Bufalin Alters Gene Expressions Associated DNA Damage, Cell Cycle, and Apoptosis in Human Lung Cancer NCI-H460 Cells in Vitro


Abstract
:1. Introduction
2. Results and Discussion
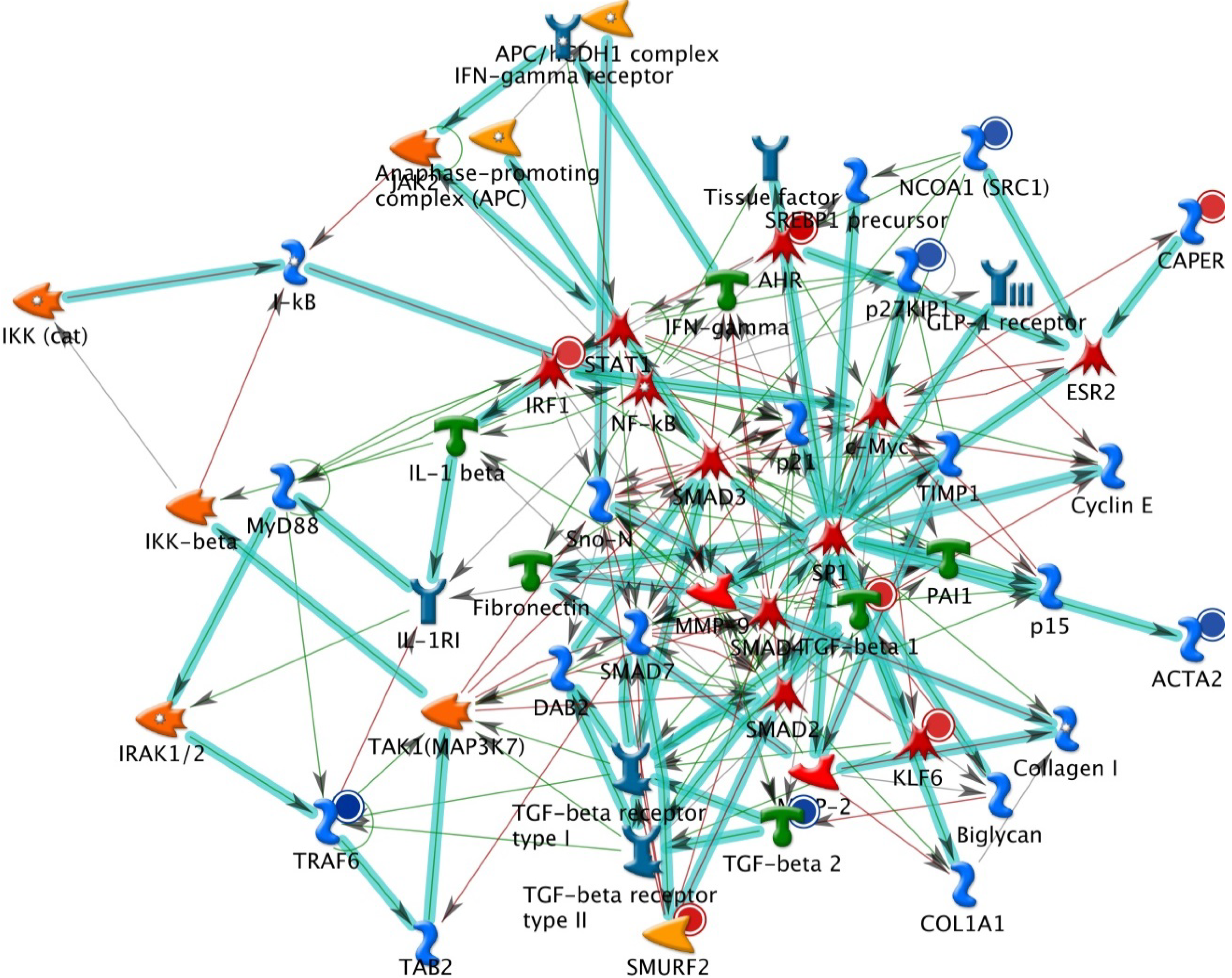
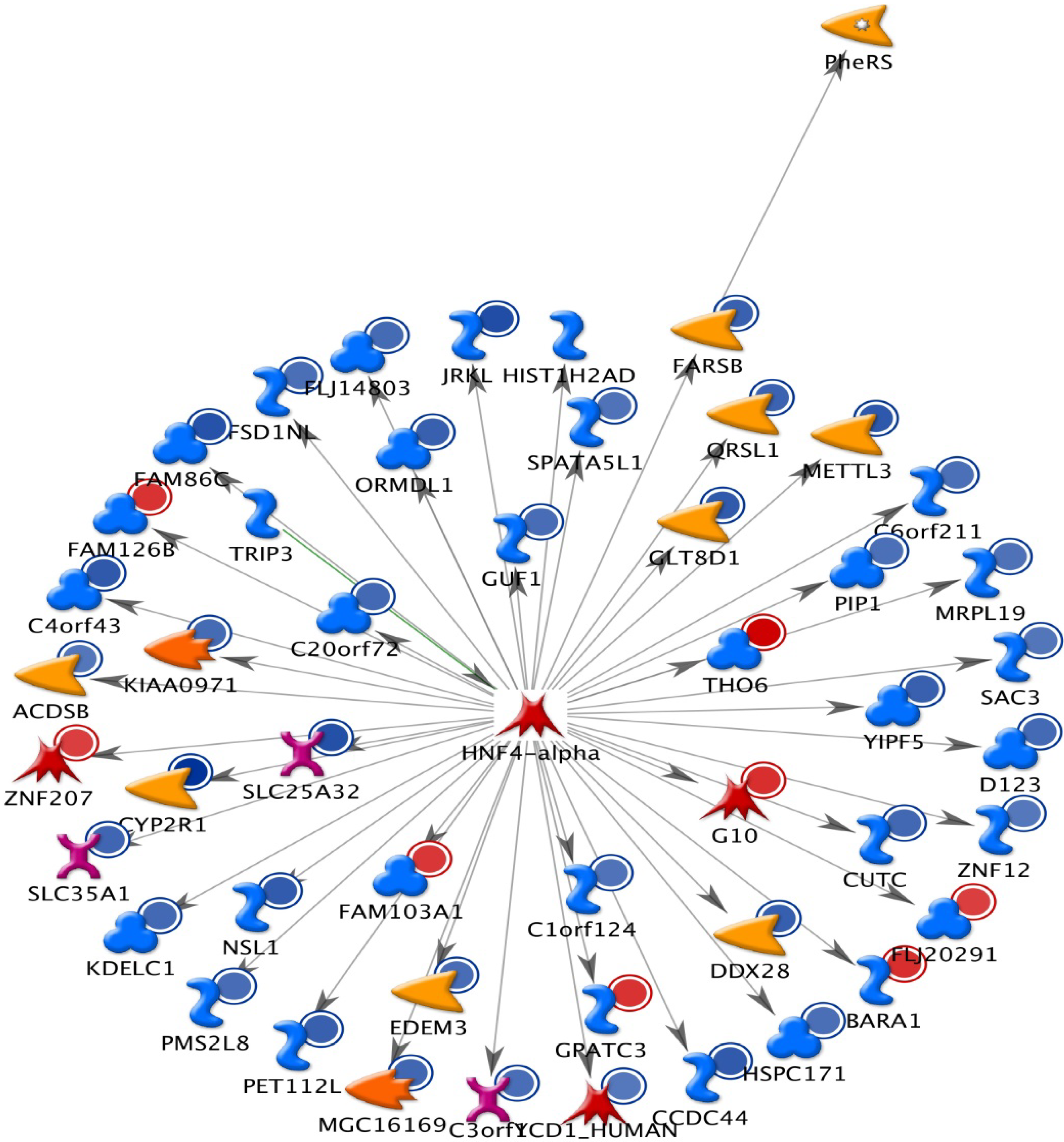
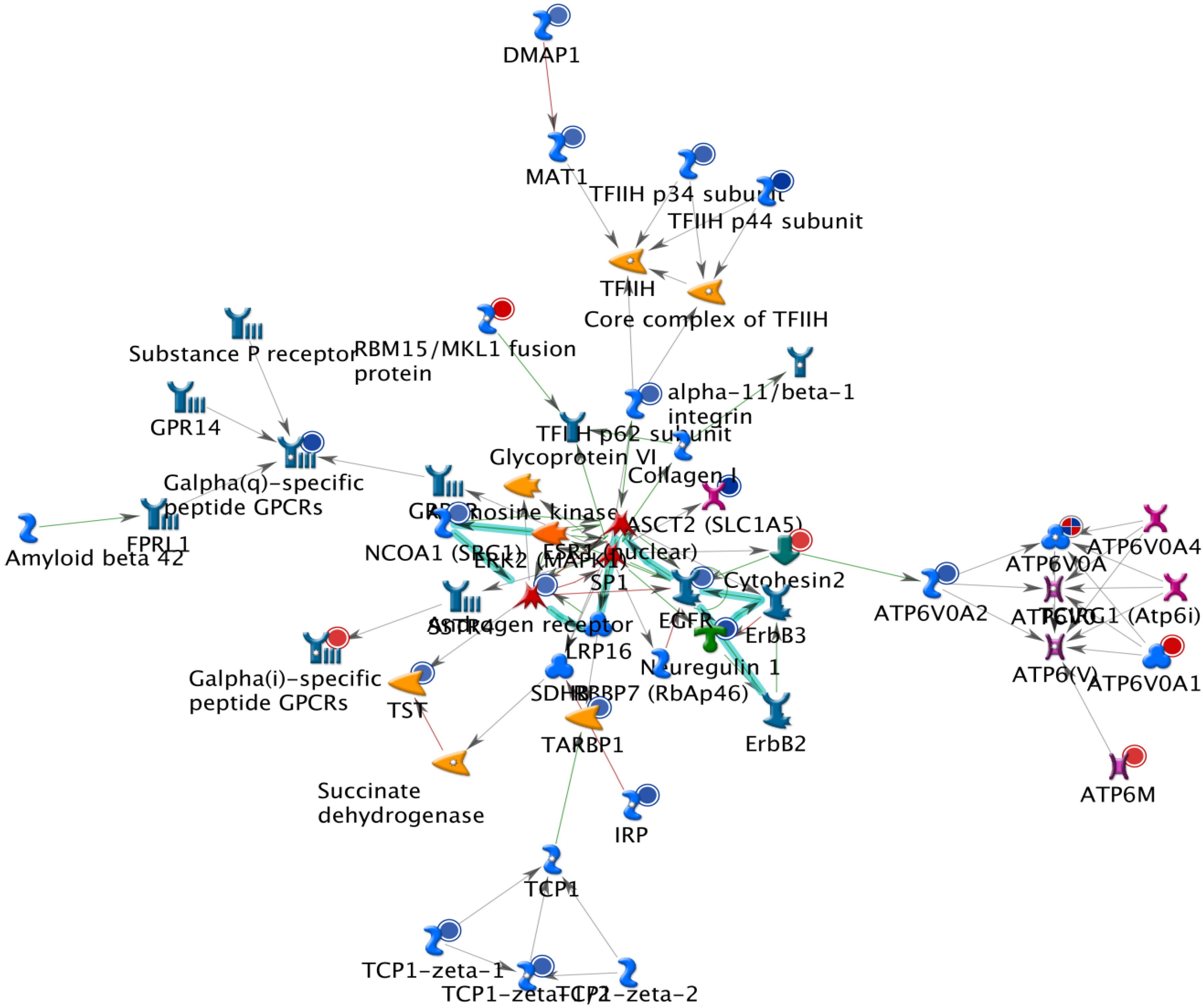
2.1. The Up-Regulated and Down-Regulated Gene Expression in H460 Exposed to Bufalin

{kind=link}
{kind=link}
{kind=link}
| Fold Change | Number of Genes |
|---|---|
| ≥20 | 6 |
| ≥10 and <20 | 21 |
| ≥5 and <10 | 59 |
| ≥4 and <5 | 53 |
| ≥3 and <4 | 118 |
| ≥2 and <3 | 488 |
| >−3 and ≤−2 | 1215 |
| >−4 and ≤−3 | 348 |
| >−5 and ≤−4 | 99 |
| >−6 and ≤−5 | 57 |
| >−10 and ≤−6 | 42 |
| <−10 | 11 |
2.2. GeneGo Analysis Program from Bufalin Treated H460 Cells Demonstrated the Top Alteration in Gene Expression Scored by the Number of Pathway Networks
| Fold change | Gene Symbol | mRNA Description |
|---|---|---|
| 66.33 | EGR1 | early growth response 1 |
| 26.16 | EGR2 | GTP binding protein overexpressed in skeletal muscle |
| 24.09 | EGR3 | pyruvate dehydrogenase kinase, isozyme 4 |
| 23.48 | EGR4 | NOTCH-regulated ankyrin repeat protein |
| 23.10 | EGR5 | histone cluster 1, H4d |
| 20.63 | EGR6 | FBJ murine osteosarcoma viral oncogene homolog |
| 5.52 | BTG | BTG family, member 2 |
| 5.51 | CASP9 | caspase 9, apoptosis-related cysteine peptidase |
| 4.54 | CRY2 | cryptochrome 2 (photolyase-like) |
| 4.36 | AEN | apoptosis enhancing nuclease |
| 3.72 | GADD45B | growth arrest and DNA-damage-inducible, beta |
| 2.75 | THAP1 | THAP domain containing, apoptosis associated protein 1 |
| 2.02 | FLJ43315 | similar to Asparagine synthetase [glutamine-hydrolyzing] (Glutamine-dependent asparagine synthetase) (TS11 cell cycle control protein) |
| −2.11 | CCPG1 | cell cycle progression 1 |
| −2.22 | MDC1 | mediator of DNA-damage checkpoint 1 |
| −2.24 | CCAR1 | cell division cycle and apoptosis regulator 1 |
| −2.25 | SMC3 | structural maintenance of chromosomes 3 |
| −2.52 | DDIT4 | DNA-damage-inducible transcript 4 |
| −2.71 | CDCA7L | cell division cycle associated 7-like |
| −3.18 | DCLRE1C | DNA cross-link repair 1C (PSO2 homolog, S. cerevisiae) |
| −3.93 | UIMC1 | ubiquitin interaction motif containing 1 |
| −5.04 | DCLRE1A | DNA cross-link repair 1A (PSO2 homolog, S. cerevisiae) |
| −10.55 | CCND2 | cyclin D2 |
| −10.57 | C5orf33 | chromosome 5 open reading frame 33 |
| −11.08 | GPR65 | G protein-coupled receptor 65 |
| −11.38 | PRICKLE1 | prickle homolog 1 (Drosophila) |
| −11.93 | ORC5L | origin recognition complex, subunit 5-like (yeast) |
| −12.02 | AHNAK2 | AHNAK nucleoprotein 2 |
| −12.24 | ICT1 | immature colon carcinoma transcript 1 |
| −12.61 | KRT19 | keratin 19 |
| −15.54 | OCLN | occludin |
| −27.38 | MIR1977 | microRNA 1977 |
2.3. Discussion



3. Experimental
3.1. Chemicals and Reagents
3.2. Lung Cancer Cells
3.3. cDNA Microarray Assay for Genes Expression in H460 Cells after Exposure to Bufalin
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2013. CA Cancer J. Clin. 2013, 63, 11–30. [Google Scholar] [CrossRef]
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Global cancer statistics, 2002. CA Cancer J. Clin. 2005, 55, 74–108. [Google Scholar] [CrossRef]
- Non-small Cell Lung Cancer Collaborative Group. Chemotherapy in non-small cell lung cancer: A meta-analysis using updated data on individual patients from 52 randomised clinical trials. BMJ 1995, 311, 899–909. [CrossRef]
- Sandler, A.; Gray, R.; Perry, M.C.; Brahmer, J.; Schiller, J.H.; Dowlati, A.; Lilenbaum, R.; Johnson, D.H. Paclitaxel-carboplatin alone or with bevacizumab for non-small-cell lung cancer. N. Engl. J. Med. 2006, 355, 2542–2550. [Google Scholar] [CrossRef]
- Zhu, Z.; Sun, H.; Ma, G.; Wang, Z.; Li, E.; Liu, Y.; Liu, Y. Bufalin induces lung cancer cell apoptosis via the inhibition of PI3K/Akt pathway. Int. J. Mol. Sci. 2012, 13, 2025–2035. [Google Scholar] [CrossRef]
- Han, K.Q.; Huang, G.; Gu, W.; Su, Y.H.; Huang, X.Q.; Ling, C.Q. Anti-tumor activities and apoptosis-regulated mechanisms of bufalin on the orthotopic transplantation tumor model of human hepatocellular carcinoma in nude mice. World J. Gastroenterol. 2007, 13, 3374–3379. [Google Scholar]
- Chen, A.; Yu, J.; Zhang, L.; Sun, Y.; Zhang, Y.; Guo, H.; Zhou, Y.; Mitchelson, K.; Cheng, J. Microarray and biochemical analysis of bufalin-induced apoptosis of HL-60 Cells. Biotechnol. Lett. 2009, 31, 487–494. [Google Scholar] [CrossRef]
- Chen, H.Y.; Lu, H.F.; Yang, J.S.; Kuo, S.C.; Lo, C.; Yang, M.D.; Chiu, T.H.; Chueh, F.S.; Ho, H.C.; Ko, Y.C.; et al. The novel quinolone CHM-1 induces DNA damage and inhibits DNA repair gene expressions in a human osterogenic sarcoma cell line. Anticancer Res. 2010, 30, 4187–4192. [Google Scholar]
- Yu, C.H.; Kan, S.F.; Pu, H.F.; Jea Chien, E.; Wang, P.S. Apoptotic signaling in bufalin- and cinobufagin-treated androgen-dependent and -independent human prostate cancer cells. Cancer Sci. 2008, 99, 2467–2476. [Google Scholar] [CrossRef]
- Takai, N.; Kira, N.; Ishii, T.; Yoshida, T.; Nishida, M.; Nishida, Y.; Nasu, K.; Narahara, H. Bufalin, a traditional oriental medicine, induces apoptosis in human cancer cells. Asian Pac. J. Cancer Prev. 2012, 13, 399–402. [Google Scholar] [CrossRef]
- Huang, H.; Cao, Y.; Wei, W.; Liu, W.; Lu, S.Y.; Chen, Y.B.; Wang, Y.; Yan, H.; Wu, Y.L. Targeting poly (ADP-ribose) polymerase partially contributes to bufalin-induced cell death in multiple myeloma cells. PLoS One 2013, 8, e66130. [Google Scholar]
- Nasu, K.; Nishida, M.; Ueda, T.; Takai, N.; Bing, S.; Narahara, H.; Miyakawa, I. Bufalin induces apoptosis and the G0/G1 cell cycle arrest of endometriotic stromal cells: A promising agent for the treatment of endometriosis. Mol. Hum. Reprod. 2005, 11, 817–823. [Google Scholar]
- Cao, H.; Shibayama-Imazu, T.; Masuda, Y.; Shinki, T.; Nakajo, S.; Nakaya, K. Involvement of Tiam1 in apoptosis induced by bufalin in HeLa cells. Anticancer Res. 2007, 27, 245–249. [Google Scholar]
- Masuda, Y.; Kawazoe, N.; Nakajo, S.; Yoshida, T.; Kuroiwa, Y.; Nakaya, K. Bufalin induces apoptosis and influences the expression of apoptosis-related genes in human leukemia cells. Leuk. Res. 1995, 19, 549–556. [Google Scholar] [CrossRef]
- Lee, J.H.; Koh, Y.A.; Cho, C.K.; Lee, S.J.; Lee, Y.S.; Bae, S. Identification of a novel ionizing radiation-induced nuclease, AEN, and its functional characterization in apoptosis. Biochem. Biophys. Res. Commun. 2005, 337, 39–47. [Google Scholar] [CrossRef]
- Kawase, T.; Ichikawa, H.; Ohta, T.; Nozaki, N.; Tashiro, F.; Ohki, R.; Taya, Y. p53 target gene AEN is a nuclear exonuclease required for p53-dependent apoptosis. Oncogene 2008, 27, 3797–3810. [Google Scholar] [CrossRef]
- Zumbrun, S.D.; Hoffman, B.; Liebermann, D.A. Distinct mechanisms are utilized to induce stress sensor gadd45b by different stress stimuli. J. Cell. Biochem. 2009, 108, 1220–1231. [Google Scholar] [CrossRef]
- Vairapandi, M.; Balliet, A.G.; Hoffman, B.; Liebermann, D.A. GADD45b and GADD45g are cdc2/cyclinB1 kinase inhibitors with a role in S and G2/M cell cycle checkpoints induced by genotoxic stress. J. Cell. Physiol. 2002, 192, 327–38. [Google Scholar] [CrossRef]
- Zenmyo, M.; Tanimoto, A.; Sakakima, H.; Yokouchi, M.; Nagano, S.; Yamamoto, T.; Ishido, Y.; Komiya, S.; Ijiri, K. Gadd45beta expression in chondrosarcoma: A pilot study for diagnostic and biological implications in histological grading. Diagn. Pathol. 2010, 5, 69–73. [Google Scholar] [CrossRef]
- Richie, C.T.; Peterson, C.; Lu, T.; Hittelman, W.N.; Carpenter, P.B.; Legerski, R.J. hSnm1 colocalizes and physically associates with 53BP1 before and after DNA damage. Mol. Cell. Biol. 2002, 22, 8635–8647. [Google Scholar] [CrossRef]
- Akhter, S.; Richie, C.T.; Deng, J.M.; Brey, E.; Zhang, X.; Patrick, C., Jr.; Behringer, R.R.; Legerski, R.J. Deficiency in SNM1 abolishes an early mitotic checkpoint induced by spindle stress. Mol. Cell. Biol. 2004, 24, 10448–10455. [Google Scholar] [CrossRef]
- Koyama-Nasu, R.; Nasu-Nishimura, Y.; Todo, T.; Ino, Y.; Saito, N.; Aburatani, H.; Funato, K.; Echizen, K.; Sugano, H.; Haruta, R.; et al. The critical role of cyclin D2 in cell cycle progression and tumorigenicity of glioblastoma stem cells. Oncogene 2013, 32, 3840–3845. [Google Scholar] [CrossRef]
- Igawa, T.; Sato, Y.; Takata, K.; Fushimi, S.; Tamura, M.; Nakamura, N.; Maeda, Y.; Orita, Y.; Tanimoto, M.; Yoshino, T. Cyclin D2 is overexpressed in proliferation centers of chronic lymphocytic leukemia/small lymphocytic lymphoma. Cancer Sci. 2011, 102, 2103–2107. [Google Scholar] [CrossRef]
- Liu, S.C.; Bassi, D.E.; Zhang, S.Y.; Holoran, D.; Conti, C.J.; Klein-Szanto, A.J. Overexpression of cyclin D2 is associated with increased in vivo invasiveness of human squamous carcinoma cells. Mol. Carcinog. 2002, 34, 131–139. [Google Scholar] [CrossRef]
- Chang, Y.-M.; Velmurugan, B.K.; Kuo, W.-W.; Chen, Y.-S.; Ho, T.-J.; Tsai, C.-T.; Ye, C.-X.; Tsai, C.-H.; Tsai, F.-J.; Huang, C.-Y. Inhibitory effect of alpinate Oxyphyllae fructus extracts on Ang II-induced cardiac pathological remodeling-related pathways in H9c2 cardiomyoblast cells. Biomedicine 2013, 3, 148–152. [Google Scholar] [CrossRef]
- Lin, M.-C.; Tsai, S.-Y.; Wang, F.-Y.; Liu, F.-H.; Syu, J.-N.; Tang, F.-Y. Leptin induces cell invasion and the upregulation of matrilysin in human colon cancer cells. Biomedicine 2013, 3, 174–180. [Google Scholar] [CrossRef]
- Leung, Y.M.; Wong, K.L.; Chen, S.W.; Lu, D.Y.; Kuo, C.S.; Chen, Y.R.; Chen, Y.W.; Cheng, T.H. Down-regulation of voltage-gated Ca2+ channels in Ca2+ store-depleted rat insulinoma RINm5F cells. Biomedicine 2013, 3, 130–139. [Google Scholar] [CrossRef]
- Wu, R.S.; Liu, K.C.; Tang, N.Y.; Chung, H.K.; Ip, S.W.; Yang, J.S.; Chung, J.G. cDNA microarray analysis of the gene expression of murine leukemia RAW 264.7 cells after exposure to propofol. Environ. Toxicol. 2013, 28, 471–478. [Google Scholar] [CrossRef]
- Gardina, P.J.; Clark, T.A.; Shimada, B.; Staples, M.K.; Yang, Q.; Veitch, J.; Schweitzer, A.; Awad, T.; Sugnet, C.; Dee, S.; et al. Alternative splicing and differential gene expression in colon cancer detected by a whole genome exon array. BMC Genomics 2006, 7, 325. [Google Scholar] [CrossRef]
- Douglas, D.; Hsu, J.H.; Hung, L.; Cooper, A.; Abdueva, D.; van Doorninck, J.; Peng, G.; Shimada, H.; Triche, T.J.; Lawlor, E.R. BMI-1 promotes ewing sarcoma tumorigenicity independent of CDKN2A repression. Cancer Res. 2008, 68, 6507–6515. [Google Scholar] [CrossRef]
- Jacobs, A.T.; Marnett, L.J. HSF1-mediated BAG3 expression attenuates apoptosis in 4-hydroxynonenal-treated colon cancer cells via stabilization of anti-apoptotic Bcl-2 proteins. J. Biol. Chem. 2009, 284, 9176–9183. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, S.-H.; Hsiao, Y.-T.; Chen, J.-C.; Lin, J.-H.; Hsu, S.-C.; Hsia, T.-C.; Yang, S.-T.; Hsu, W.-H.; Chung, J.-G. Bufalin Alters Gene Expressions Associated DNA Damage, Cell Cycle, and Apoptosis in Human Lung Cancer NCI-H460 Cells in Vitro. Molecules 2014, 19, 6047-6057. https://doi.org/10.3390/molecules19056047
Wu S-H, Hsiao Y-T, Chen J-C, Lin J-H, Hsu S-C, Hsia T-C, Yang S-T, Hsu W-H, Chung J-G. Bufalin Alters Gene Expressions Associated DNA Damage, Cell Cycle, and Apoptosis in Human Lung Cancer NCI-H460 Cells in Vitro. Molecules. 2014; 19(5):6047-6057. https://doi.org/10.3390/molecules19056047
Chicago/Turabian StyleWu, Shin-Hwar, Yung-Ting Hsiao, Jaw-Chyum Chen, Ju-Hwa Lin, Shu-Chun Hsu, Te-Chun Hsia, Su-Tso Yang, Wu-Huei Hsu, and Jing-Gung Chung. 2014. "Bufalin Alters Gene Expressions Associated DNA Damage, Cell Cycle, and Apoptosis in Human Lung Cancer NCI-H460 Cells in Vitro" Molecules 19, no. 5: 6047-6057. https://doi.org/10.3390/molecules19056047