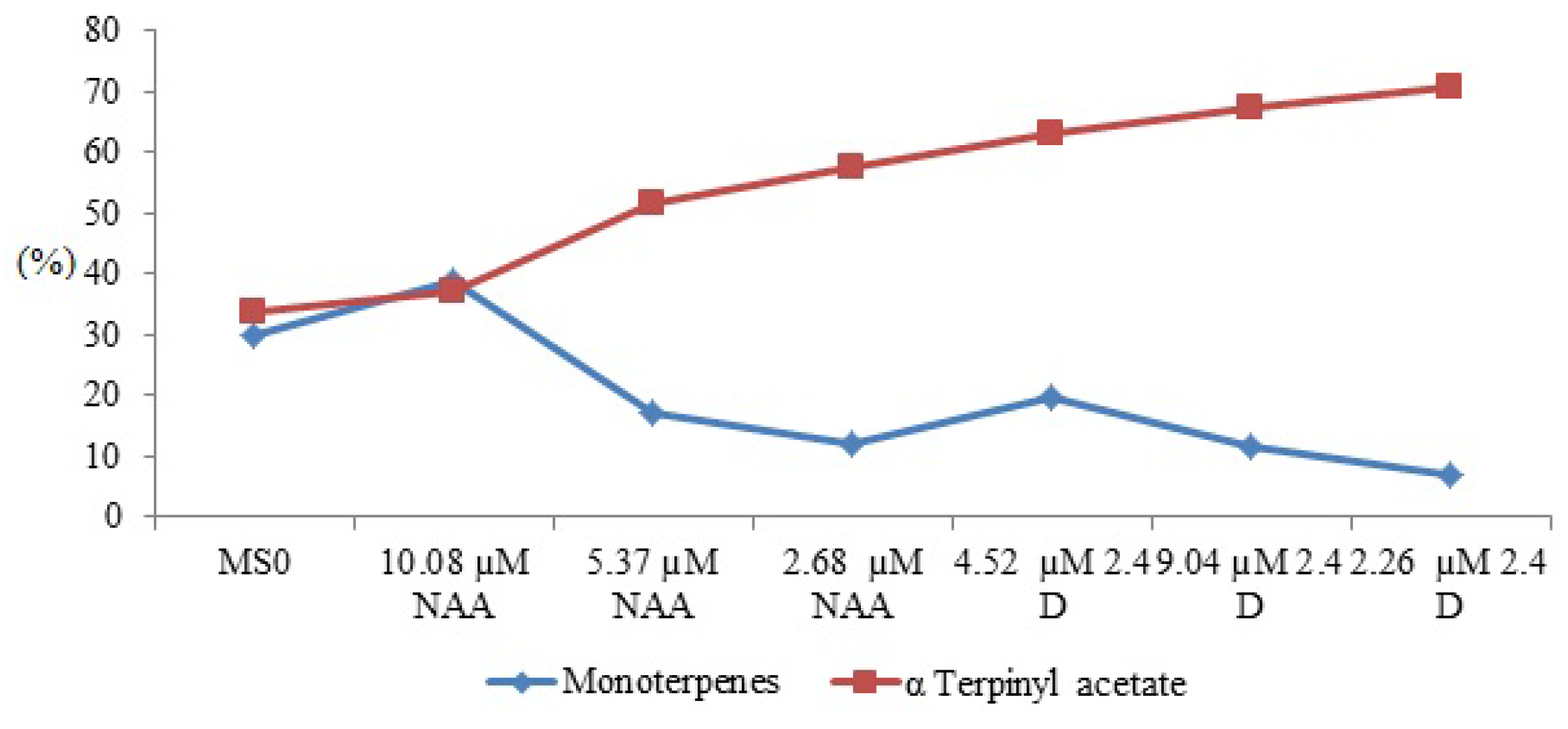
| 1. | α-thujene | 917 | 923 | - | - | - | 1.8 | - | 0.2 | - | - | - | - | - | - |
| 2. | α-pinene | 926 | 933 | 9.0 | 1.9 | 3.1 | 5.4 | 1.6 | 2.4 | 2.2 | 6.9 | - | - | 2.2 | - |
| 3. | camphene | 941 | 952 | 8.0 | 1.8 | 2.9 | 5.2 | 1.3 | 2.3 | 0.9 | - | 2.0 | 1.6 | 2.1 | - |
| 4. | sabinene | 972 | 973 | 2.8 | 1.9 | 1.7 | - | 0.8 | 1.3 | 2.1 | - | 2.1 | - | - | - |
| 5. | β-pinene | 977 | 980 | 2.1 | 2.5 | 3.9 | 6.4 | 2.1 | 2.9 | 2.9 | - | - | - | 2.7 | - |
| 6. | 1-octen-3-ol | 976 | 978 | - | - | - | 6.0 | - | - | - | - | 2.6 | - | - | - |
| 7. | α-limonene | 1,033 | 1,031 | 16.5 | 5.6 | 12.8 | 9.8 | 4.0 | 6.2 | 2.9 | - | 6.8 | 5.4 | 5.3 | - |
| 8. | terpinolene | 1,087 | 1,084 | - | - | - | - | - | 0.6 | 1.4 | - | - | - | - | - |
| 9. | borneol | 1,133 | 1,165 | 2.9 | 1.4 | 3.6 | 5.7 | 1.0 | 1.8 | - | - | 2.0 | - | 3.0 | - |
| 10. | (Z)-p-menth-1-ol | 1,135 | 1,136 | - | 0.4 | 1.1 | - | - | 0.5 | - | - | 0.5 | - | 2.0 | - |
| 11. | p-menth-8-ol | 1,141 | 1,144 | 0.7 | 0.7 | 1.2 | - | 0.6 | 0.6 | - | - | - | - | - | - |
| 12. | isobornyl acetate | 1,287 | 1,285 | 16.8 | 15.6 | 2.7 | 11.3 | 16.4 | 19.2 | 21.4 | - | 1.1 | - | - | - |
| 13. | α-terpinyl acetate | 1,356 | 1,350 | 31.3 | 46.4 | 51.6 | 35.9 | 60.4 | 56.3 | 63.0 | 72.1 | 73.5 | 73.6 | 57.7 | 53.4 |
| 14. | β-isocumene | 1,408 | 1,403 | 4.0 | 1.7 | 2.8 | 3.2 | 0.7 | 2.0 | 2.1 | - | 2.0 | 1.6 | 3.1 | 1.5 |
| 15. | decanol acetate | 1,412 | 1,409 | 1.8 | 1.6 | - | - | - | - | 2.5 | - | - | - | - | - |
| 16. | β-caryophyllene | 1,420 | 1,418 | 1.5 | 0.7 | 1.0 | - | 0.4 | 0.3 | - | 15.2 | - | - | - | - |
| 17. | α-humulene | 1,454 | 1,454 | 3.3 | 1.9 | 1.8 | 3.0 | 1.7 | - | - | - | 1.8 | - | - | - |
| 18. | kessane | 1,532 | 1,528 | - | - | - | - | - | 7.8 | 3.1 | - | - | - | - | - |
| 19. | sesquisabinene hydrate | 1,543 | 1,545 | 1.5 | 1.2 | 1.8 | 3.6 | 1.2 | 1.7 | 2.4 | - | 1.5 | - | 1.8 | - |
| Peak no. | Compound | RIexp | RIlit | Relative area (%) |
|---|---|---|---|---|
| 1. | 2-hexenal | 854 | 854 | 0.50 |
| 2. | α-thujene | 917 | 921 | 13.64 |
| 3. | α-pinene | 926 | 939 | 9.97 |
| 4. | sabinene | 972 | 976 | 5.46 |
| 5. | β-pinene | 977 | 980 | 3.09 |
| 6. | 1-octen-3-ol | 976 | 978 | 9.23 |
| 7. | β-myrcene | 992 | 991 | 0.40 |
| 8. | 3-octanol | 995 | 993 | 0.81 |
| 9. | (Z)-hexenol acetate | 1,005 | 1,004 | 0.16 |
| 10. | α-limonene | 1,033 | 1,031 | 0.43 |
| 11. | (Z)-β-ocimene | 1,038 | 1,040 | 0.21 |
| 12. | (E)-β-ocimene | 1,048 | 1,050 | 2.54 |
| 13. | Decanal | 1,142 | 1,204 | 0.10 |
| 14. | α-cubebene | 1,356 | 1,351 | 0.69 |
| 15. | α-copaene | 1,381 | 1,376 | 1.59 |
| 16. | β-bourbonen | 1,389 | 1,384 | 0.88 |
| 17. | β-cubebene | 1,392 | 1,390 | 1.35 |
| 18. | α-gurjunene | 1,397 | 1,409 | 1.75 |
| 19. | β-caryophyllene | 1,420 | 1,418 | 30.34 |
| 20. | β-gurjunene | 1,429 | 1,432 | 0.41 |
| 21. | (Z)-β-farnesene | 1,434 | 1,433 | 0.15 |
| 22. | (Z)-muurola-4(14),5-diene | 1,444 | 1,460 | 0.14 |
| 23. | (E)-β-farnesene | 1,454 | 1,458 | 0.18 |
| 24. | α-humulene | 1,454 | 1,454 | 1.82 |
| 25. | alloaromadendrene | 1,468 | 1,461 | 0.19 |
| 26. | γ-muurolene | 1,471 | 1,477 | 0.08 |
| 27. | germacrene D | 1,479 | 1,480 | 8.20 |
| 28. | β-selinene | 1,489 | 1,485 | 0.25 |
| 29. | Β-guaiene | 1,494 | 1,490 | 0.29 |
| 30. | (E)-β-guaiene | 1,498 | 1,500 | 3.77 |
| 31. | germacrene-D-ol | 1,586 | 1,574 | 0.58 |
| 32. | caryophyllenoxide | 1,592 | 1,581 | 0.72 |
| 33. | α-muurolol <epi> | 1,641 | 1,641 | 0.08 |
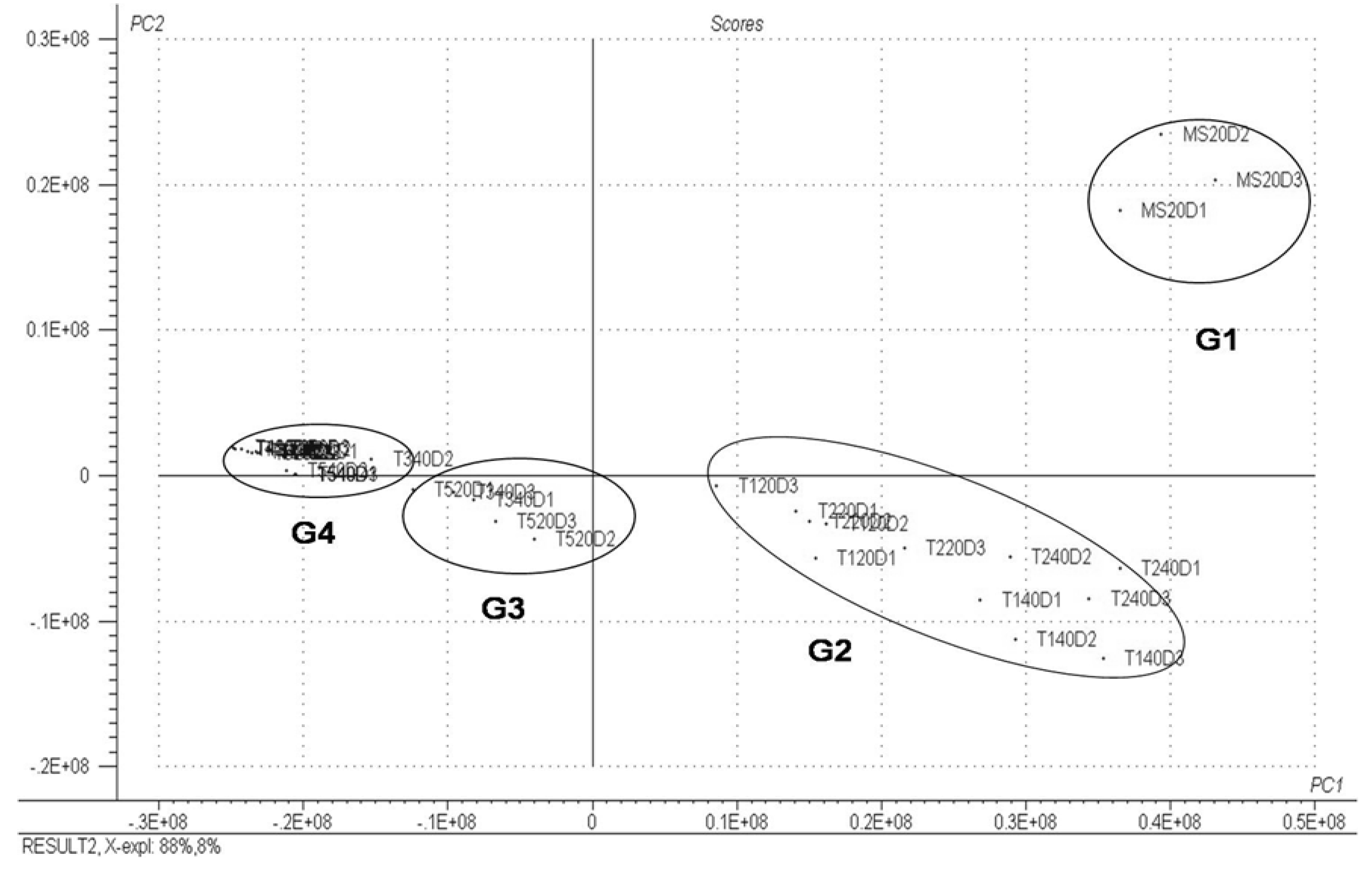
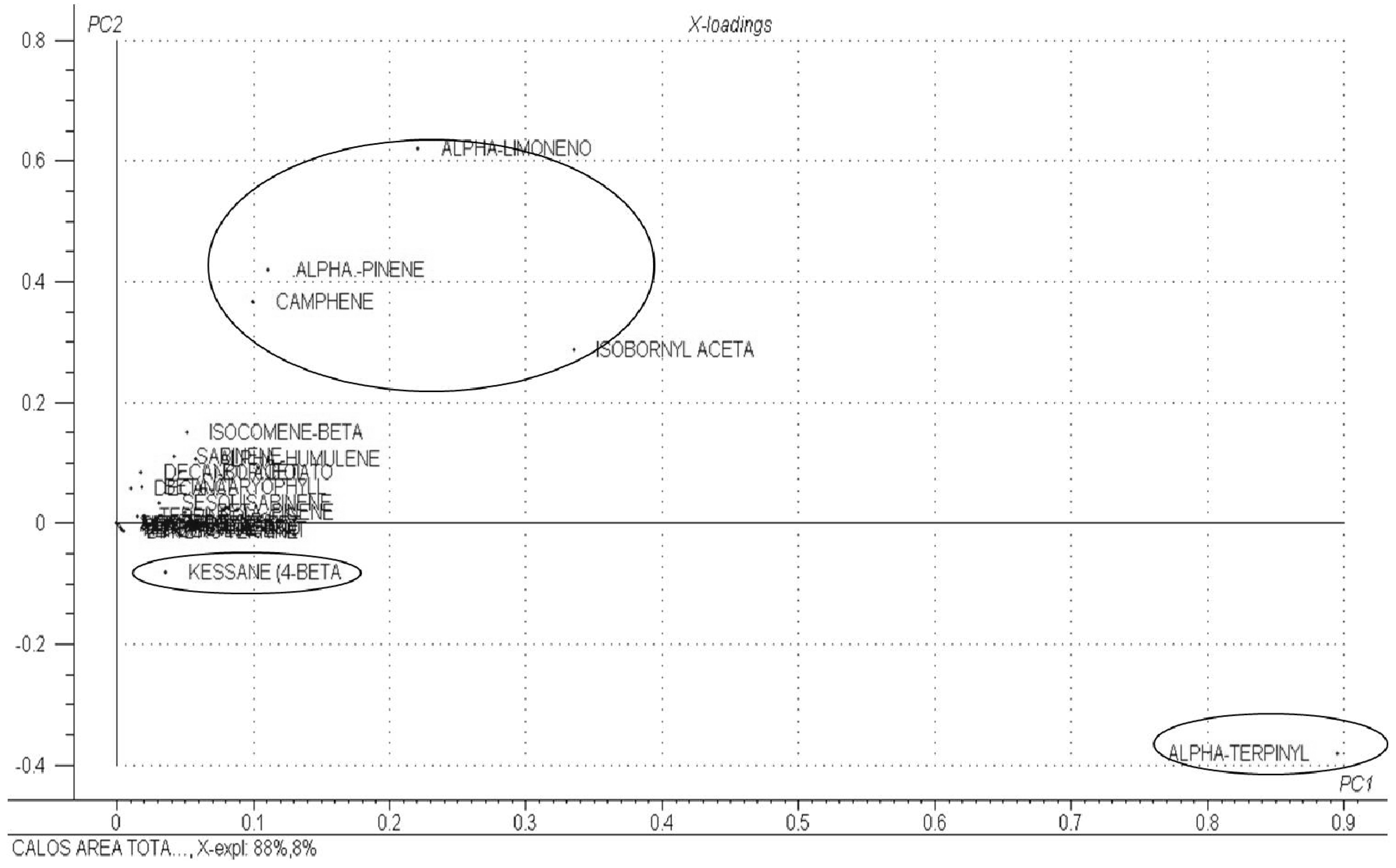

3. Experimental
3.1. Plant Material
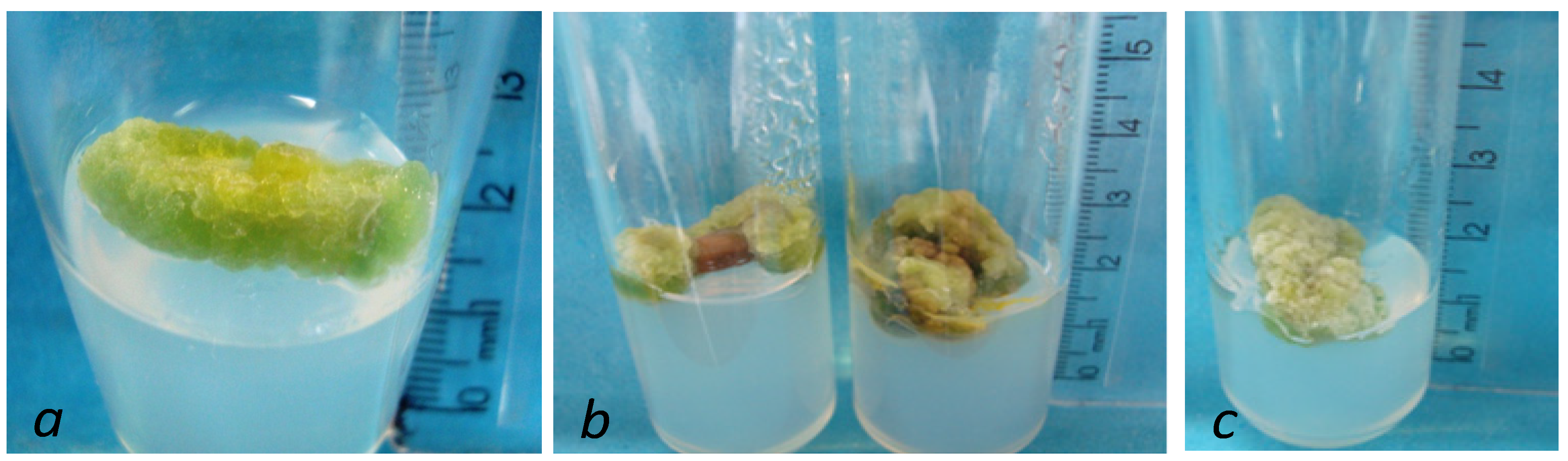
3.2. In Vitro Establishment of Plectranthus Ornatus
 and the counting numbers on
and the counting numbers on  . Data were analyzed using the SISVAR program, v 4.3, developed by Ferreira [22].
. Data were analyzed using the SISVAR program, v 4.3, developed by Ferreira [22].3.3. In Vitro Production and Establishment of Callus Tissues
3.4. Characterization of Metabolites Produced in Callus Tissues
Extraction of Volatile Organic Compounds (VOC)
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Lukhoba, C.W.; Simmonds, M.S.J.; Paton, A.J. Plectranthus: A review of ethnobotanical uses. J. Ethnopharmacol. 2006, 103, 1–24. [Google Scholar] [CrossRef]
- Dellar, J.E.; Cole, M.D.; Waterman, P.G. Antimicrobial abietane diterpenoids from Plectranthus elegans. Phytochemistry 1996, 41, 735–738. [Google Scholar] [CrossRef]
- Wellsow, J.; Grayer, R.J.; Veitch, N.C.; Kokubun, T.; Lelli, R.; Kite, G.C.; Simmonds, M.S.J. Insect-antifeedant and antibacterial activity of diterpenoids from species of Plectranthus. Phytochemistry 2006, 67, 1818–1825. [Google Scholar] [CrossRef]
- Abdel-Mogib, M.; Albar, H.A.; Batterjee, S.M. Chemistry of the genus Plectranthus. Molecules 2002, 7, 271–301. [Google Scholar] [CrossRef]
- Zuzarte, M.R.; Dinis, A.M.; Cavaleiro, C.; Salgueiro, L.R.; Canhoto, J.M. Trichomes, essential oils and in vitro propagation of Lavandula pedunculata (Lamiaceae). Ind. Crop. Prod. 2010, 32, 580–587. [Google Scholar] [CrossRef]
- Dörnenburg, H. Plant cell culture technology-harnessing a biological approach for competitive cyclotides production. Biotechnol. Lett. 2008, 30, 1311–1321. [Google Scholar] [CrossRef]
- Ma, X.; Gang, D.R. Metabolic profiling of in vitro micropropagated and conventionally greenhouse grown ginger (Zingiber officinale). Phytochemistry 2006, 67, 2239–2255. [Google Scholar] [CrossRef]
- Charchoglyan, A.; Abrahamyan, A.; Fujii, I.; Boubakir, Z.; Gulder, T.A.M.; Kutchan, T.M.; Vardapetyan, H.; Bringmann, G.; Ebizuka, Y.; Beerhues, L. Differential accumulation of hyperforin and secohyperforin in Hypericum perforatum tissue cultures. Phytochemistry 2007, 68, 2670–2677. [Google Scholar] [CrossRef]
- Reyes-Escogido, M.L.; Gonzalez-Mondragon, E.G.; Vazquez-Tzompantzi, E. Chemical and pharmacological aspects of capsaicin. Molecules 2011, 16, 1253–1270. [Google Scholar] [CrossRef]
- Zhao, P.; Iwamoto, Y.; Kouno, I.; Egami, Y.; Yamamoto, H. Stimulating the production of homoisoflavonoids in cell suspension cultures of Caesalpinia pulcherrima using cork tissue. Phytochemistry 2004, 65, 2455–2461. [Google Scholar] [CrossRef]
- Lazzeri, P.A.; Hildebrand, D.F.; Collins, G.B. A procedure for plant regeneration from immature cotyledon tissue of soybean. Plant Mol. Biol. Re. 1985, 3, 160–167. [Google Scholar] [CrossRef]
- Kaseleht, K.; Leitner, E.; Paalme, T. Determining aroma-active compounds in Kama flour using SPME-GC/MS and GC–olfactometry. Flavour Frag. J. 2011, 26, 122–128. [Google Scholar] [CrossRef]
- Marks, T.R.; Simpson, S.E. Factors affecting shoot development in apically dominant Acer cultivars in vitro. J. Hortic. Sci. 1994, 69, 543–551. [Google Scholar]
- Affonso, V.R.; Bizzo, H.R.; Lima, S.S.; Esquibel, M.A.; Sato, A. Solid Phase Microextraction (SPME) analysis of volatile compounds produced by in vitro shoots of Lantana camara L. under the influence of auxins and cytokinins. J. Braz. Chem.Soc. 2007, 18, 1504–1508. [Google Scholar] [CrossRef]
- Maes, K.; Vercammen, J.; Pham-Tuan, H.; Sandra, P.; Debergh, P.C. Critical aspects for the reliable headspace analysis of plants cultivated in vitro. Phytochem. Anal. 2001, 12, 153–158. [Google Scholar] [CrossRef]
- Affonso, V.R.; Bizzo, H.R.; Lage, C.L.S.; Sato, A. Influence of growth regulators in biomass production and volatile profile of in vitro plantlets of Thymus vulgaris L. J. Agric. Food. Chem. 2009, 57, 6392–6395. [Google Scholar] [CrossRef]
- Williams, M.; Morales, M.T.; Aparicio, R.; Harwood, J.L. Analysis of volatiles from callus cultures of olive Olea europaea. Phytochemistry 1998, 47, 1253–1259. [Google Scholar] [CrossRef]
- Turner, G.W.; Gershenzon, J.; Croteau, R.B. Development of peltate glandular trichomes of peppermint. Plant Physiol. 2000, 124, 655–680. [Google Scholar] [CrossRef]
- Lane, A.; Boecklemann, A.; Woronuk, G.N.; Sarker, L.; Mahmoud, S.S. A genomics resource for investigating regulation of essential oil production in Lavandula angustifolia. Planta 2010, 231, 835–845. [Google Scholar] [CrossRef]
- Silva, S.; Sato, A.; Lage, C.L.S.; Gil, R.A.S.S.; Azevêdo, D.A.; Esquibel, M.A. Essential oil composition of Melissa officinalis L. in vitro produced under the influence of growth regulators. J. Braz. Chem. Soc. 2005, 16, 1387–1390. [Google Scholar] [CrossRef]
- Avato, P.; Fortunato, I.M.; Ruta, C.; D’Elia, R. Glandular hairs and essential oils in micropropagated plants of Salvia officinalis L. Plant Sc. 2005, 169, 29–36. [Google Scholar] [CrossRef]
- Ferreira, D.F. Análises estatísticas por meio do Sisvar para Windows versão 4.0 (in portuguese). Technical tips online. 2008. Available online: http://www.dex.ufla.br/~danielff/sotwares.htm (accessed on 4 November 2012).
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Lord, H.L.; Grant, R.P.; Walles, M.; Incledon, B.; Fahie, B.; Pawliszyn, J.B. Development and evaluation of a solid-phase microextraction probe for in vivo pharmacokinetic studies. Anal. Chem. 2003, 75, 5103–5115. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; p. 804. [Google Scholar]
- Sample Availability: Not available.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Passinho-Soares, H.C.; Meira, P.R.; David, J.P.; Mesquita, P.R.R.; Vale, A.E.d.; De M. Rodrigues, F.; De P. Pereira, P.A.; De Santana, J.R.F.; De Oliveira, F.S.; De Andrade, J.B.; et al. Volatile Organic Compounds Obtained by in Vitro Callus Cultivation of Plectranthus ornatus Codd. (Lamiaceae). Molecules 2013, 18, 10320-10333. https://doi.org/10.3390/molecules180910320
Passinho-Soares HC, Meira PR, David JP, Mesquita PRR, Vale AEd, De M. Rodrigues F, De P. Pereira PA, De Santana JRF, De Oliveira FS, De Andrade JB, et al. Volatile Organic Compounds Obtained by in Vitro Callus Cultivation of Plectranthus ornatus Codd. (Lamiaceae). Molecules. 2013; 18(9):10320-10333. https://doi.org/10.3390/molecules180910320
Chicago/Turabian StylePassinho-Soares, Helna C., Paloma R. Meira, Juceni P. David, Paulo R. R. Mesquita, Ademir E. do Vale, Frederico De M. Rodrigues, Pedro A. De P. Pereira, José Raniere F. De Santana, Fabio S. De Oliveira, Jailson B. De Andrade, and et al. 2013. "Volatile Organic Compounds Obtained by in Vitro Callus Cultivation of Plectranthus ornatus Codd. (Lamiaceae)" Molecules 18, no. 9: 10320-10333. https://doi.org/10.3390/molecules180910320