CopA3 Peptide Prevents Ultraviolet-Induced Inhibition of Type-I Procollagen and Induction of Matrix Metalloproteinase-1 in Human Skin Fibroblasts

Abstract
:1. Introduction
2. Results and Discussion
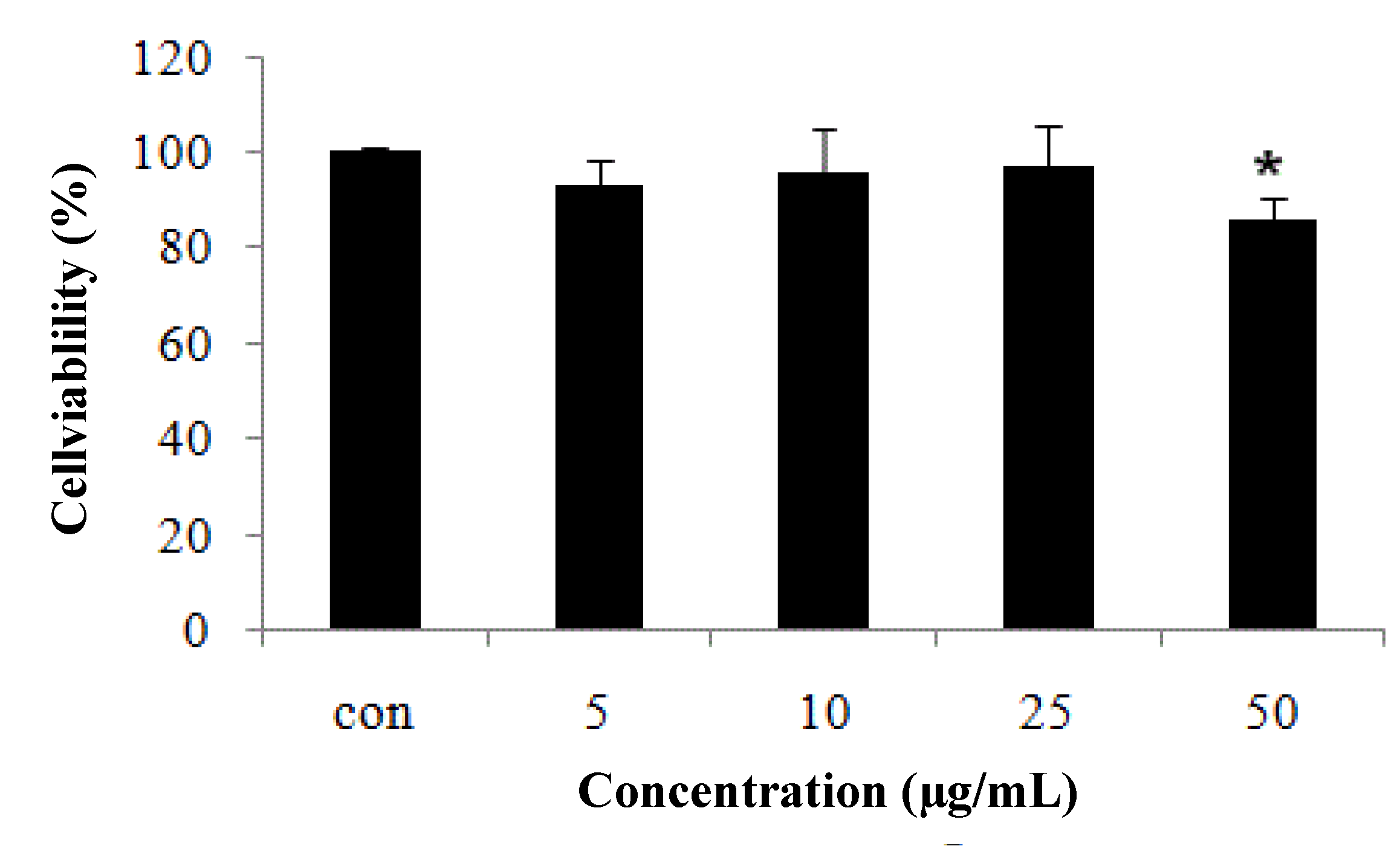
2.1. CCD-986sk Fibroblast Cell Viability Measured by MTT Assay
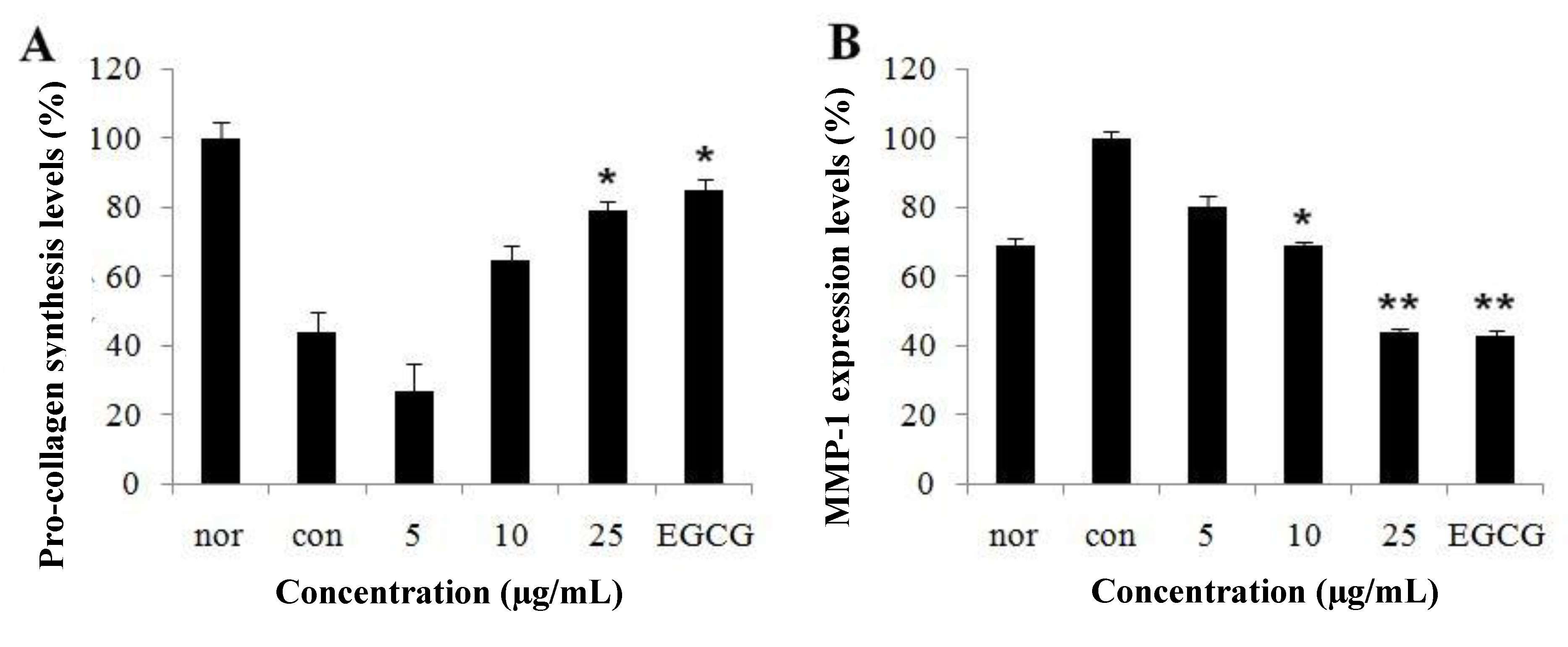
2.2. Effect of CopA3 on Type-I Procollagen Synthesis and Matrix Metalloproteinase-1 Levels


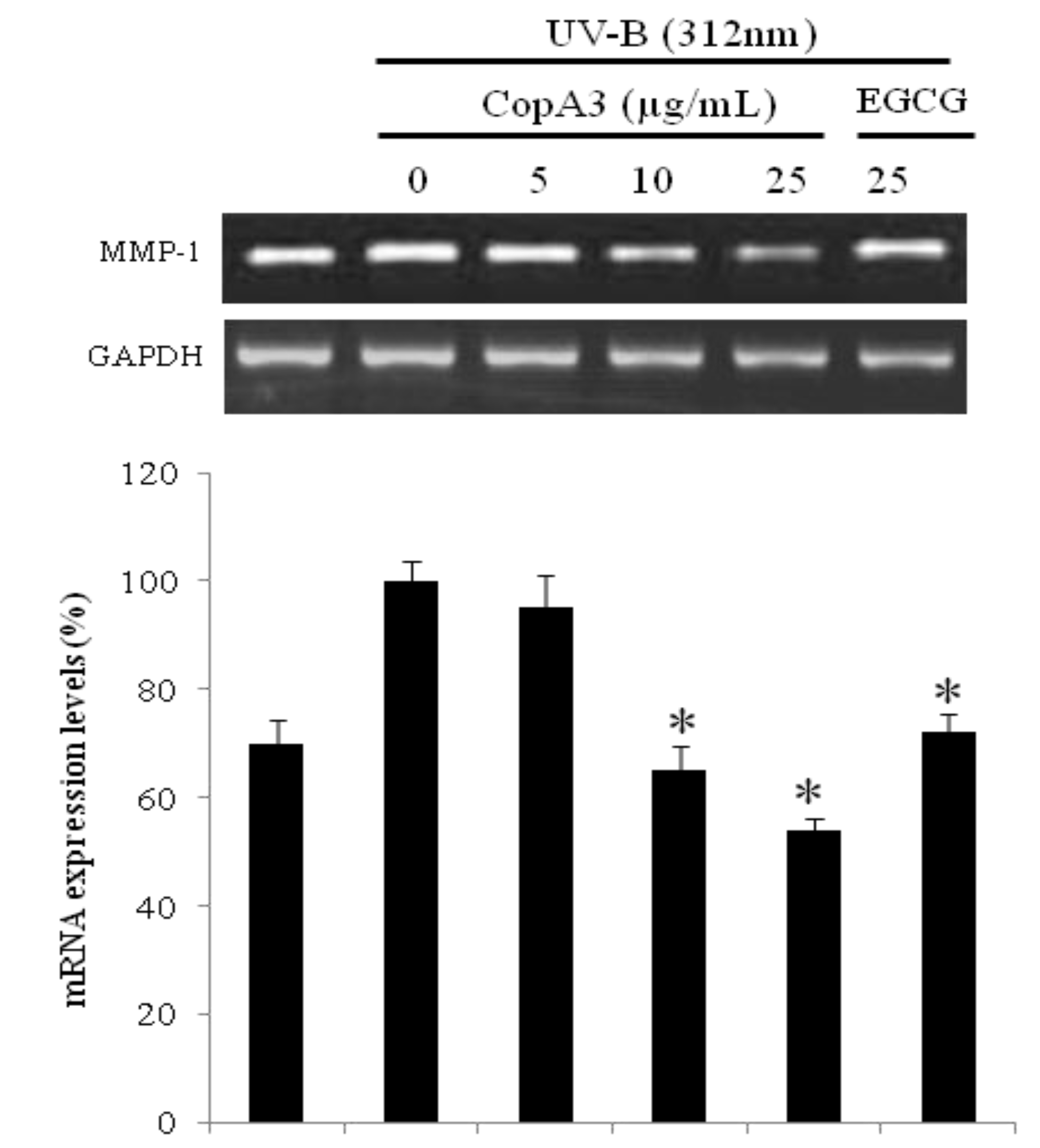
2.3. mRNA of MMP-1 was Decreased in CCD-986sk Cells by CopA3

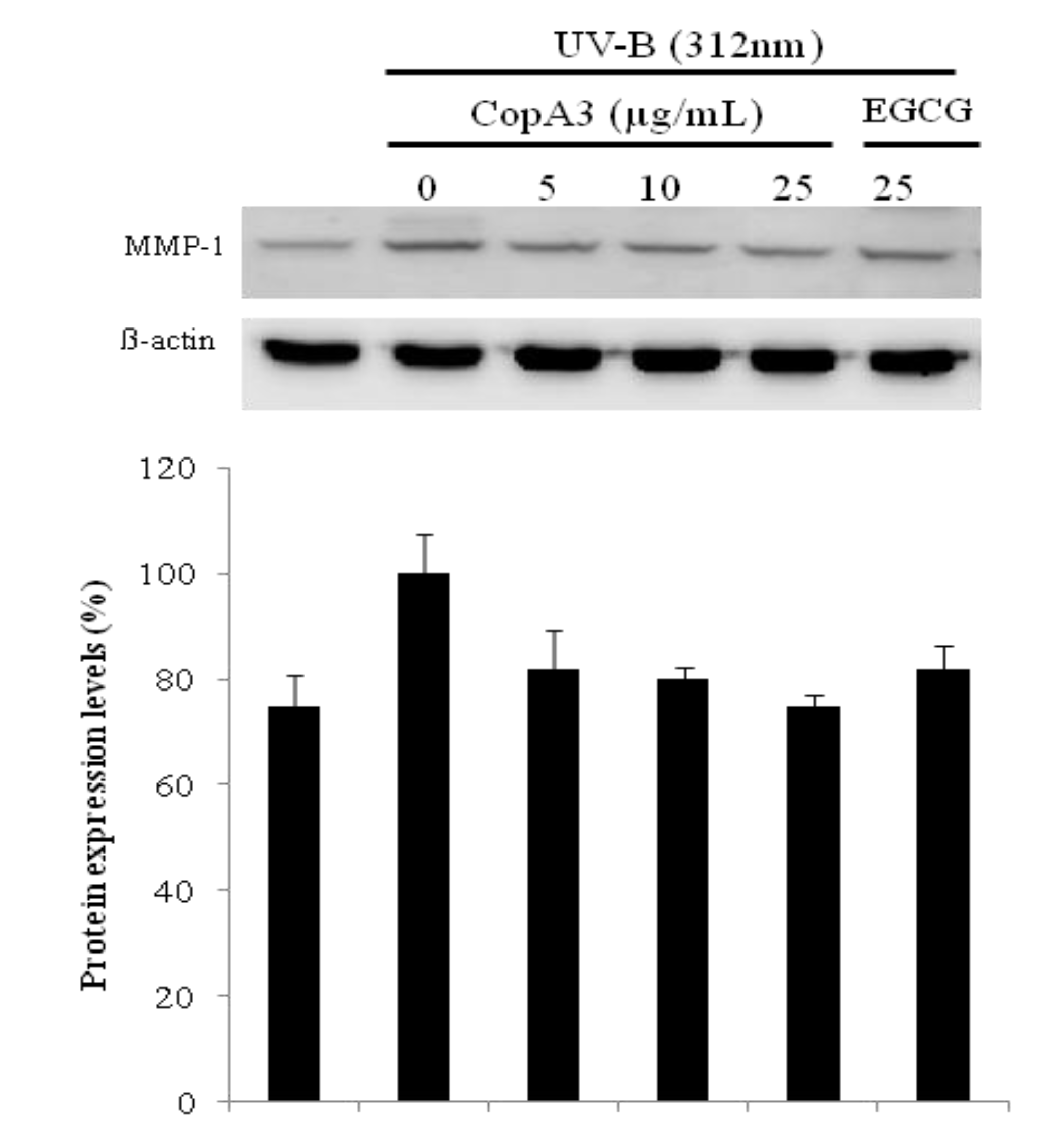
2.4. UVB Induced Aging Related MMP-1 Protein Expression was Suppressed by CopA3

3. Experimental
3.1. CopA3 (Disulfide Dimer) Synthesis, Dimer Peptide Structure Determination
3.2. Cell Culture
3.3. Cell Viability
3.4. Determination of MMP-1 and Type-I Procollagen
3.5. cDNA Synthesis and RT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5' → 3') |
|---|---|---|
| MMP-1 | Forward | AGC GTG TGA CAG TAA GCT AA |
| Reverse | GTT TTC CTC AGA AAG AGC AGC AT | |
| β-actin | Forward | ATT GTT GCC ATC AAT GAC CC |
| Reverse | AGT AGA GGC AGG GAT GAT |
3.6. Western Blotting
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fisher, G.J.; Voorhees, J.J. Molecular mechanisms of photoaging and its prevention by retinoic acid: Ultraviolet irradiation induces MAP kinase signal transduction cascades that induce Ap-1-regulated matrix metalloproteinases that degrade human skin in vivo. J. Invest. Derm. Symp. P. 1998, 3, 61–68. [Google Scholar]
- Seo, J.E.; Kim, S.; Shin, M.H.; Kim, M.S.; Eun, H.C.; Park, C.H.; Chung, J.H. Ultraviolet irradiation induces thrombospondin-1 which attenuates type-I procollagen downregulation in human dermal fibroblasts. J. Derm. Sci. 2010, 59, 16–24. [Google Scholar] [CrossRef]
- Yun, H.J.; Yoo, W.H.; Han, M.K.; Lee, Y.R.; Kim, J.S.; Lee, S.I. Epigallocatechin-3-gallate suppresses TNF-alpha-induced production of MMP-1 and -3 in rheumatoid arthritis synovial fibroblasts. Rheumatol. Int. 2008, 29, 23–29. [Google Scholar] [CrossRef]
- Kim, D.H.; Moon, Y.S.; An, B.J.; Son, J.H. Potent anti-aging activity of Aruncus dioicus, a native plant of Ulleung-do, South Korea, in CCD-986sk fibroblasts via suppression of matrix metalloproteinases. J. Nat. Med. 2012, 66, 631–636. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Xu, Y.; Voorhees, J.J.; Fisher, G.J. UV irradiation induces Snail expression by AP-1 dependent mechanism in human skin keratinocytes. J. Derm. Sci. 2010, 60, 105–113. [Google Scholar] [CrossRef]
- Kang, B.R.; Kim, H.; Nam, S.H.; Yun, E.Y.; Kim, S.R.; Ahn, M.Y.; Chung, J.S.; Hwang, J.S. CopA3 peptide from Copris tripartitus induces apoptosis in human leukemia cells via a caspase-independent pathway. Biochem. Mol. Biol. Rep. 2012, 45, 85–90. [Google Scholar]
- Hwang, J.S.; Lee, J.Y.; Kim, Y.J.; Bang, H.S.; Yun, E.Y.; Kim, S.R. Isolation and characterization of a defensin-like peptide (coprisin) from the dung beetle, Copris tripartitus. Int. J. Pept. 2009. [Google Scholar] [CrossRef]
- Kang, J.K.; Hwang, J.S.; Nam, H.J.; Ahn, K.J.; Seok, H.; Kim, S.K. The insect peptide coprisin prevents Clostridium difficile-mediated acute inflammation and mucosal damage through selective antimicrobial activity. Antimicrob. Agents Chmother. 2011, 55, 4850–4857. [Google Scholar] [CrossRef]
- Nam, H.J.; Oh, A.R.; Nam, S.T.; Kang, J.K.; Chang, J.S.; Kim, D.H. The insect peptide CopA3 inhibits lipopolysaccharide-induced macrophage activation. J. Pept. Sci. 2012, 18, 650–656. [Google Scholar] [CrossRef]
- Shin, M.H.; Rhie, G.E.; Kim, Y.K.; Park, C.H.; Cho, K.H.; Kim, K.H.; Chung, J.H. H2O2 accumulation by catalase reduction changes MAP kinase signaling in aged human skin in vivo. J. Invest. Derm. 2005, 125, 221–229. [Google Scholar]
- Tsoyi, K.; Park, H.B.; Kim, Y.M.; Chung, J.I.; Shin, S.C.; Lee, W.S. Anthocyanins from black soybean seed coats inhibit UVB-induced inflammatory cylooxygenase-2 gene expression and PGE2 production through regulation of the nuclear factor-κB and phosphatidylinositol 3-kinase/Akt pathway. J. Agric. Food Chem. 2008, 56, 8969–8974. [Google Scholar] [CrossRef]
- Wang, H.; Mehta, J.L.; Chen, K.; Zhang, X.; Li, D. Human urotensin II modulates collagen synthesis and the expression of MMP-1 in human endothelial cells. J. Cardiovasc. Pharmacol. 2004, 44, 577–581. [Google Scholar] [CrossRef]
- Chen, C.L.; Liou, S.F.; Chen, S.J.; Shih, M.F. Protective effects of Chlorella-derived peptide on UVB-induced production of MMP-1 and degradation of procollagen genes in human skin fibroblasts. Regul. Toxicol. Pharmacol. 2011, 60, 112–119. [Google Scholar] [CrossRef]
- Quan, T.; Qin, Z.; Xu, Y.; He, T.; Kang, S.; Voorhees, J.J. Ultraviolet irradiation induces CYR61/CCN1, a mediator of collagen homeostasis, through activation of transcription factor AP-1 in human skin fibroblasts. J. Invest. Derm. 2010, 130, 1679–1706. [Google Scholar]
- Sample Availability: Sample of the CopA3 peptide is not available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, D.-H.; Kim, H.-H.; Kim, H.-J.; Jung, H.-G.; Yu, J.-M.; Lee, E.-S.; Cho, Y.-H.; Kim, D.-I.; An, B.-J. CopA3 Peptide Prevents Ultraviolet-Induced Inhibition of Type-I Procollagen and Induction of Matrix Metalloproteinase-1 in Human Skin Fibroblasts. Molecules 2014, 19, 6407-6414. https://doi.org/10.3390/molecules19056407
Kim D-H, Kim H-H, Kim H-J, Jung H-G, Yu J-M, Lee E-S, Cho Y-H, Kim D-I, An B-J. CopA3 Peptide Prevents Ultraviolet-Induced Inhibition of Type-I Procollagen and Induction of Matrix Metalloproteinase-1 in Human Skin Fibroblasts. Molecules. 2014; 19(5):6407-6414. https://doi.org/10.3390/molecules19056407
Chicago/Turabian StyleKim, Dong-Hee, Han-Hyuk Kim, Hyeon-Jeong Kim, Hyun-Gug Jung, Jae-Myo Yu, Eun-Su Lee, Yong-Hun Cho, Dong-In Kim, and Bong-Jeun An. 2014. "CopA3 Peptide Prevents Ultraviolet-Induced Inhibition of Type-I Procollagen and Induction of Matrix Metalloproteinase-1 in Human Skin Fibroblasts" Molecules 19, no. 5: 6407-6414. https://doi.org/10.3390/molecules19056407