Studies on the Bioactive Flavonoids Isolated from Pithecellobium clypearia Benth

Abstract
:1. Introduction
2. Results and Discussion
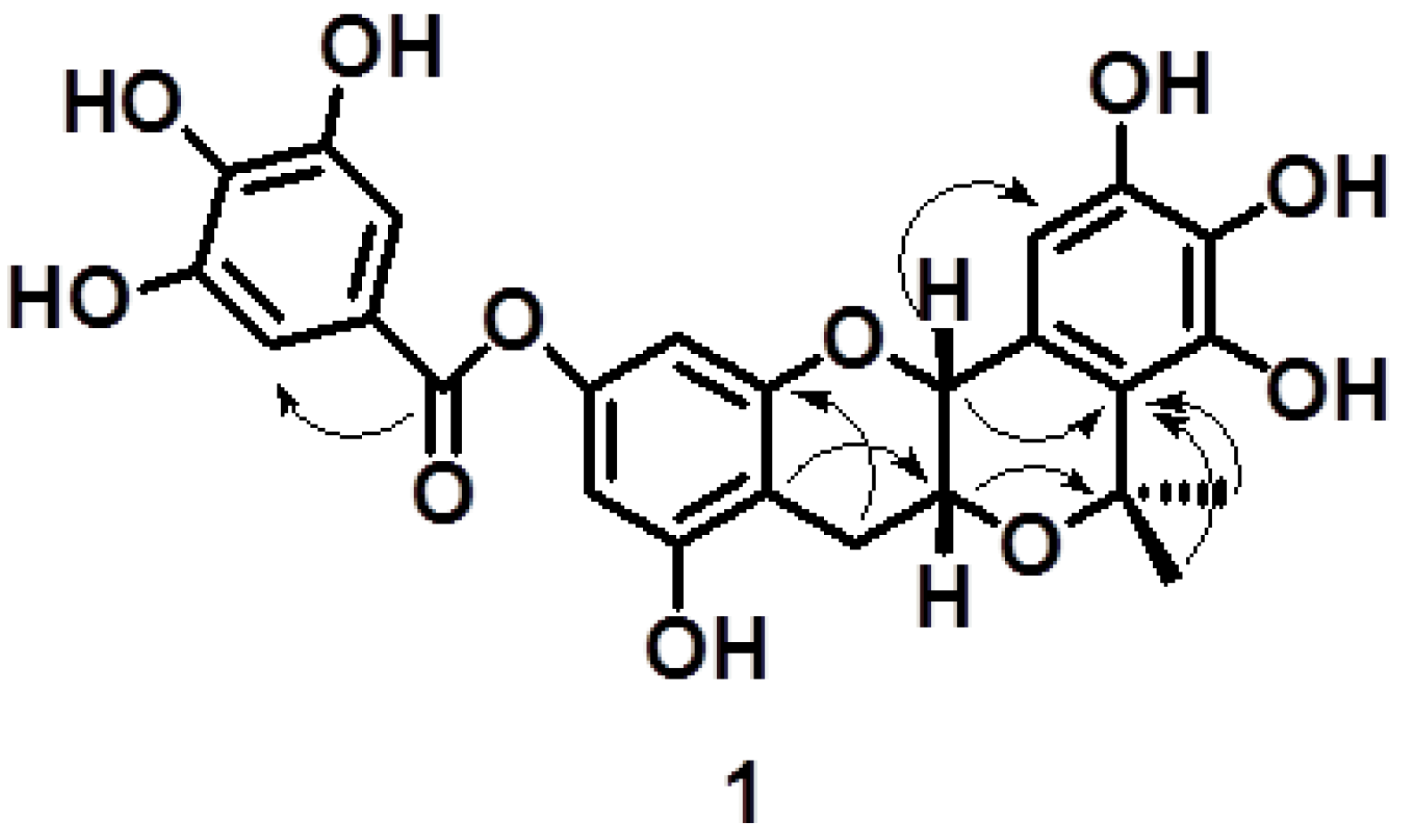
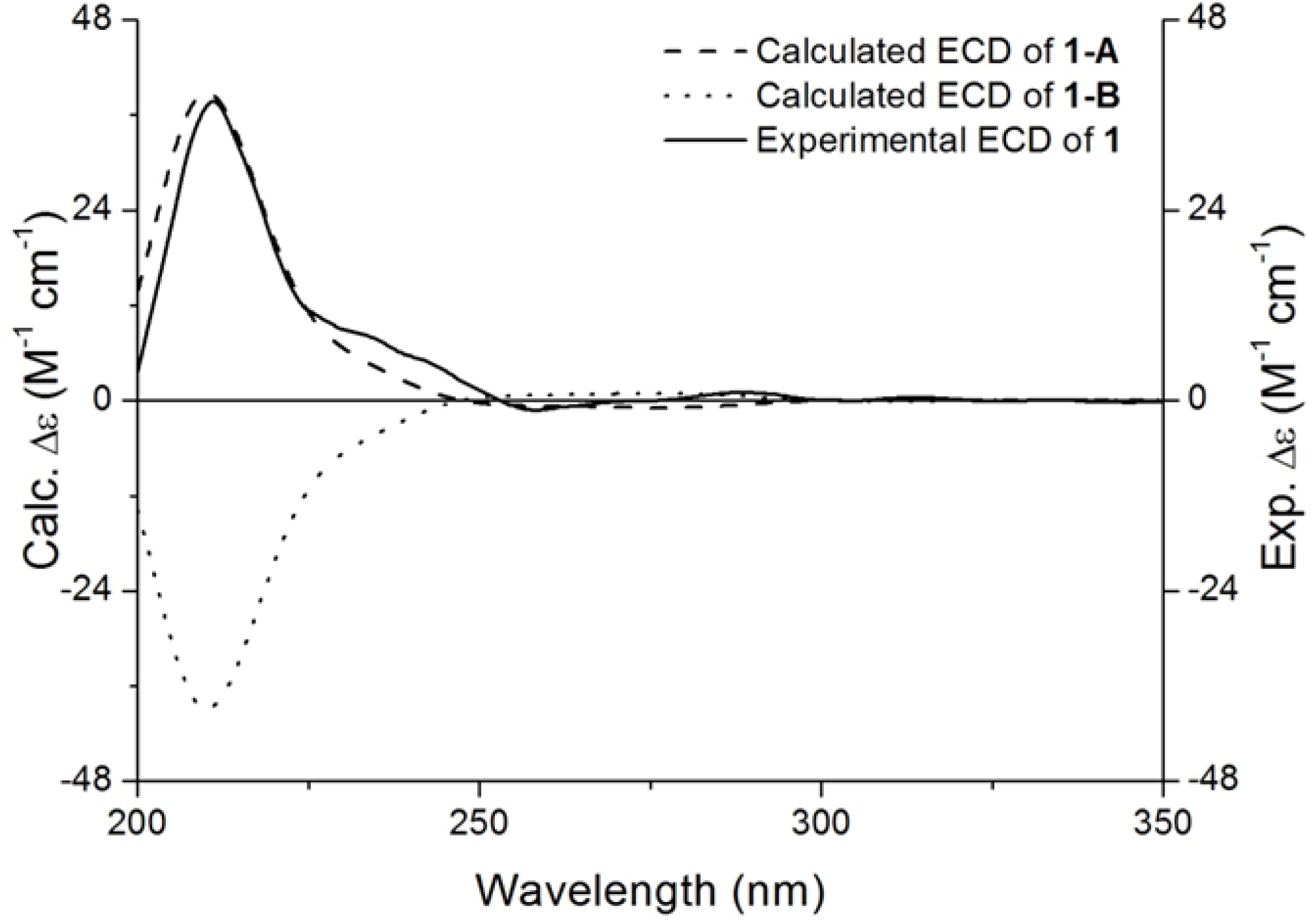
2.1. Characterisation of Compound 1



2.2. NA Inhibitory Effects

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Names | IC50 (µg/mL) | ||
|---|---|---|---|---|
| A/PR/8/34(H1N1) | A/Sydney/5/97(H3N2) | B/Jiangsu/10/2003 | ||
| 1 | 7-O-galloylplumbocatechin A | 29.77 ± 6.12 | 32.23 ± 1.65 | 39.15 ± 4.67 |
| 2 | (−)-5,3′,4′,5′-tetrahydroxyflavan-7-gallate | 36.91 ± 3.80 | >40 | >40 |
| 3 | (+)-3,5,3′,4′,5′-pentahydroxyflavan-7-gallate | >40 | >40 | >40 |
| 4 | (−)-7,4′-di-O-galloyltricetiflavan | >40 | >40 | >40 |
| positive control | zanamivir | 3.00 × 10−5 ± 2.03 × 10−6 | 2.10 × 10−4 ± 1.84 × 10−5 | 3.00 × 10−4 ± 2.09 × 10−5 |
2.3. Inhibition of Cytokine Production in H1N1-Infected Cells


3. Experimental
3.1. Plant Material
3.2. Chemicals and Instruments
3.3. Cells and Virus Strains
3.4. Extraction and Isolation
3.5. Characterization of Compounds 1–4
| No. position | 1 | 13C | 2 | 13C | 3 | 13C | 4 | 13C |
|---|---|---|---|---|---|---|---|---|
| δH (J in Hz) | δH (J in Hz) | δH (J in Hz) | δH (J in Hz) | |||||
| 2 | 4.49 (s) | 71.1 | 4.78 (d, 9.9) | 77.5 | 4.57 (d, 6.4) | 56.8 | 4.89 (d, 10.4) | 76.7 |
| 3 | 4.17 (d, 4.4) | 62.4 | 1.85 (m) 2.02 (m) | 29.1 | 3.89 (m) | 81.2 | 1.86 (m) 2.11 (m) | 28.7 |
| 4 | 2.83 (dd17.2, 4.4) 2.64 (d, 17.2) | 25.8 | 2.60 (br s) | 19.6 | 2.66 (dd, 16.4, 4.8) 2.44 (overlapped) | 27.1 | 2.62 (2H, br s) | 19.0 |
| 5 | 155.5 | 156.4 | 155.1 | 155.8 | ||||
| 6 | 6.14 (s) | 100.9 | 6.16 (s) | 100.9 | 6.17 (s) | 100.4 | 6.18 (s) | 100.6 |
| 7 | 149.9 | 150.1 | 149.8 | 149.7 | ||||
| 8 | 6.00 (s) | 101.0 | 6.09 (s) | 101.3 | 6.11 (s) | 100.6 | 6.13 (s) | 100.9 |
| 9 | 156.7 | 156.5 | 156.1 | 156.0 | ||||
| 10 | 104.9 | 107.4 | 105.6 | 107.0 | ||||
| 1' | 123.2 | 133.0 | 132.6 | 138.7 | ||||
| 2' | 121.1 | 6.31 (s) | 105.5 | 6.25 (s) | 105.7 | 6.43 (s) | 104.9 | |
| 3' | 144.5 | 146.4 | 145.7 | 150.2 | ||||
| 4' | 134.3 | 132.0 | 129.4 | 126.4 | ||||
| 5' | 142.8 | 146.4 | 145.7 | 150.2 | ||||
| 6' | 6.33 (s) | 108.8 | 6.31 (s) | 105.5 | 6.25 (s) | 105.7 | 6.43 (s) | 104.9 |
| 7' | 75.3 | |||||||
| 8' | 1.45 (3H, s) | 28.9 | ||||||
| 9' | 1.51 (3H, s) | 24.6 | ||||||
| 1" | 164.9 | 164.9 | 164.9 | 164.5 | ||||
| 2" | 118.9 | 118.9 | 118.4 | 118.9 | ||||
| 3" | 7.02 (s) | 109.5 | 7.04 (s) | 109.5 | 7.04 (s) | 109.0 | 7.04 (s) | 109.2 |
| 4" | 146.2 | 146.2 | 145.7 | 145.6 | ||||
| 5" | 139.6 | 139.6 | 139.2 | 139.2 | ||||
| 6" | 146.2 | 146.2 | 145.7 | 145.6 | ||||
| 7" | 7.02 (s) | 109.5 | 7.04 (s) | 109.5 | 7.04 (s) | 109.0 | 7.04 (s) | 109.2 |
| 1"' | 163.9 | |||||||
| 2"' | 118.4 | |||||||
| 3"' | 7.06 (s) | 109.0 | ||||||
| 4"' | 145.8 | |||||||
| 5"' | 138.9 | |||||||
| 6"' | 145.8 | |||||||
| 7"' | 7.06 (s) | 109.0 |
3.6. Neuraminidase Inhibition Assay
3.7. Cytokine Secretion Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jonathan, D.; Joshua, B.P.; Cecile, V.; David, J.D.E.; Lone, S. Mortality due to influenza in the United States — An annualized regression approach using multiple-cause mortality data. Am. J. Epidemiol. 2005, 163, 181–187. [Google Scholar] [CrossRef]
- Liu, Q.; Xiong, H.-R.; Lu, L.; Liu, Y.-Y.; Luo, F.; Hou, W.; Yang, Z.-Q. Antiviral and anti-inflammatory activity of arbidol hydrochloride in influenza A (H1N1) virus infection. Acta Pharm. Sin. 2013, 34, 1075–1083. [Google Scholar] [CrossRef]
- Chan, M.C.W.; Cheung, C.Y.; Chui, W.H.; Tsao, S.W.; Nicholls, J.M.; Chan, Y.O.; Chan, R.W.Y.; Long, H.T.; Poon, L.L.M.; Guan, Y.; et al. Proinflammatory cytokine responses induced by influenza A (H5N1) viruses in primary human alveolar and bronchial epithelial cells. Respir. Res. 2005, 6. [Google Scholar] [CrossRef] [Green Version]
- Shtyrya, Y.A.; Mochalova, L.V.; Bovin, N.V. Influenza virus neuraminidase: Structure and function. Acta Naturae 2009, 1, 26–32. [Google Scholar]
- Larisa, V.G. Molecular mechanisms of influenza virus resistance to neuraminidase inhibitors. Virus Res. 2004, 103, 199–203. [Google Scholar] [CrossRef]
- Yuen, K.Y.; Chan, P.K.; Peiris, M.; Tsang, D.N.C.; Que, T.L.; Shortridge, K.F.; Cheung, P.T.; To, W.K.; Ho, E.T.F.; Sung, R.; et al. Clinical features and rapid viral diagnosis of human disease associated with avian influenza A H5N1 virus. Lancet 1998, 351, 467–471. [Google Scholar] [CrossRef]
- Cheung, C.Y.; Poon, M.; Lau, A.S.; Luk, W.; Lau, Y.L.; Shortridge, K.F.; Gordon, S.; Guan, Y.; Peiris, J.S. Induction of proinflammatory cytokines in human macrophages by influenza A (H5N1) viruses: A mechanism for the unusual severity of human disease? Lancet 2002, 360, 1831–1837. [Google Scholar] [CrossRef]
- Alok, K.C.; Veena, C.V.; Sanjay, M.; Rashmi, S.; Shailesh, D.P.; Akhilesh, C.M. Host gene expression profiling in influenza A virus-infected lung epithelial (A549) cells: A comparative analysis between highly pathogenic and modified H5N1 viruses. Virol. J. 2010, 7. [Google Scholar] [CrossRef]
- Yaolan, L.; Kamtong, L.; Fenghe, Y.; Linda, S.M.O.; Vincent, E.C.O. Antiviral flavans from the leaves of Pithecellobium clypearia. J. Nat. Prod. 2006, 69, 833–835. [Google Scholar] [CrossRef]
- Xie, C.Y.; Lin, L.W. Study on the chemical constituents of Pithecellobium clypearia. J. Chin. Med. Mater. 2011, 34, 1060–1062. [Google Scholar]
- Wang, Y.G.; Dan, M.; Li, Y.H. Studies on the chemical constituents of Pithecellobium clypearia. J. Chin. Med. Mater. 2005, 28, 774. [Google Scholar]
- Li, Y.L.; Li, K.M.; Su, M.X.; Leung, K.T.; Chen, Y.W.; Zhang, Y.W. Studies on antiviral constituents in stems and leaves of Pithecellibium clypeari. China J. Chin. Mater. Med. 2006, 31, 397–400. [Google Scholar]
- Maldini, M.; Montora, P.; Hamed, A.; Mahalel, U.; Oleszek, W.; Stochmal, A.; Piacente, S. Strong antioxidant phenolics from Acacia nilotica: Profiling by ESI-MS and qualitative-quantitative determination by LC-ESI-MS. J. Pharm. Biol. Anal. 2011, 56, 228–239. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Yang, C.R. Chemical constituents of crude green tea, the material of Pu-er Tea in Yunnan. Acta Bot. Yunnanica 2000, 22, 343–350. [Google Scholar]
- Yue, J.M.; Zhao, Y.; Zhao, Q.S.; Wu, H.M.; Xu, J.F.; Lin, Z.W.; Sun, H.D. A novel compound from Ceratostigma Minus. Chin. Chem. Lett. 1998, 9, 647–649. [Google Scholar]
- Beigel, J.H.; Farrar, J.; Han, A.M.; Hayden, F.G.; Hyer, R.; de Jong, M.D.; Lochindarat, S.; Nguyen, T.K.; Nguyen, T.H.; Tran, T.H.; et al. Avian influenza A (H5N1) infection in humans. N. Engl. J. Med. 2005, 353, 1374–1385. [Google Scholar] [CrossRef]
- De Jong, M.D.; Simmons, C.P.; Thanh, T.T.; Hien, V.M.; Smith, G.J.; Chau, T.N.; Hoang, D.M.; Chau, N.V.; Khanh, T.H.; Dong, V.C.; et al. Fatal outcome of human influenza A (H5N1) is associated with high viral load and hypercytokinemia. Nat. Med. 2006, 12, 1203–1207. [Google Scholar] [CrossRef]
- Guo, X.Y.; Wang, N.L.; Bo, L.; Li, Y.H.; Xu, Q.; Yao, X.S. Chemical constituents from Pithecellobium clypearia and their effects on Tlymphocytes proliferation. J. Chin. Pharm. Sci. 2007, 16, 208–213. [Google Scholar]
- Zu, M.; Yang, F.; Zhou, W.; Liu, A.; Du, G.; Zheng, L. In vitro anti-influenza virus and anti-inflammatory activities of theaflavin derivatives. Antivir. Res. 2012, 94, 217–224. [Google Scholar] [CrossRef]
- Geiler, J.; Michaelis, M.; Naczk, P.; Leutz, A.; Langer, K.; Doerr, H.W.; Cinatl, J., Jr. N-acetyl-l-cysteine (NAC) inhibits virus replication and expression of proinflammatory molecules in A549 cells infected with highly pathogenic H5N1 influenza A virus. Biochem. Pharmacol. 2010, 79, 413–420. [Google Scholar] [CrossRef]
- Jeffrey, J.A.; Pamela, E.O.; Sunil, V.K.; Robert, C.B.; Jeffrey, C.G. A convenient large-scale synthesis of 5-methoxyflavone and its application to analog preparation. J. Org. Chem. 1993, 68, 7903–7905. [Google Scholar]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef]
- Liu, A.; Wang, H.; Lee, S.M.Y.; Wang, Y.; Du, G. Structure-activity relationship of flavonoids as influenza virus neuraminidase inhibitors and their in vitro anti-viral activities. Bioorg. Med. Chem. 2008, 16, 7141–7147. [Google Scholar] [CrossRef]
- Liu, A.; Yang, F.; Zhu, M.; Zhou, D.; Lin, M.; Lee, S.M.Y.; Wang, Y.; Du, G. In vitro anti-influenza viral activities of stilbenoids from the lianas of Gnetum pendulum. Planta Med. 2010, 76, 1874–1876. [Google Scholar] [CrossRef]
- Liu, A.; Liu, B.; Qin, H.; Lee, S.M.Y.; Wang, Y.; Du, G. Anti-influenza virus activities of flavonoids from the medicinal plant Elsholtzia rugulosa. Planta Med. 2008, 74, 847–851. [Google Scholar] [CrossRef]
- Ulrike, G.; Michaela, S.; Susanne, G.; Johannes, K.; Klaus, L.; Judith, R. Influenza neuraminidase: A druggable target for natural products. Nat. Prod. Rep. 2012, 29, 11–36. [Google Scholar] [CrossRef]
- Tumpey, T.M.; Basler, C.F.; Aguilar, P.V.; Zeng, H.; Solorzano, A.; Swayne, D.E. Characterization of the reconstructed 1918 Spanish influenza pandemic virus. Science 2005, 310, 77–80. [Google Scholar] [CrossRef]
- Sladkova, T.; Kostolansky, F. The role of cytokines in the immune response to influenza A virus infection. Acta Virol. 2006, 50, 151–162. [Google Scholar]
- Uchide, N.; Ohyama, K.; Toyoda, H. Current and future anti-influenza virus drugs. Open Antimicrob. Agents J. 2010, 2, 34–48. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds 1–4 are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kang, J.; Liu, C.; Wang, H.; Li, B.; Li, C.; Chen, R.; Liu, A. Studies on the Bioactive Flavonoids Isolated from Pithecellobium clypearia Benth. Molecules 2014, 19, 4479-4490. https://doi.org/10.3390/molecules19044479
Kang J, Liu C, Wang H, Li B, Li C, Chen R, Liu A. Studies on the Bioactive Flavonoids Isolated from Pithecellobium clypearia Benth. Molecules. 2014; 19(4):4479-4490. https://doi.org/10.3390/molecules19044479
Chicago/Turabian StyleKang, Jie, Chao Liu, Hongqing Wang, Baoming Li, Chao Li, Ruoyun Chen, and Ailin Liu. 2014. "Studies on the Bioactive Flavonoids Isolated from Pithecellobium clypearia Benth" Molecules 19, no. 4: 4479-4490. https://doi.org/10.3390/molecules19044479
APA StyleKang, J., Liu, C., Wang, H., Li, B., Li, C., Chen, R., & Liu, A. (2014). Studies on the Bioactive Flavonoids Isolated from Pithecellobium clypearia Benth. Molecules, 19(4), 4479-4490. https://doi.org/10.3390/molecules19044479