Appraisal of Antioxidant and Anti-Inflammatory Activities of Various Extracts from the Fruiting Bodies of Pleurotus florida

Abstract
:1. Introduction
2. Results and Discussion
2.1. Evaluation of Antioxidant Activity
2.1.1. Antioxidant Activity on β-Carotene-linoleic Acid

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solvent and Control | Sample Concentration (mg/mL) | |||
|---|---|---|---|---|
| 0.5 | 2.0 | 8.0 | 20.0 | |
| Acetone | 36.47 ± 0.11 | 77.05 ± 0.72 | 80.92 ± 0.54 | 91.62 ± 0.04 |
| Methanol | 31.56 ± 0.05 | 71.11 ± 0.14 | 87.99 ± 0.16 | 91.41 ± 0.08 |
| Hot water | 36.74 ± 0.14 | 69.04 ± 0.12 | 84.80 ± 0.14 | 90.42 ± 0.06 |
| BHT | 95.21 ± 0.17 | - | - | - |
| TOC | 96.02 ± 0.18 | - | - | - |
2.1.2. Reducing Power
| Solvent and Control | Sample Concentration (mg/mL) | |||
|---|---|---|---|---|
| 1.0 | 2.0 | 4.0 | 8.0 | |
| Acetone | 0.76 ± 0.05 | 1.32 ± 0.06 | 1.83 ± 0.03 | 1.95 ± 0.07 |
| Methanol | 0.87 ± 0.22 | 1.46 ± 0.21 | 1.82 ± 0.22 | 1.94 ± 0.19 |
| Hot water | 0.54 ± 0.18 | 0.73 ± 0.17 | 1.09 ± 0.16 | 1.88 ± 0.28 |
| BHT | 3.212 ± 0.49 | - | - | - |
| TOC | 2.162 ± 0.32 | - | - | - |
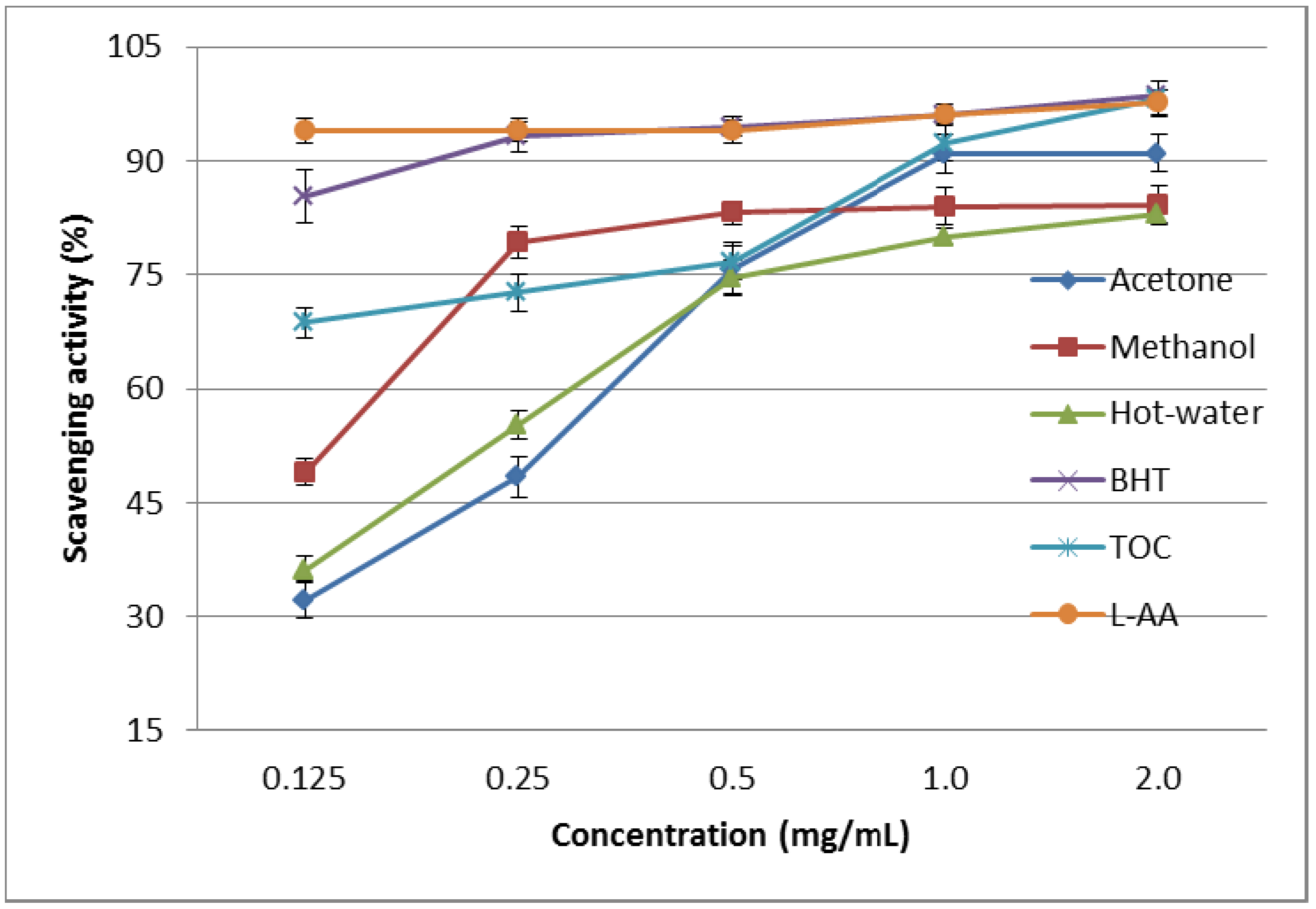
2.1.3. DPPH Scavenging Effect

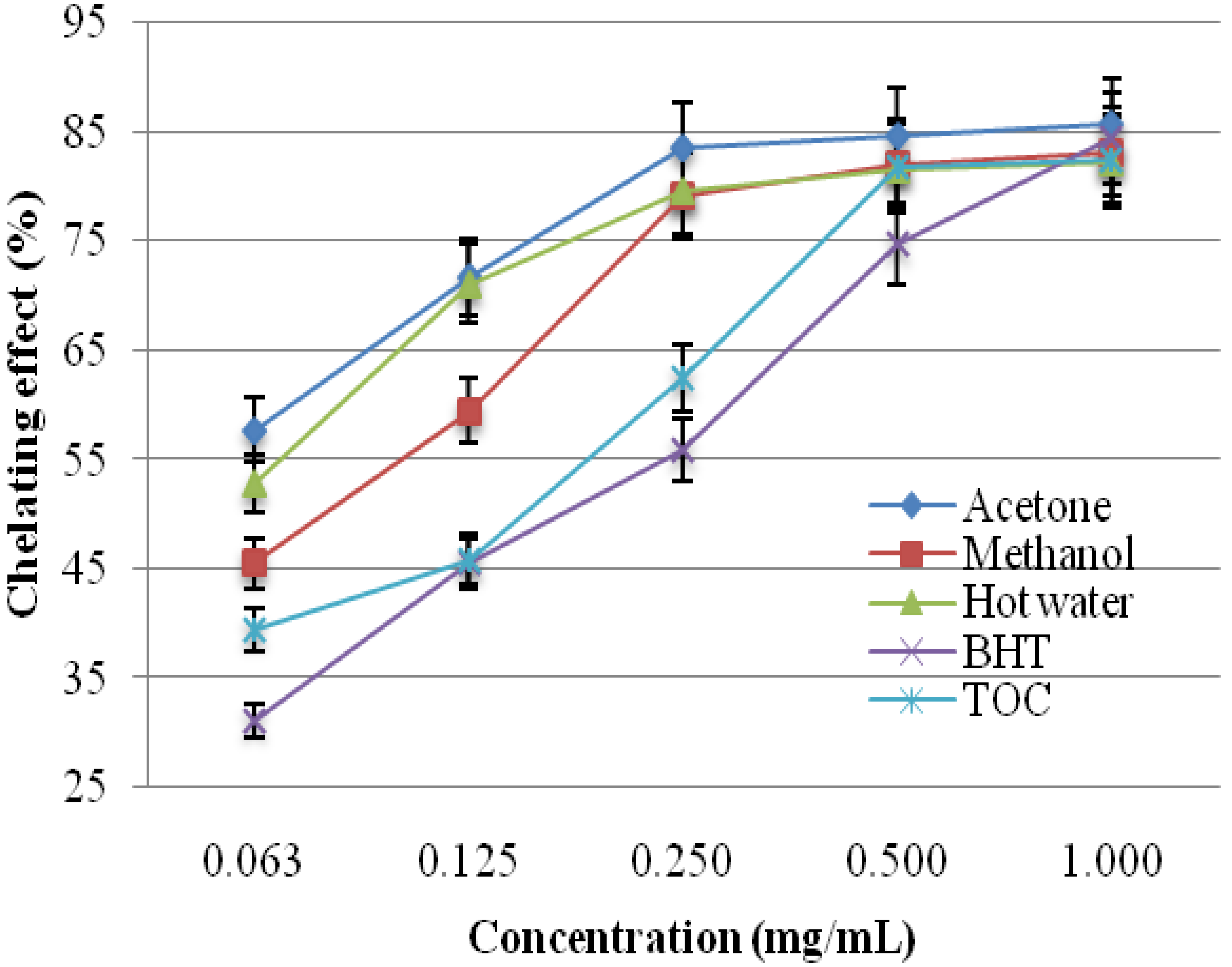
2.1.4. Chelating Effects on Ferrous Ions

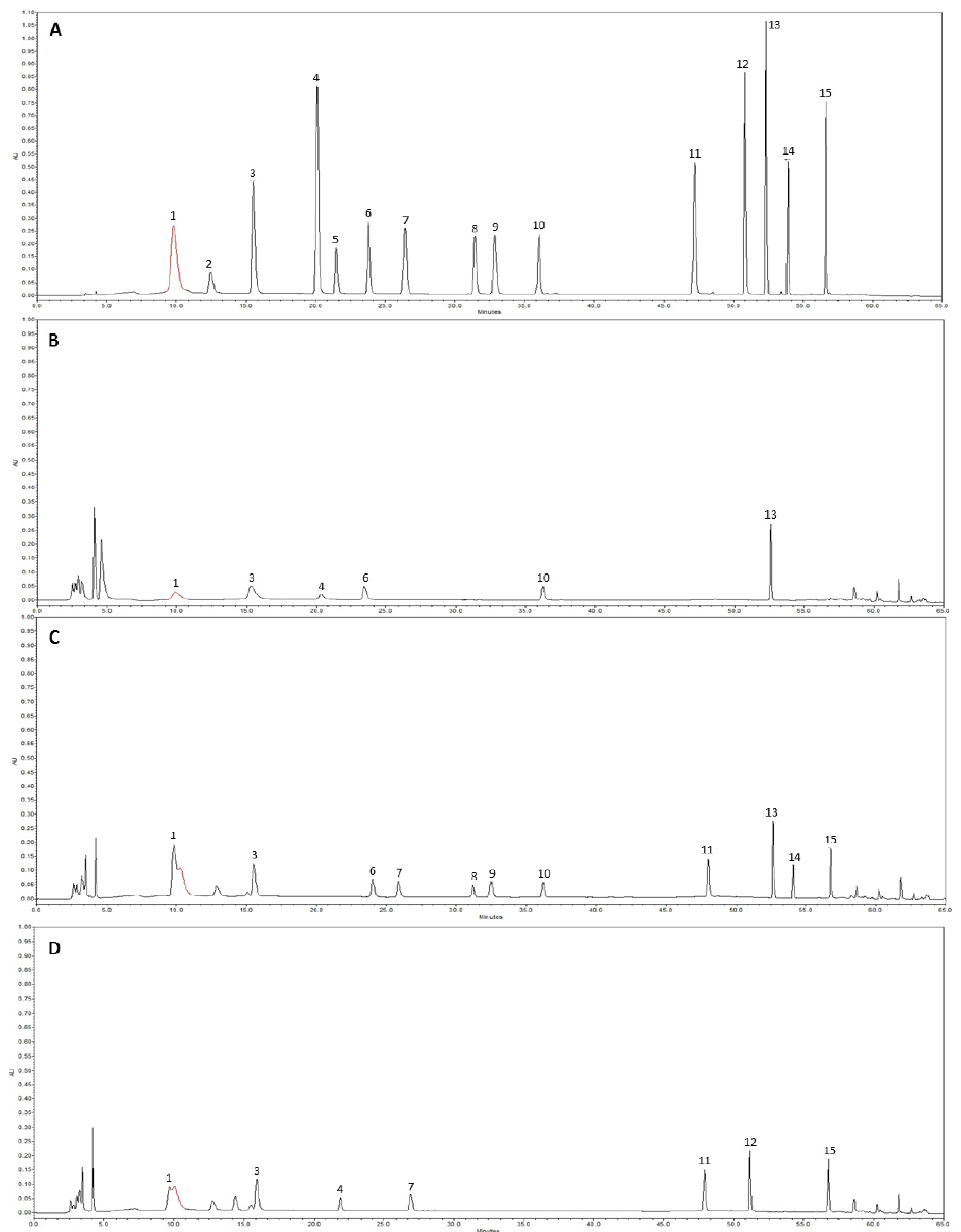
2.2. HPLC Analysis of Phenolic Compounds

2.3. Evaluation of Anti-Inflammatory Activity
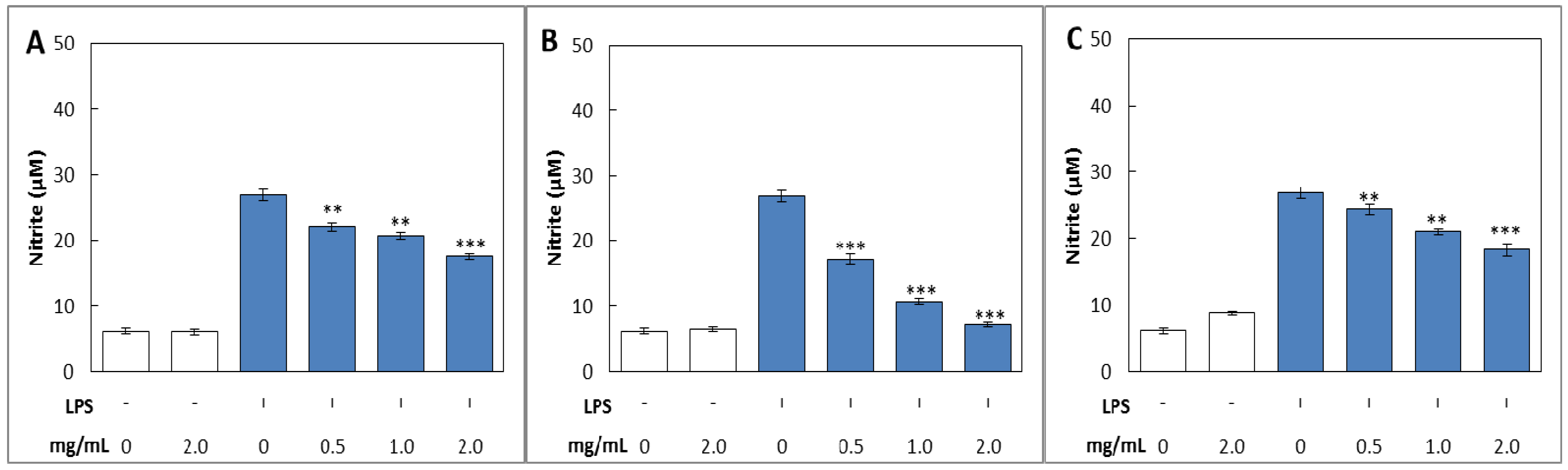
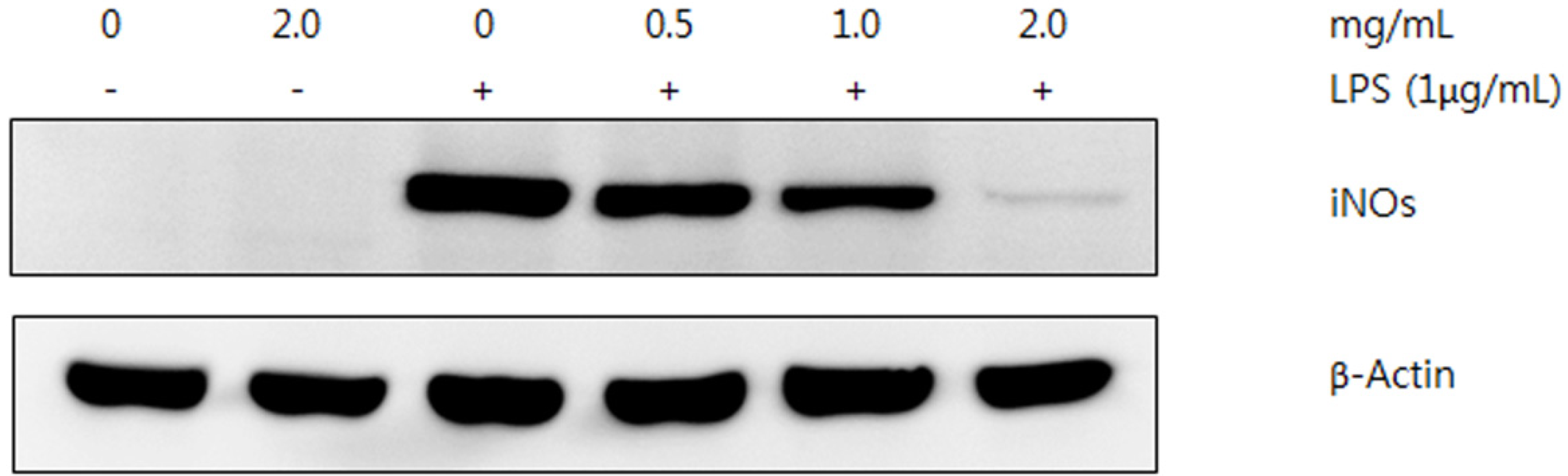
2.3.1. NO Production

2.3.2. Western Blot Analysis

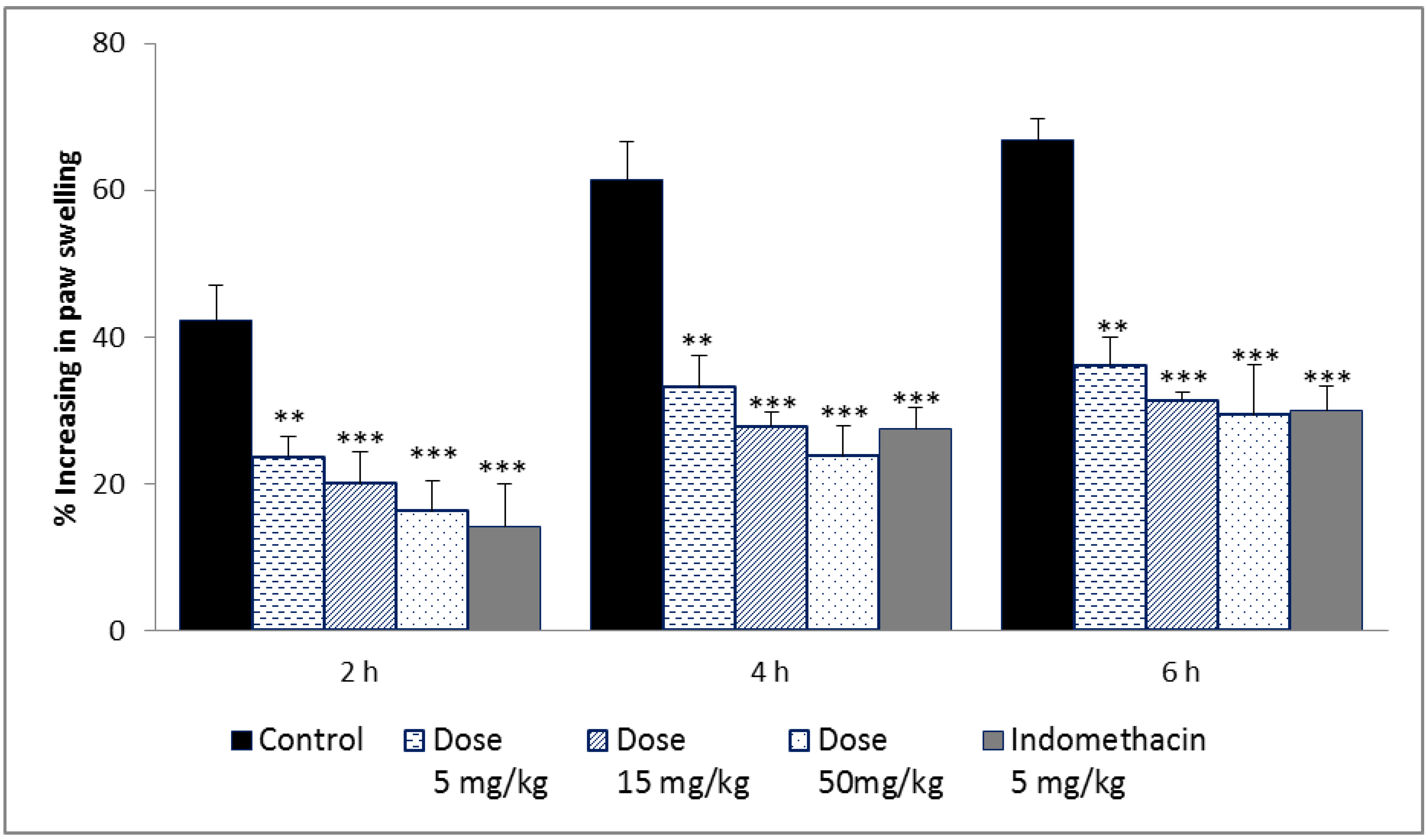
2.3.3. Carrageenan-Induced Rat Paw Edema

3. Experimental
3.1. Chemicals and Reagents
3.2. Animals
3.3. Mushrooms and Extraction
3.4. Evaluation of Anti-Oxidant Activity
3.4.1. Antioxidant Activity due to β-Carotene-linoleic Acid
3.4.2. Reducing Power
3.4.3. Scavenging Effect on DPPH Radicals
3.4.4. Chelating Effects on Ferrous Ions
3.5. HPLC Analysis of Phenolic Compounds
3.6. Evaluation of Anti-Inflammatory Activity
3.6.1. NO Assay
3.6.2. Western Blot Analysis
3.6.3. Carrageenan-Induced Hind Paw Edema in Rats
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflictts of Interest
References
- Choi, D.; Kang, S.H.; Song, Y.H.; Kwun, K.H.; Jeong, K.J.; Cha, W.S. Exopolysaccharide production in liquid culture of Pleurotus ferulae. J. Microbiol. Biotechnol. 2005, 146, 209–221. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar]
- Simic, M.G. Mechanisms of inhibition of free-radical processes in mutagenesis and carcinogenesis. Mutat. Res. 1988, 202, 377–386. [Google Scholar] [CrossRef]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Balkwill, F. Tumour necrosis factor and cancer. Nat. Rev. Cancer 2009, 9, 361–371. [Google Scholar] [CrossRef]
- Aggarwal, B.B. Nuclear factor-[kappa]B: The enemy within. Cancer Cell 2004, 6, 203–208. [Google Scholar]
- Knowles, R.G.; Moncada, S. Nitric oxide synthases in mammals. Biochem. J. 1994, 298, 249–258. [Google Scholar]
- Nathan, C.; Xie, Q.W. Nitric oxide synthases: Roles, tolls, and controls. Cell 1994, 78, 915–918. [Google Scholar] [CrossRef]
- Jayaprakasha, G.K.; Singh, R.P.; Sakariah, K.K. Antioxidant activity of grape seed (Vitis vinifera) extracts on peroxidation models in vitro. Food Chem. 2001, 73, 285–290. [Google Scholar] [CrossRef]
- Barros, L.; Ferreira, M.J.; Queiros, B.; Ferreira, I.C.; Bapista, P. Total phenols, ascorbic acid, β-carotene and lycopene in Portuguese wild edible mushrooms and their antioxidant activities. Food Chem. 2007, 103, 413–419. [Google Scholar] [CrossRef]
- Menaga, D.; Rajakumar, S.; Ayyasamy, P.M. Free radical scavenging activity of methanol extract of Pleurotus florida mushroom. Int. J. Pharm. Pharmaceut. Sci. 2013, 5, 601–606. [Google Scholar]
- Rahman, M.M.; Rahman, S.; Nahar, T.; Uddin, B.; Basunia, M.A.; Hossain, S. Antioxidant and anti-microbial activity of Pleurotus florida cultivated in Bangladesh. J. Medic. Plant Stud. 2013, 1, 166–175. [Google Scholar]
- Lee, Y.L.; Huang, G.W.; Liang, Z.C.; Mau, J.L. Antioxidant properties of three extracts from Pleurotus citrinopileatus. LWT-Food Sci. Technol. 2007, 40, 823–833. [Google Scholar] [CrossRef]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Frankel, E.N. Recent advances in lipid oxidation. J. Sci. Food Agric. 1991, 54, 495–511. [Google Scholar] [CrossRef]
- Herraiz, T.; Galisteo, J.; Chamorro, C. L-tryptophan reacts with naturally occurring and food-occurring phenolic aldehydes to give phenolic tetrahydro-β-caroline alkaloids: Activity as antioxidants and free radical scavengers. J. Agric. Food Chem. 2003, 51, 2168–2173. [Google Scholar] [CrossRef]
- Mau, J.L.; Chang, C.N.; Huang, S.J.; Chen, C.C. Antioxidant properties of methanolic extracts from Grifola frondosa, Morchella esculenta, and Termitomyces albuminosus mycelia. Food Chem. 2004, 87, 111–118. [Google Scholar] [CrossRef]
- Tsai, S.Y.; Huang, S.J.; Mau, J.L. Antioxidant properties of hot water extracts from Agrocybe cylindracea. Food Chem. 2006, 98, 670–677. [Google Scholar] [CrossRef]
- Lee, Y.L.; Yen, M.T.; Mau, J.L. Antioxidant properties of various extracts from Hypsizigus marmoreus. Food Chem. 2007, 104, 1–9. [Google Scholar] [CrossRef]
- Yamaguchi, R.; Tatsumi, Y.; Asano, M.; Kato, K.; Ueno, Y. Effect of metal salts and fructose on the autoxidation of methyl linoleate in emulsions. Agric. Biol. Chem. 1988, 52, 849–850. [Google Scholar] [CrossRef]
- Kim, M.Y.; Seguin, P.; Ahn, J.K.; Kim, J.J.; Chun, S.C.; Kim, E.H.; Seo, S.H.; Kang, E.Y.; Kim, S.L.; Park, Y.J.; et al. Phenolic compound concentration and antioxidant activities of edible and medicinal mushrooms from Korea. J. Agric. Food Chem. 2008, 56, 7265–7270. [Google Scholar] [CrossRef]
- Duan, X.; Wu, G.; Jiang, Y. Evaluation of antioxidant properties of lichi fruit phenolics in relation to pericarp browning prevention. Molecules 2007, 12, 759–771. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, K.; Huang, S.; Wang, H.; Mu, X.; He, C.; Ji, X.; Zhang, J.; Huang, F. Antioxidant activity of microwave-assisted extract of longan (Dimocarpus longan Lour.) peel. Food Chem. 2008, 106, 1264–1270. [Google Scholar] [CrossRef]
- Zhao, M.; Yang, B.; Wang, J.; Li, B.; Jiang, Y. Identification of the major flavonoids from pericarp tissues of lychee fruit in relation to their antioxidant activities. Food Chem. 2006, 98, 539–544. [Google Scholar] [CrossRef]
- Prasad, K.N.; Divakar, S.; Shivamurthy, G.R.; Aradhya, S.M. Isolation of a free radical scavenging antioxidant from water spinach (Ipomoea aquatica Forsk). J. Sci. Food Agric. 2005, 85, 1461–1468. [Google Scholar] [CrossRef]
- Rangkadilok, N.; Sitthimonchai, S.; Worasuttayangkurn, L.; Mahidol, C.; Ruchirawat, M.; Satayavivad, J. Evaluation of free radical scavenging and antityrosinase activities of standardized longan fruit extract. Food Chem. Toxicol. 2007, 45, 328–336. [Google Scholar] [CrossRef]
- Green, I.C.; Wagner, D.A.; Glowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.B. Analysis of nitrate, nitrite and [15N] nitrate in biological fluids. Analyt. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Won, S.Y.; Park, E.H. Anti-inflammatory and related pharmacological activities of cultured mycelia and fruiting bodies of Cordyceps militaris. J. Ethnoparmacol. 2005, 95, 555–561. [Google Scholar]
- Kim, H.G.; Yoon, D.H.; Lee, W.H.; Han, S.G.; Shrestha, B.; Kim, C.H.; Lim, M.H.; Chang, W.C.; Lim, S.Y.; Choi, S.A.; et al. Phellinus linteus inhibits inflammatory mediators by suppressing redox-based NF-kB and MARKs activation in lipopolysaccharide-induced RAW 264.7 macrophage. J. Ethnoparmacol. 2007, 114, 307–315. [Google Scholar] [CrossRef]
- Vinegar, R.; Schreiber, W.; Hugo, R. Biphasic development of carrageenan oedema in rats. J. Pharmacol. Exp. Ther. 1969, 166, 96–103. [Google Scholar]
- Niemegeers, C.J.E.; Verbruggen, F.J.; Janssen, P.A.J. Effect of various drugs on carrageenan-induced oedema in the rat hind paw. J. Pharmacol. 1964, 16, 810–816. [Google Scholar] [CrossRef]
- Dapkevicius, A.; Venskutonis, R.; van-Beek, T.A.; Linssen, J.P. Antioxidant activity of extracts obtained by different isolation procedures from some aromatic herbs grown in Lithuania. J. Sci. Food Agric. 1998, 77, 140–146. [Google Scholar] [CrossRef]
- Gülçin, I.; Büyükokuroglu, M.E.; Oktay, M.; Küfrevioglu, O.I. Antioxidant and analgesic activities of turpentine of Pinus nigra Arn. subsp. pallsiana (Lamb.) Holmboe. J. Ethnopharmacol. 2003, 86, 51–58. [Google Scholar] [CrossRef]
- Cuendet, M.; Hostettmann, K.; Potterat, O.; Dyatmiko, W. Iridoid glucosides with free radical scavenging properties from Fagraea blumei. Helv. Chim. Acta 1997, 80, 1144–1152. [Google Scholar] [CrossRef]
- Dinis, T.C.; Madeira, V.M.; Almeida, L.M. Action of phenolic derivatives (acetaminophen, salicylate, and 5-amino salicylate) as inhibitors of membrane lipid peroxidation and as peroxyl radical scavengers. Arch. Biochem. Biophys. 1994, 315, 161–169. [Google Scholar] [CrossRef]
- Winter, C.A.; Risley, E.A.; Nuss, G.W. Carregeenin-induced edema in hind paw of the rat as assay for anti-inflammatory drugs. Proc. Soc. Exp. Biol. Med. 1962, 11, 544–547. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the fruiting body powder of Pleurotus florida are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Im, K.H.; Nguyen, T.K.; Shin, D.B.; Lee, K.R.; Lee, T.S. Appraisal of Antioxidant and Anti-Inflammatory Activities of Various Extracts from the Fruiting Bodies of Pleurotus florida. Molecules 2014, 19, 3310-3326. https://doi.org/10.3390/molecules19033310
Im KH, Nguyen TK, Shin DB, Lee KR, Lee TS. Appraisal of Antioxidant and Anti-Inflammatory Activities of Various Extracts from the Fruiting Bodies of Pleurotus florida. Molecules. 2014; 19(3):3310-3326. https://doi.org/10.3390/molecules19033310
Chicago/Turabian StyleIm, Kyung Hoan, Trung Kien Nguyen, Do Bin Shin, Kyung Rim Lee, and Tae Soo Lee. 2014. "Appraisal of Antioxidant and Anti-Inflammatory Activities of Various Extracts from the Fruiting Bodies of Pleurotus florida" Molecules 19, no. 3: 3310-3326. https://doi.org/10.3390/molecules19033310
APA StyleIm, K. H., Nguyen, T. K., Shin, D. B., Lee, K. R., & Lee, T. S. (2014). Appraisal of Antioxidant and Anti-Inflammatory Activities of Various Extracts from the Fruiting Bodies of Pleurotus florida. Molecules, 19(3), 3310-3326. https://doi.org/10.3390/molecules19033310