1. Introduction
Burkitt’s lymphoma is considered to be one of the clinically most aggressive of B-cell lymphomas. It is characterized by a translocation of the proto-oncogene
c-Myc and immunoglobulin promoter regions, which leads to constitutive expression of the MYC protein [
1,
2]. Burkitt’s lymphoma most commonly affects children (representing 30%–50% of childhood non-Hodgkin’s lymphomas) and young adults. If left untreated, this disease is rapidly fatal, as it progresses extremely fast (doubling time, 24–48 h), and it has a high propensity to metastasize into the central nervous system [
3]. Therapeutic intervention is initiated immediately, as it is accepted that the high growth fraction and the short doubling time of Burkitt’s lymphoma make intensive short-cycle chemotherapy a necessity. Treatments usually combine etoposide, prednisone, vincristine, cyclophosphamide, doxorubicine and rituximab (e.g., the EPOCH-R regiment) [
4,
5,
6]. Due to the intense chemotherapy, there are often acute cytotoxic effects, including neutropenia, thrombocytopenia, myelosuppression, nausea and vomiting, neurotoxicity, hyperglycemia, stomach problems, cardiotoxicity, hair loss, constipation, and hypotension [
3,
5,
7].
Achieving selective cytotoxicity against malignantly transformed cells remains the main goal in the development of anticancer therapeutics. Therefore, a continuous search for novel, more target-specific agents is needed. Using this approach, the switch of equilibrium between tumor-cell progression and cell death in favor of disease regression is possible, while sparing normal tissue. In our previous study, we demonstrated that compounds with the
N-amidinopiperidine moiety are selectively cytotoxic towards malignantly transformed B-cells (Ramos and Daudi Burkitt’s lymphoma cells). The molecular pathways involve disruption of NFκB through inhibition of the trypsin-like activity of the proteasome [
8].
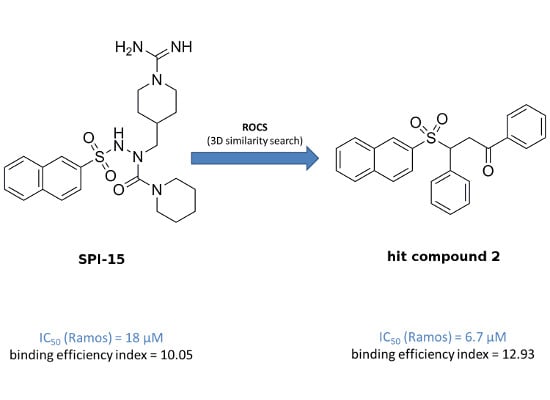
The overall objective of the present study was a search for improved inducers of apoptosis that show enhanced toxicity towards B-cell malignancies. To achieve this goal, a ligand-based three-dimensional (3D) similarity search was initiated, based on a previously described selective compound
SPI-15 (
Figure 1). We used the method of rapid overlay of chemical structures (ROCS), which performs shape-based overlays of conformers of a candidate molecule to a query molecule, in one or more conformations. The algorithm is based on the shape comparison and overlap of the volume of two molecules. In comparison to 2D similarity search methods, the new lead structures retrieved usually show relatively diverse chemical structures, and therefore enable scaffold hopping to be performed, and chemical diversity of a given set of compounds to be acquired [
9,
10].
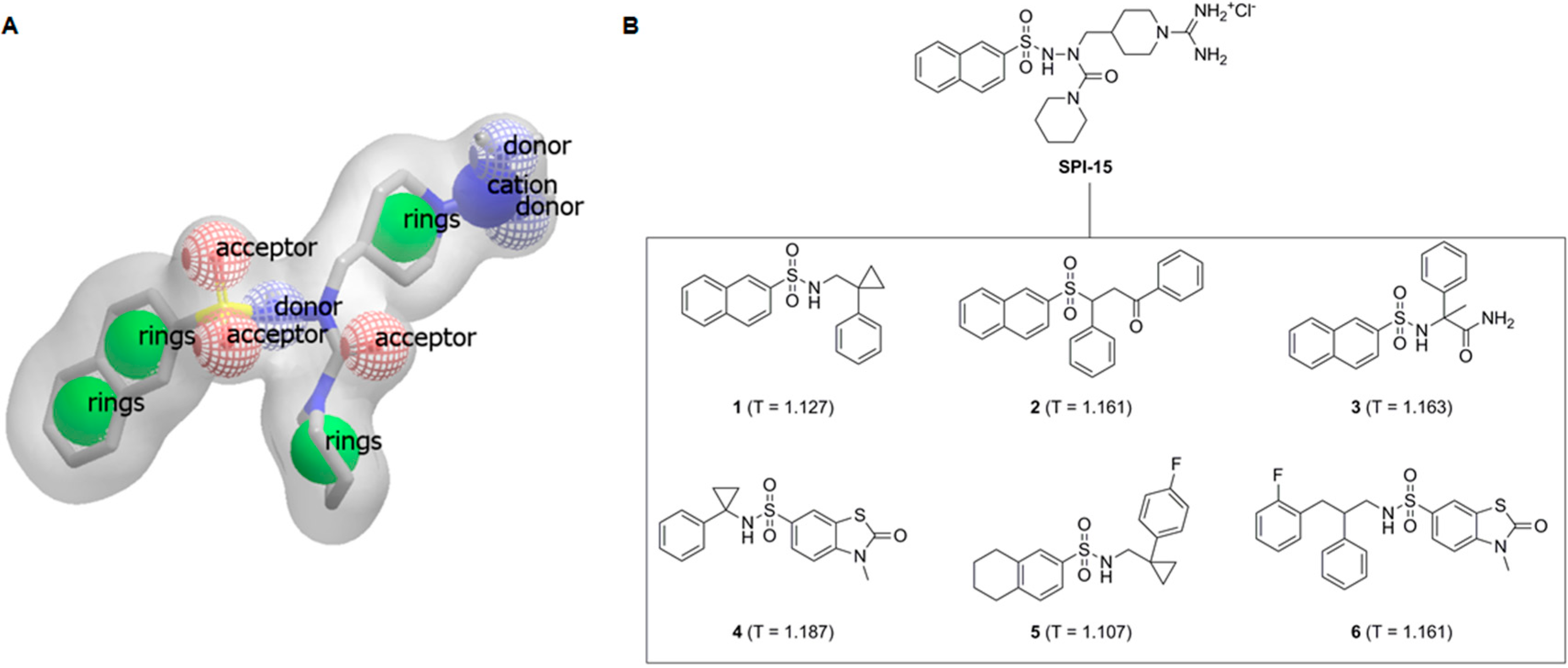
Figure 1.
(A) Shape and features of compound SPI-15, as perceived by the rapid overlay of chemical structures (ROCS) software; (B) Structural formulae of SPI-15 and its spatial analogs investigated here with their TanimotoCombo scores, Compounds 1–6, T = TanimotoCombo score.
Figure 1.
(A) Shape and features of compound SPI-15, as perceived by the rapid overlay of chemical structures (ROCS) software; (B) Structural formulae of SPI-15 and its spatial analogs investigated here with their TanimotoCombo scores, Compounds 1–6, T = TanimotoCombo score.
3. Experimental Section
3.1. Computer Hardware and Compound Database Preparation
The computational study was carried out on a workstation with four dual-core AMD Opteron 2.0-GHz processors, 16 GB RAM, four 320-GB hard drives in a RAID10 array, and Nvidia GeForce 7900 graphic cards, running 64-bit Fedora 7. For the virtual screening, the ZINC drug-like subset was used, which now includes 11.3 million drug-like compounds [
13]. To cover as much conformational space as possible, the database was first processed with the Omega 2.4.3 software (OpenEye Scientific Software Inc, Santa Fe, NM, USA) using the default settings, which yielded an average of 152 conformations per compound [
14,
15].
3.2. Ligand-Based Virtual Screening
For the ligand-based 3D similarity search, the compound
SPI-15 was used as a query molecule [
8]. The Omega 2.4.3 software was used to generate six conformers of
SPI-15 (by the default settings the RMS values between the conformers are 0.5 Å or more). To compare the molecules in the ZINC drug-like database to all of the query conformers, ROCS 3.1.1 (OpenEye Scientific Software Inc., Santa Fe, NM, USA) was used, with the default settings [
11]. The single best overlay hits were ranked according to the TanimotoCombo scoring function, which considers similarities in the molecular shapes and colors (
i.e., atom types). From the compounds with the highest similarities to
SPI-15, six that were available were evaluated
in vitro (
Figure 1B, Compounds
1–
6).
3.3. Materials and Methods
Compounds 1–6 were purchased from Asinex Chemicals (Winston-Salem, NC, USA). Prior to use in the biochemical assays, the purities of all of the compounds were determined by HPLC. Compound 2 was subsequently carefully characterized. The melting point of Compound 2 was determined on a Reichert hot-stage apparatus, and is uncorrected. The 1H-NMR spectrum was recorded on a Bruker Avance 400 DPX spectrometer (Bruker BioSpin Corporation, Billerica, MA, USA) at 295 K, and is reported as ppm, using solvent as an internal standard (DMSO-d6 at 2.50 ppm). The coupling constants (J) are given in Hz, and the splitting patterns are designated as: s, singlet; d, doublet; dd, double doublet; and m, multiplet. The 13C-NMR spectrum was recorded on a Bruker Avance 400 spectrometer at 295 K, and is reported in ppm using solvent as internal standard. Infrared (IR) spectrum was recorded on Fourier transform-IR spectrometer (Thermo Nicolet Nexus 470 ESP, Thermo Fisher Scietific Inc, Waltham, MA, USA). Mass spectrometry and high-resolution mass measurement was performed on a VG-Analytical Autospec Q mass spectrometer (VG Analytical, Manchester, UK). The HPLC analysis was run on an Agilent 1100 system equipped with a quaternary pump and a multiple wavelength detector. An Agilent Eclipse C18 column (4.6 mm × 50 mm; 5 mm) was used, with a flow rate of 1.0 mL/min, detection at 254 nm, and an eluent system of: A, 0.1% trifluoroacetic acid in H2O; B, MeOH. The following gradient was applied: 0–3 min, 40% B; 3–18 min, 40%–80% B; 18–23 min, 80% B; 23–30 min, 80%–40% B; run time, 30 min; temperature, 25 °C.
3-(Naphthalen-2-ylsulfonyl)-1,3-diphenylpropan-1-one (2) (Scheme 1)
Scheme 1.
Structural formula of hit compound 2.
Scheme 1.
Structural formula of hit compound 2.
White solid. Mp = 179.0–181.0 °C; 1H-NMR (400 MHz, DMSO-d6) δ (ppm) = 3.97 (dd, B part of ABX, JAB = 18.0 Hz, JBX = 4.5 Hz, COCH2CH, 1H), 4.10 (dd, A part of ABX, JAB = 18.0 Hz, JAX = 9.0 Hz, COCH2CH, 1H), 5.19 (dd, X part of ABX, JAX = 9.0 Hz, JBX = 4.5 Hz, COCH2CH, 1H), 7.18–7.27 (m, 3H, Ar-H), 7.29–7.33 (m, 2H, Ar-H), 7.46–7.51 (m, 2H, Ar-H), 7.60–7.68 (m, 3H, Ar-H), 7.70–7.75 (m, 1H, Ar-H), 7.92–7.96 (m, 2H, Ar-H), 8.02–8.10 (m, 3H, Ar-H), 8.35 (d, J = 1.5 Hz, 1H, Ar-H); 13C-NMR (100 MHz, DMSO-d6) δ (ppm) = 36.70, 65.41, 123.35, 127.58, 127.77, 128.06 (2C), 128.51, 128.66, 128.92, 129.37, 129.40, 129.97, 130.57, 131.43, 132.31, 133.57, 133.92, 134.69, 135.82, 195.13; IR (ATR) υ = 3406, 3052, 1681, 1593, 1448, 1338, 1303, 1233, 1141, 1124, 1072, 982, 747, 684 cm−1; ESI-MS [M+H]+: m/z = 401; HRMS (ESI) m/z calculated for C25H21O3S [M+H]+ 401.0773, found 401.0768; purity by HPLC: 99.10%.
3.4. Cell Culture
The Ramos, Thp-1, Jurkat, PC3 and MCF-7 cell lines were from ATCC (LGC Standards, Middlesex, UK), and Ramos-Blue cells were from Invivogen (San Diego, CA, USA). The Ramos, Thp-1 and Jurkat cell lines were cultured in RPMI 1640 medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (Gibco, Grand Island, NY, USA), 2 mM L-glutamine, 100 U/mL penicillin, 100 µg/mL streptomycin and 50 µM 2-mercaptoethanol (all from Sigma-Aldrich), in a humidified chamber at 37 °C and 5% CO2. PC3 cells were cultured in advanced Dulbecco’s Modified Eagle’s medium (DMEM)/F-12 (1:1; Gibco, Grand Island, NY, USA), HOS cells were cultured in DMEM (Sigma-Aldrich), and MCF-7 cells were cultured in advanced DMEM/F-12 with 10 μg/mL insulin, 20 ng/mL epidermal growth factor, and 0.5 μg/mL hydrocortisone (all from Sigma-Aldrich). Each medium was supplemented with 10% fetal bovine serum (Gibco), 2 mM l-glutamine, 100 U/mL penicillin, and 100 µg/mL streptomycin. The Ramos-Blue cells were cultured according to the manufacturer instructions.
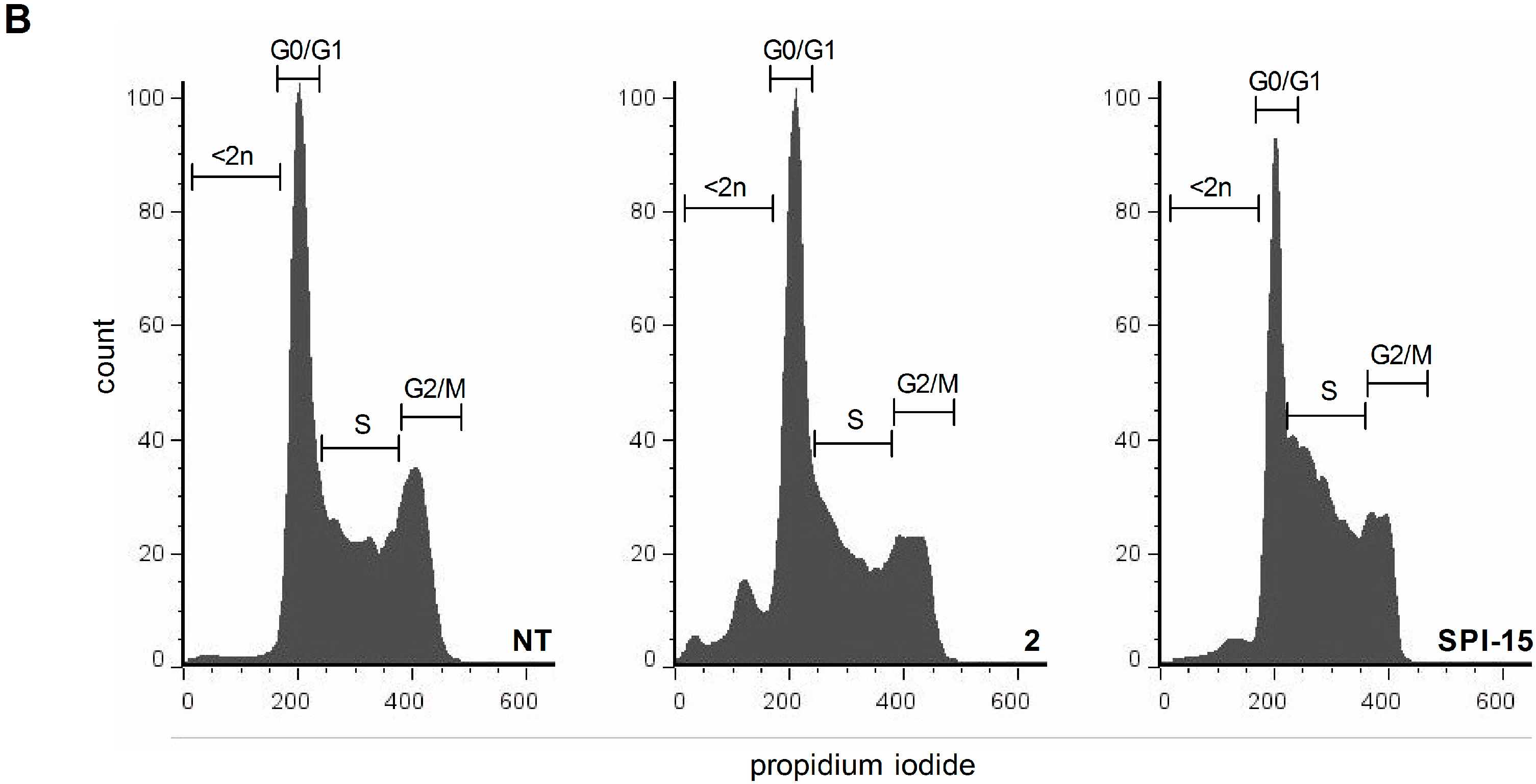
3.6. Cell-Cycle Analysis
The Ramos cells (5 × 105 cells) were treated with the vehicle and the compounds of interest for 24 h. The cells were then washed with phosphate-buffered saline (PBS) and fixed with 80% ethanol at –20 °C for 15 min. The fixed cells were pelleted by centrifugation and rehydrated in 5 mL PBS for 15 min at room temperature. The collected cells were resuspended in 500 µL staining buffer (3 µM propidium iodide, 100 mM Tris, pH 7.4, 150 mM NaCl, 1 mM CaCl2, 0.5 mM MgCl2, 0.1% Nonident P-40). After a 15 min incubation, the samples were analyzed using a FACScalibur and evaluated using the FlowJo software (BD Bioscience, Franklin Lakes, NJ, USA).
3.7. Annexin V Assay
We used annexin V-FITC Apoptosis Detection kits (Sigma-Aldrich), according to the manufacturer instructions. Briefly, following treatments with vehicle and the compound of interest, the Ramos cells were washed with PBS and resuspended in kit’s binding buffer. The cell suspensions were transferred to 5 mL tubes and the annexin V and propidium iodide were added. The cell suspensions were gently vortexed and incubated for 15 min at room temperature in the dark, followed by analysis using a FACScalibur flow cytometer (BD Bioscience, Franklin Lakes, NJ, USA).
3.8. Determination of Caspase 3/7 Activity
Caspase 3/7 activity was measured in total cell lysates using the fluorescent N-Acetyl-Asp-Glu-Val-Asp-7-amido-4-trifluoromethylcoumarin (Ac-DEVD-AFC) substrate (Bachem, Bubendorf, Switzerland). The Ramos cells incubated with vehicle or compounds for the indicated times were washed twice in PBS and resuspended in 200 µL ice-cold caspase lysis buffer (0.1% Triton X-100, 100 mM phosphate buffer, pH 6.0, 1.3 mM EDTA, 100 mM NaCl), sonicated, and left on ice for 30 min. After centrifugation (14,000× g, 15 min, 4 °C), the total protein concentrations in the supernatants were measured using BioRad Protein Assay kits (Bio-Rad, Hercules, CA, USA), according to the manufacturer instructions. Cell lysates (20 μg protein) were incubated for 30 min at 37 °C in caspase reaction buffer (20 mM PIPES, pH 7.2, 10% sucrose, 0.1% CHAPS, 1 mM EDTA, 100 mM NaCl), after which 100 µM Ac-DEVD-AFC peptide substrate was added. Immediately following substrate addition, the fluorescence intensities were monitored continuously for 30 min using a fluorescence microplate reader (BioTek Synergy HT, BioTek, Winooski, VT, USA) at the excitation and emission wavelengths of 405 nm and 535 nm, respectively. Data are expressed as the relative increase in fluorescence as a function of time (ΔF/Δt).
3.9. Quanti-Blue Assay
Ramos-Blue cells stably expressing an NFκB/AP-1-inducible secreted embryonic alkaline phosphate (SEAP) reporter construct were assayed for NFκB transcriptional activity changes upon pretreatment with vehicle and the compounds of interest for 1 h, and subsequent stimulation with 50 ng/mL recombinant TNF-α. The SEAP activity was determined in the supernatants, according to the manufacturer instructions. Briefly, the cell supernatants (40 µL) were added to 160 µL QUANTI-Blue reagent and incubated at 37 °C for 2 h. The absorbance was measured on a microplate reader (BioTek Synergy HT), at 640 nm.
3.10. Proteasome Activity Measurements
Purified human 20S proteasome (1 μg/mL; Boston Biochem, Inc., Cambridge, MA, USA) was incubated with vehicle or the compounds of interest at 37 °C for 30 min in assay buffer (20 mM Tris, pH 8.0, 0.5 mM EDTA, 1 mM dithiothreitol). Then, 100 μM Suc-Leu-Leu-Val-Tyr-7-amino-4-methylcoumarin (AMC), Boc-Leu-Leu-Arg-Arg-AMC, and Z-Leu-Leu-Glu-AMC (Bachem, Bubendorf, Switzerland) were added for the assessment of the chymotrypsin-like, trypsin-like and caspase-like activities, respectively. The flourescence of the released AMC was continuously monitored at the excitation and emission wavelengths of 320 nm and 420 nm, respectively, at 37 °C, using an automated microplate reader (BioTek Synergy HT). The measured relative fluorescence (RFU) was monitored over time.
3.11. Statistics
All experiments were performed at least three times, with average values expressed as means ± SD. Statistical significance was determined by the Student’s t-test. Differences were considered significant for p < 0.05.
4. Conclusions
This computationally supported search for novel structural analogs of SPI-15 resulted in the identification of six structurally related compounds. The initial screening showed a comparable toxicity profile to SPI-15 for one of these spatial analogs, Compound 2. Indeed, as well as having lower molecular mass and better ligand efficiency, Compound 2 showed improved activity, as a lower concentration was needed to induce caspase-dependent apoptosis of Ramos Burkitt’s lymphoma cells, in comparison with the parent compound, SPI-15. However, the molecular mechanisms of Compound 2 that lead to cell death are distinct to those of SPI-15, as Compound 2 does not involve inhibition of the proteasome or the NFκB pathway.
Further studies are needed to elucidate the molecular mechanisms and/or target(s) that are modulated by Compound 2. Despite significant differences in the molecular mechanisms, we believe that the lower concentrations needed to have the desirable effects represent a promising step towards improved and more selective agents against Burkitt’s lymphoma. In addition, the data presented here provide a solid basis for further ligand-based design of improved compounds, and more importantly, for the identification of the target(s) involved in Burkitt’s lymphoma. This target identification will undoubtedly provide valuable information for future structure-based optimization efforts in the battle against this disease, and against other hematological malignancies.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





