Complement Fixing Polysaccharides from Terminalia macroptera Root Bark, Stem Bark and Leaves

Abstract
:1. Introduction
2. Results and Discussion
2.1. Crude Extracts
2.1.1. Yields

{kind=link}
{kind=link}
{kind=link}
| 50WTRBH | 100WTRBH | 50WTSBH | 100WTSBH | 50WTLH | 100WTLH | |
|---|---|---|---|---|---|---|
| Ara a | 5.9 | 0.4 | 14.0 | 0.6 | 15.3 | 10.2 |
| Rha a | 11.8 | 0.6 | 9.5 | 1.3 | 7.7 | 2.4 |
| Xyl a | 2.1 | 0.3 | 4.7 | 0.7 | 3.3 | 2.9 |
| Man a | 1.0 | 0.2 | 0.6 | Trace | 2.0 | 7.8 |
| Gal a | 20.2 | 1.1 | 23.7 | 1.3 | 18.5 | 8.4 |
| Glc a | 36.5 | 97.4 | 23.3 | 92.1 | 14.8 | 49.8 |
| GlcA a | 1.0 | n.d. | 4.3 | Trace | 2.6 | Trace |
| GalA a | 21.4 | Trace | 19.9 | 4.0 | 35.8 | 18.4 |
| Yield (% w/w) b | 0.6 | 2.5 | 0.5 | 1.3 | 0.3 | 0.6 |
| Presence of starch | + | + | + | + | + | + |
2.1.2. Chemical Compositions
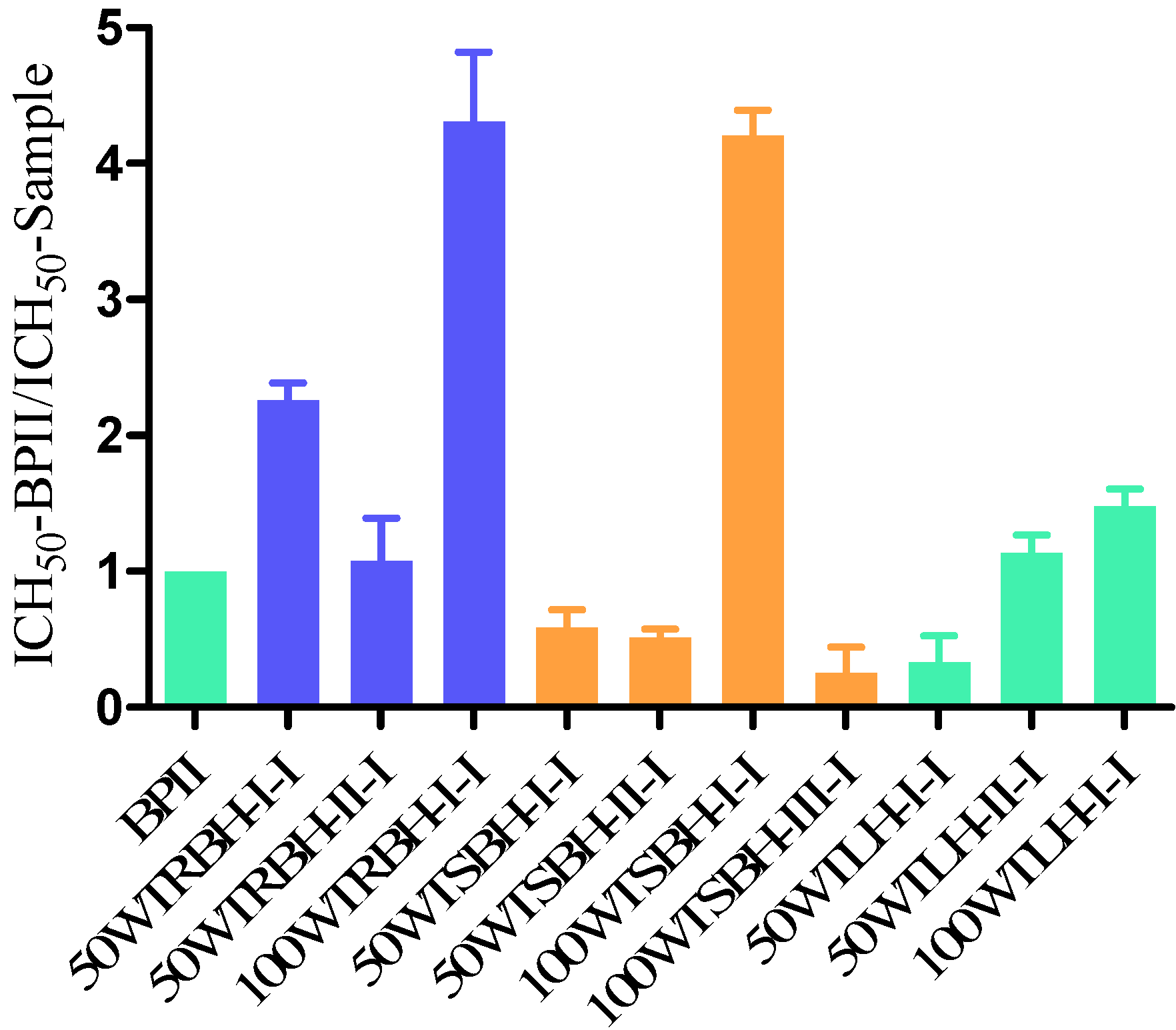
2.1.3. Complement Fixation Activity

2.2. Studies on Purified Polysaccharide Fractions
2.2.1. Yields
2.2.2. Chemical Compositions
2.2.3. Molecular Weight Distribution
2.2.4. Complement Fixation Activity
| 50WTRBH-I-I | 50WTRBH-II-I | 100WTRBH-I-I | 50WTSBH-I-I | 50WTSBH-II-I | 100WTSBH-I-I | 100WTSBH-III-I | 50WTLH-I-I | 50WTLH-II-I | 100WTLH-I-I | |
|---|---|---|---|---|---|---|---|---|---|---|
| Ara a | 19.6 | 7.1 | n.d. | 15.4 | 10.8 | 16.4 | 9.7 | 15.2 | 11.7 | 25.4 |
| Rha a | 3.5 | 29.1 | 23.6 | 4.3 | 18.4 | 4.3 | 14.4 | 2.3 | 12.5 | 3.1 |
| Xyl a | 6.4 | 2.3 | n.d. | 21.4 | 4.3 | 1.6 | 4.2 | 34.8 | 1.5 | 23.9 |
| Man a | 1.0 | 2.2 | 1.0 | 2.6 | 0.4 | 0.8 | n.d. | 2.7 | 0.3 | 4.1 |
| Gal a | 48.3 | 15.3 | 3.7 | 34.7 | 16.2 | 19.3 | 7.5 | 21.2 | 10.7 | 23.6 |
| Glc a | 2.1 | 1.6 | 25.8 | 3.6 | 2.7 | 19.0 | 9.5 | 7.1 | 2.2 | 7.2 |
| GlcA a | 2.1 | 3.1 | n.d. | 2.8 | 3.4 | 1.7 | 4.9 | 0.9 | 5.9 | 0.3 |
| GalA a | 17.0 | 39.2 | 45.9 | 15.1 | 43.6 | 36.9 | 49.9 | 15.8 | 55.2 | 12.4 |
| Yield (% w/w) b | 0.001 | 0.01 | 0.01 | 0.003 | 0.014 | 0.003 | 0.033 | 0.003 | 0.016 | 0.01 |
| Mw (kDa) | 136.1 | 115.9 | 491.8 | 19.8 | 23.2 | 98.7 | 44.2 | 19.8 | 220.3 | 136.1 |
| The Yariv test c | ++ | ++ | - | + | ++ | ++ | ++ | + | ++ | + |
| Presence of starch | - | - | + | - | - | + | + | + | - | + |
| Protein (% w/w) | 0.1 | n.d. | n.d. | 0.9 | n.d. | n.d. | 0.7 | 0.3 | 0.2 | n.d. |
| Phenols (% w/w) d | n.d. | n.d. | n.d. | 0.4 | n.d. | n.d. | 3.0 | 2.1 | n.d. | 0.6 |

2.2.5. Linkage Analysis of the Polysaccharide Fractions
| 100WTRBH-I-I | 100WTSBH-I-I | 100WTLH-I-I | ||
|---|---|---|---|---|
| Ara | Tf | n.d. | 10.2 | 14.6 |
| 1,2f | n.d. | 0.1 | 1.2 | |
| 1,3f | n.d. | 0.2 | 0.7 | |
| 1,5f | n.d. | 4.3 | 6.7 | |
| 1,3,5f | n.d. | 1.6 | 2.1 | |
| Rha | Tp | n.d. | 1.5 | 1.3 |
| 1,3p | 1.3 | 0.1 | 0.1 | |
| 1,2p | 21.3 | 0.5 | n.d. | |
| 1,2,4p | 1.0 | 2.2 | 1.7 | |
| Xyl | Tp | n.d. | n.d. | Trace |
| 1,4p | n.d. | 1.5 | 23.8 | |
| Gal | Tp | 1.2 | 3.4 | 2.1 |
| 1,4p | n.d. | 0.5 | 3.3 | |
| 1,3p | 2.1 | 1.2 | 4.8 | |
| 1,6p | n.d. | 9.9 | 2.1 | |
| 1,3,6p | 0.4 | 3.1 | 10.7 | |
| 1,3,4,6p | n.d. | 0.6 | 0.3 | |
| Glc | 1,3p | n.d. | 0.7 | 2.3 |
| 1,4p | 21.8 | 13.9 | 3.5 | |
| 1,6p | 4.0 | 3.8 | 0.7 | |
| 1,4,6p | Trace | 0.4 | 0.5 | |
| GlcA | Tp | n.d. | 1.0 | 0.3 |
| 1,4p | n.d. | 0.7 | n.d. | |
| GalA | Tp | 1.3 | n.d. | n.d. |
| 1,4p | 44.6 | 34.6 | 12.3 | |
| 1,3,4p | n.d. | 2.3 | 0.1 |
2.3. Discussion
3. Experimental
3.1. Plant Material
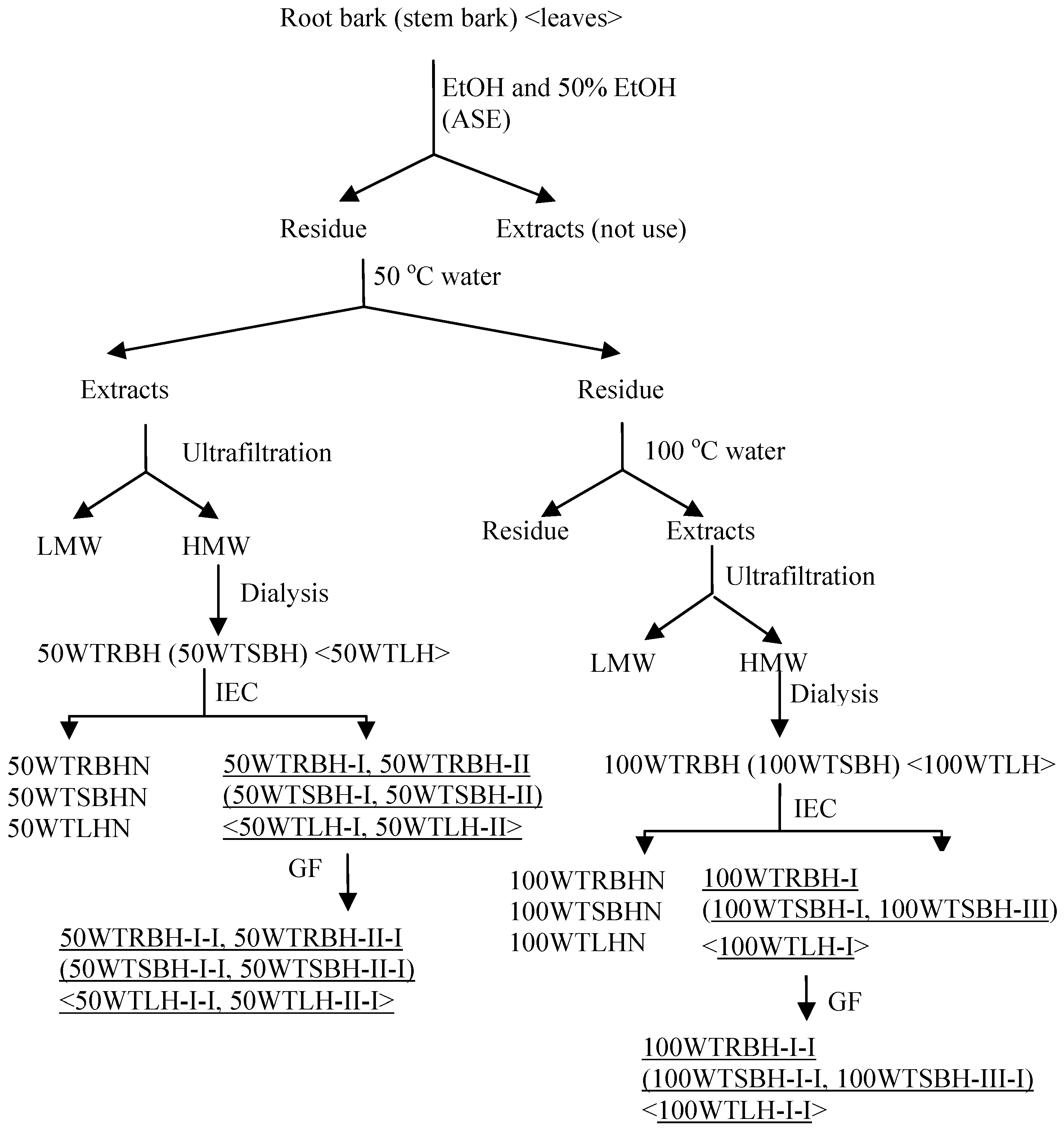
3.2. Extraction of Polysaccharides
3.3. Fractionation and Characterization of Polysaccharides

3.3.1. Ion Exchange Chromatography and Gel Filtration
3.3.2. Determination of Monosaccharide Composition
3.3.3. Test for the Presence of Starch
3.3.4. Molecular Weight Determination
3.3.5. Precipitation with the Yariv β-Glucosyl Reagent
3.3.6. Determination of Phenolic Content
3.3.7. Determination of Protein Content
3.3.8. Complement Fixation Assay
3.3.9. Determination of Glycosidic Linkages
4. Conclusions
Acknowledgments
Abbreviations
| AG-I | arabinogalactan type I |
| AG-II | arabinogalactan type II |
| Ara | arabinose |
| ASE | accelerated solvent extraction |
| DCM | dichloromethane |
| EtOH | ethanol |
| Gal | galactose |
| GalA | galacturonic acid |
| Glc | glucose |
| GlcA | glucuronic acid |
| GF | gel filtration |
| HMW | high molecular weight |
| IEC | ion exchange chromatography |
| LMW | low molecular weight |
| Man | mannose |
| MeOH | methanol |
| RG-I | rhamnogalacturonan type I |
| Rha | rhamnose |
| Xyl | xylose |
Author Contributions
Conflicts of Interest
References
- Pham, A.T.; Dvergsnes, C.; Togola, A.; Wangensteen, H.; Diallo, D.; Paulsen, B.S.; Malterud, K.E. Terminalia macroptera, its current medicinal use and future perspectives. J. Ethnopharmacol. 2011, 137, 1486–1491. [Google Scholar] [CrossRef]
- Silva, O.; Viegas, S.; de Mello-Sampayo, C.; Costa, M.J.; Serrano, R.; Cabrita, J.; Gomes, E.T. Anti-Helicobacter pylori activity of Terminalia macroptera root. Fitoterapia 2012, 83, 872–876. [Google Scholar] [CrossRef]
- Silva, O.; Gomes, E.T.; Wolfender, J.L.; Marston, A.; Hostettmann, K. Application of high-performance liquid chromatography coupled with ultraviolet spectroscopy and electrospray mass spectrometry to the characterisation of ellagitannins from Terminalia macroptera roots. Pharmaceut. Res. 2000, 17, 1396–1401. [Google Scholar] [CrossRef]
- Silva, O.; Duarte, A.; Cabrita, J.; Pimentel, M.; Gomes, E.T. Antimicrobial activity of Terminalia macroptera root. J. Ethnopharmacol. 1997, 57, 203–207. [Google Scholar] [CrossRef]
- Silva, O.; Duarte, A.; Cabrita, J.; Pimentel, M.; Diniz, A.; Gomes, E. Antimicrobial activity of Guinea-Bissau traditional remedies. J. Ethnopharmacol. 1996, 50, 55–59. [Google Scholar] [CrossRef]
- Nongonierma, R.; Proliac, A.; Raynaud, J. O-glycosyl flavonoids from the flowers of Terminalia macroptera Guill and Perr (Combretaceae). Pharm. Acta Helv. 1990, 65, 233–235. [Google Scholar]
- Nongonierma, R.; Proliac, A.; Raynaud, J. Vitexin and isovitexin in Terminalia macroptera Guill et Perr flowers (Combretaceae). Pharmazie 1988, 43, 293–300. [Google Scholar]
- Nongonierma, R.; Proliac, A.; Raynaud, J. 2 Mono-C-glycosyl flavonoids from flowers of Terminalia macroptera Guill and Perr (Combretaceae). Pharmazie 1987, 42, 871–872. [Google Scholar]
- Conrad, J.; Vogler, B.; Klaiber, I.; Reeb, S.; Guse, J.H.; Roos, G.; Kraus, W. Vanillic acid 4-O-β-d-(6'-O-galloyl) glucopyranoside and other constituents from the bark of Terminalia macroptera Guill. et Perr. Nat. Prod. Lett. 2001, 15, 35–42. [Google Scholar] [CrossRef]
- Conrad, J.; Vogler, B.; Klaiber, I.; Roos, G.; Walter, U.; Kraus, W. Two triterpene esters from Terminalia macroptera bark. Phytochemistry 1998, 48, 647–650. [Google Scholar] [CrossRef]
- Pham, A.T.; Malterud, K.E.; Paulsen, B.S.; Diallo, D.; Wangensteen, H. DPPH radical scavenging and xanthine oxidase inhibitory activity of Terminalia macroptera leaves. Nat. Prod. Commun. 2011, 6, 1125–1128. [Google Scholar]
- Conrad, J.; Vogler, B.; Reeb, S.; Klaiber, I.; Papajewski, S.; Roos, G.; Vasquez, E.; Setzer, M.C.; Kraus, W. Isoterchebulin and 4,6-O-terchebuloyl-d-glucose, novel hydrolysable tannins from Terminalia macroptera. J. Nat. Prod. 2001, 64, 294–299. [Google Scholar] [CrossRef]
- Adesina, S.K.; Idowu, O.; Ogundaini, A.O.; Oladimeji, H.; Olugbade, T.A.; Onawunmi, G.O.; Pais, M. Antimicrobial constituents of the leaves of EAcalypha wilkesiana and Acalypha hispida. Phytother. Res. 2000, 14, 371–374. [Google Scholar] [CrossRef]
- Paulsen, B.S.; Barsett, H. Bioactive pectic polysaccharides. Adv. Polym. Sci. 2005, 186, 69–101. [Google Scholar] [CrossRef]
- Austarheim, I.; Mahamane, H.; Sanogo, R.; Togola, A.; Khaledabadi, M.; Vestrheim, A.C.; Inngjerdingen, K.T.; Michaelsen, T.E.; Diallo, D.; Paulsen, B.S. Anti-ulcer polysaccharides from Cola cordifolia bark and leaves. J. Ethnopharmacol. 2012, 143, 221–227. [Google Scholar] [CrossRef]
- Lin, M.; Xia, B.; Yang, M.; Gao, S.; Hou, Y.; Lou, G. Anti-ovarian cancer potential of two acidic polysaccharides from the rhizoma of Menispermum dauricum. Carbohydr. Polym. 2013, 92, 2212–2217. [Google Scholar] [CrossRef]
- Samuelsen, A.B.; Paulsen, B.S.; Wold, J.K.; Otsuka, H.; Kiyohara, H.; Yamada, H.; Knutsen, S.H. Characterization of a biologically active pectin from Plantago. Major L. Carbohydr. Polym. 1996, 30, 37–44. [Google Scholar] [CrossRef]
- Yamada, H.; Kiyohara, H. Complement-Activating Polysaccharides from Medicinal Herbs. In Immunomodulatory Agents from Plants; Wanger, H., Ed.; Birkhäuser Berlag: Basel, Switzerland, 1999; pp. 161–202. [Google Scholar]
- Zong, A.; Cao, H.; Wang, F. Anticancer polysaccharides from natural resources: A review of recent research. Carbohyd. Polym. 2012, 90, 1395–1410. [Google Scholar] [CrossRef]
- Ezzell, J.L.; Richter, B.E.; Felix, W.D.; Black, S.R.; Meikle, J.E. A comparison of accelerated solvent extraction with conventional solvent extraction for organophosphorus pesticides and herbicides. LC GC 1995, 13, 390–398. [Google Scholar]
- Richter, B.E.; Ezzell, J.L.; Felix, D.; Roberts, K.A.; Later, D.W. An accelerated solvent-extraction system for the rapid preparation of environmental organic-compounds in soil. Am. Lab. 1995, 27, 24–28. [Google Scholar]
- Richter, B.E.; Raynie, D. Accelerated Solvent Extraction (ASE) and High-Temperature Water Extraction. In Comprehensive Sampling and Sample Preparation: Analytical Techniques for Scientists; Pawliszyn, J., Ed.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 2; pp. 105–115. [Google Scholar]
- Wang, P.; Zhang, Q.; Wang, Y.; Wang, T.; Li, X.; Ding, L.; Jiang, G. Evaluation of Soxhlet extraction, accelerated solvent extraction and microwave-assisted extraction for the determination of polychlorinated biphenyls and polybrominated diphenyl ethers in soil and fish samples. Anal. Chim. Acta 2010, 663, 43–48. [Google Scholar] [CrossRef]
- Olivella, M.À. Isolation and analysis of polycyclic aromatic hydrocarbons from natural water using accelerated solvent extraction followed by gas chromatography–mass spectrometry. Talanta 2006, 69, 267–275. [Google Scholar]
- Sporring, S.; Bøwadt, S.; Svensmark, B.; Björklund, E. Comprehensive comparison of classic Soxhlet extraction with Soxtec extraction, ultrasonication extraction, supercritical fluid extraction, microwave assisted extraction and accelerated solvent extraction for the determination of polychlorinated biphenyls in soil. J. Chromatogr. A 2005, 1090, 1–9. [Google Scholar] [CrossRef]
- Wang, W.; Meng, B.; Lu, X.; Liu, Y.; Tao, S. Extraction of polycyclic aromatic hydrocarbons and organochlorine pesticides from soils: A comparison between Soxhlet extraction, microwave-assisted extraction and accelerated solvent extraction techniques. Anal. Chim. Acta 2007, 602, 211–222. [Google Scholar] [CrossRef]
- Xu, X.; Li, Q.; Yuan, J.; Wang, S.; Wang, W.; Lee, F.; Wang, X. Determination of three kinds of chloroacetanilide herbicides in Radix Pseudostellariae by accelerated solvent extraction and gas chromatography-mass Spectrometry. Chin. J. Anal. Chem. 2007, 35, 206–210. [Google Scholar] [CrossRef]
- Schäfer, K. Accelerated solvent extraction of lipids for determining the fatty acid composition of biological material. Anal. Chim. Acta 1998, 358, 69–77. [Google Scholar] [CrossRef]
- Chen, J.; Li, W.; Yang, B.; Guo, X.; Lee, F.; Wang, X. Determination of four major saponins in the seeds of Aesculus chinensis Bunge using accelerated solvent extraction followed by high-performance liquid chromatography and electrospray-time of flight mass spectrometry. Anal. Chim. Acta 2007, 596, 273–280. [Google Scholar] [CrossRef]
- Chitescu, C.L.; Oosterink, E.; de Jong, J.; Stolker, A.A. Ultrasonic or accelerated solvent extraction followed by U-HPLC-high mass accuracy MS for screening of pharmaceuticals and fungicides in soil and plant samples. Talanta 2012, 88, 653–662. [Google Scholar] [CrossRef]
- Hossain, M.B.; Barry-Ryan, C.; Martin-Diana, A.B.; Brunton, N.P. Optimisation of accelerated solvent extraction of antioxidant compounds from rosemary (Rosmarinus officinalis L.), marjoram (Origanum majorana L.) and oregano (Origanum vulgare L.) using response surface methodology. Food Chem. 2011, 126, 339–346. [Google Scholar] [CrossRef]
- Pearson, C.H.; Cornish, K.; Rath, D.J. Extraction of natural rubber and resin from guayule using an accelerated solvent extractor. Ind. Crop. Prod. 2013, 43, 506–510. [Google Scholar]
- Zhang, Y.; Liu, C.; Qi, Y.; Li, S.; Wang, J. Application of accelerated solvent extraction coupled with counter-current chromatography to extraction and online isolation of saponins with a broad range of polarity from Panax notoginseng. Sep. Purif. Technol. 2013, 106, 82–89. [Google Scholar] [CrossRef]
- Morgan, J.N.; Rosenblum, L.; Garris, S. Comparison of five extraction methods for determining incurred and fortified pesticides in dietary composites. J. AOAC Int. 2002, 85, 1167–1176. [Google Scholar]
- Pecorelli, I.; Galarini, R.; Bibi, R.; Floridi, A.; Casciarri, E.; Floridi, A. Simultaneous determination of 13 quinolones from feeds using accelerated solvent extraction and liquid chromatography. Anal. Chim. Acta 2003, 483, 81–89. [Google Scholar] [CrossRef]
- Sun, H.; Ge, X.; Lv, Y.; Wang, A. Application of accelerated solvent extraction in the analysis of organic contaminants, bioactive and nutritional compounds in food and feed. J. Chromatogr. A 2012, 1237, 1–23. [Google Scholar] [CrossRef]
- Breithaupt, D.E. Simultaneous HPLC determination of carotenoids used as food coloring additives: Applicability of accelerated solvent extraction. Food Chem. 2004, 86, 449–456. [Google Scholar] [CrossRef]
- Klejdus, B.; Mikelová, R.; Adam, V.; Zehnálek, J.; Vacek, J.; Kizek, R.; Kubáň, V. Liquid chromatographic–mass spectrometric determination of genistin and daidzin in soybean food samples after accelerated solvent extraction with modified content of extraction cell. Anal. Chim. Acta 2004, 517, 1–11. [Google Scholar] [CrossRef]
- Le Normand, M.; Edlund, U.; Holmbom, B.; Monica, E.K. Hot-water extraction and characterization of spruce bark non-cellulosic polysaccharides. Nord. Pulp Pap. Res. J. 2012, 27, 18–23. [Google Scholar]
- Le Normand, M.; Mélida, H.; Holmbom, B.; Michaelsen, T.E.; Inngjerdingen, M.; Bulone, V.; Paulsen, B.S.; Ek, M. Hot-water extracts from the inner bark of Norway spruce with immunomodulating activities. Carbohydr. Polym. 2014, 101, 699–704. [Google Scholar] [CrossRef]
- Song, T.; Pranovich, A.; Holmbom, B. Extraction of galactoglucomannan from spruce wood with pressurized hot water. Holzforschung 2008, 62, 659–666. [Google Scholar]
- Michaelsen, T.E.; Gilje, A.; Samuelsen, A.B.; Hagasen, K.; Paulsen, B.S. Interaction between human complement and a pectin type polysaccharide fraction, PMII, from the leaves of Plantago major L. Scand. J. Immunol. 2000, 52, 483–490. [Google Scholar] [CrossRef]
- Inngjerdingen, K.T.; Ballo, N.; Zhang, B.; Malterud, K.E.; Michaelsen, T.E.; Diallo, D.; Paulsen, B.S. A comparison of bioactive aqueous extracts and polysaccharide fractions from roots of wild and cultivated Cochlospermum tinctorium A. rich. Phytochemistry 2013, 93, 136–143. [Google Scholar] [CrossRef]
- Paulsen, B.S.; Craik, D.J.; Dunstan, D.E.; Stone, B.A.; Bacic, A. The Yariv reagent: Behavior in different solvents and interaction with a gum arabic Arabinogalactanprotein. Carbohydr. Polym. 2014, in press. [Google Scholar]
- Vincken, J.P.; Schols, H.A.; Oomen, R.J.F.J.; McCann, M.C.; Ulvskov, P.; Voragen, A.G.J.; Visser, R.G.F. If homogalacturonan were a side chain of Rhamnogalacturonan I. implications for cell war architecture. Plant. Physiol. 2003, 132, 1781–1789. [Google Scholar]
- Capek, P.; Rosik, J.; Kardosova, A.; Toman, R. Polysaccharides from the roots of the Marsh mallow (Althaea. officinalis L., var. Rhobusta): Structural features of an acidic polysaccharide. Carbohydr. Res. 1987, 164, 443–452. [Google Scholar] [CrossRef]
- Renard, C.M.G.C.; Crepeau, M.J.; Thibault, J.F. Glucuronic acid directly linked to galacturonic acid in the rhamnogalacturonan backbone of beet pectins. Eur. J. Biochem. 1999, 266, 566–574. [Google Scholar] [CrossRef]
- Paulsen, B.S. Plant polysaccharides with immunostimulatory activities. Curr. Org. Chem. 2001, 5, 939–950. [Google Scholar]
- Grønhaug, T.E.; Ghildyal, P.; Barsett, H.; Michaelsen, T.E.; Morris, G.; Diallo, D.; Inngjerdingen, M.; Paulsen, B.S. Bioactive arabinogalactans from the leaves of Opilia. celtidifolia Endl. ex Walp. (Opiliaceae). Glycobiology 2010, 20, 1654–1664. [Google Scholar] [CrossRef]
- Nergård, C.S.; Matsumoto, T.; Inngjerdingen, M.; Inngjerdingen, K.; Hokputsa, S.; Harding, S.E.; Michaelsen, T.E.; Diallo, D.; Kiyohara, H.; Paulsen, B.S.; et al. Structural and immunological studies of a pectin and a pectic arabinogalactan from Vernonia kotschyana Sch. Bip. ex Walp. (Asteraceae). Carbohydr. Res. 2005, 340, 115–130. [Google Scholar] [CrossRef]
- Togola, A.; Inngjerdingen, M.; Diallo, D.; Barsett, H.; Rolstad, B.; Michaelsen, T.E.; Paulsen, B.S. Polysaccharides with complement fixing and macrophage stimulation activity from Opilia celtidifolia, isolation and partial characterization. J. Ethnopharmacol. 2008, 115, 423–431. [Google Scholar] [CrossRef]
- Waldron, K.W.; Faulds, C.B. Cell Wall Polysaccharides: Composition and Structure. In Comprehensive Glycoscience.- From Chemistry to Stystems Biology; Kamerling, J.P., Boons, G.J., Lee, Y.C., Suzuki, A., Taniguchi, N., Voragen, A.G.J., Eds.; Elsevier: Oxford, UK, 2007; Volume 1; pp. 181–201. [Google Scholar]
- Yamada, H.; Kiyohara, H. Immunomodulating Activity of Plant Polysaccharide Structures. In Comprehensive Glycoscience - From Chemistry to Stystems Biology; Kamerling, J.P., Boons, G.J., Lee, Y.C., Suzuki, A., Taniguchi, N., Voragen, A.G.J., Eds.; Elsevier: Oxford, UK, 2007; pp. 663–694. [Google Scholar]
- Inngjerdingen, K.T.; Coulibaly, A.; Diallo, D.; Michaelsen, T.E.; Paulsen, B.S. A complement fixing polysaccharide from Biophytum. petersianum Klotzsch, a medicinal plant from Mali, West Africa. Biomacromolecules 2006, 7, 48–53. [Google Scholar] [CrossRef]
- Inngjerdingen, K.T.; Meskini, S.; Austarheim, I.; Ballo, N.; Inngjerdingen, M.; Michaelsen, T.E.; Diallo, D.; Paulsen, B.S. Chemical and biological characterization of polysaccharides from wild and cultivated roots of Vernonia kotschyana. J. Ethnopharmacol. 2012, 139, 350–358. [Google Scholar] [CrossRef]
- Samuelsen, A.B.; Westereng, B.; Yousif, O.; Holtekjølen, A.K.; Michaelsen, T.E.; Knutsen, S.H. Structural features and complement-fixing activity of pectin from three Brassica oleracea varieties: White cabbage, kale, and red kale. Biomacromolecules 2007, 8, 644–649. [Google Scholar] [CrossRef]
- Pieters, L.A.C.; de Bruyne, T.E.; Vlietinck, A.J. Low molecular weight compounds with complement activity. In Immunomodulatory Agents from Plants, Progress in Inflammation Research; Wagner, H., Ed.; Birkhäuser Basel: Basel, Switzerland, 1999; pp. 137–160. [Google Scholar]
- Levigne, S.V.; Ralet, M.-C.J.; Quemener, B.C.; Pollet, B.N.-L.; Lapierre, C.; Thibault, J.-F.J. Isolation from Sugar Beet cell walls of arabinan oligosaccharides esterified by two ferulic acid monomers. Plant Physiol. 2004, 134, 1173–1180. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Austarheim, I.; Christensen, B.E.; Hegna, I.K.; Petersen, B.O.; Duus, J.O.; Bye, R.; Michaelsen, T.E.; Diallo, D.; Inngjerdingen, M.; Paulsen, B.S. Chemical and biological characterization of pectin-like polysaccharides from the bark of the Malian medicinal tree Cola cordifolia. Carbohydr. Polym. 2012, 89, 259–268. [Google Scholar] [CrossRef]
- Barsett, H.; Paulsen, B.S.; Habte, Y. Further characterization of polysaccharides in seeds from Ulmus glabra Huds. Carbohydr. Polym. 1992, 18, 125–130. [Google Scholar] [CrossRef]
- Chambers, R.E.; Clamp, J.R. Assessment of methanolysis and other factors used in the analysis of carbohydrate-containing materials. Biochem. J. 1971, 125, 1009–1018. [Google Scholar]
- Hunter, R.A.; McIntyre, B.L.; Mcilroy, R.J. Water-soluble carbohydrates of tropical pasture grasses and legumes. J. Sci. Food Agric. 1970, 21, 400–405. [Google Scholar] [CrossRef]
- Van Holst, G.J.; Clarke, A.E. Quantification of arabinogalactan-protein in plant extracts by single radial gel diffusion. Anal. Biochem. 1985, 148, 446–450. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticult. 1965, 37, 144–158. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dunkelberger, J.R.; Song, W.C. Complement and its role in innate and adaptive immune responses. Cell Res. 2010, 20, 34–50. [Google Scholar]
- Grønhaug, T.E.; Kiyohara, H.; Sveaass, A.M.; Diallo, D.; Yamada, H.; Paulsen, B.S. Beta-d-(1°4)-galactan-containing side chains in RG-I regions of pectic polysaccharides from Biophytum petersianum Klotzsch contribute to expression of immunomodulating activity against intestinal Peyer’s patch cells and macrophages. Phytochemistry 2011, 72, 2139–2147. [Google Scholar] [CrossRef]
- Kim, J.B.; Carpita, N.C. Changes in esterification of the uronic-acid groups of cell-wall polysaccharides during elongation of maize coleoptiles. Plant. Physiol. 1992, 98, 646–653. [Google Scholar] [CrossRef]
- Sweet, D.P.; Shapir, R.H.; Albersheim, P. Quantitative analysis by various GLC response factor theories for partially methylated and partially ethylated alditol acetates. Carbohydr. Res. 1975, 40, 217–225. [Google Scholar] [CrossRef]
- Sample Availability: Samples of polysaccharide fractions are available from the authors.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zou, Y.-F.; Zhang, B.-Z.; Barsett, H.; Inngjerdingen, K.T.; Diallo, D.; Michaelsen, T.E.; Paulsen, B.S. Complement Fixing Polysaccharides from Terminalia macroptera Root Bark, Stem Bark and Leaves. Molecules 2014, 19, 7440-7458. https://doi.org/10.3390/molecules19067440
Zou Y-F, Zhang B-Z, Barsett H, Inngjerdingen KT, Diallo D, Michaelsen TE, Paulsen BS. Complement Fixing Polysaccharides from Terminalia macroptera Root Bark, Stem Bark and Leaves. Molecules. 2014; 19(6):7440-7458. https://doi.org/10.3390/molecules19067440
Chicago/Turabian StyleZou, Yuan-Feng, Bing-Zhao Zhang, Hilde Barsett, Kari Tvete Inngjerdingen, Drissa Diallo, Terje Einar Michaelsen, and Berit Smestad Paulsen. 2014. "Complement Fixing Polysaccharides from Terminalia macroptera Root Bark, Stem Bark and Leaves" Molecules 19, no. 6: 7440-7458. https://doi.org/10.3390/molecules19067440