Standardized Salvia miltiorrhiza Extract Suppresses Hepatic Stellate Cell Activation and Attenuates Steatohepatitis Induced by a Methionine-Choline Deficient Diet in Mice

Abstract
:1. Introduction

2. Results and Discussion
2.1. Body Weight and Clinical Observations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight (g) | Liver/Body Weight (%) | |
|---|---|---|
| 8 weeks | ||
| Normal diet | 27.03 ± 0.5155 | |
| MCD diet | 16.76 ± 0.2428 | |
| 12 weeks | ||
| Normal control | 26.76 ± 0.7128 | 3.144 ± 0.06255 |
| MCD | 17.01 ± 0.7240 | 4.405 ± 0.1035 |
| MCD+Low | 16.72 ± 0.6787 | 3.924 ± 0.1320 * |
| MCD+Mid | 16.63 ± 0.6992 | 4.277 ± 0.1320 |
| MCD+High | 16.31 ± 0.6780 | 3.814 ± 0.1272 ** |
| 14 weeks | ||
| Normal control | 27.30 ± 0.7598 | 3.282 ± 0.09449 |
| MCD | 16.95 ± 0.7444 | 3.751 ± 0.2258 |
| MCD+Low | 17.16 ± 0.6988 | 4.035 ± 0.06351 |
| MCD+Mid | 16.75 ± 0.7078 | 4.303 ± 0.3289 |
| MCD+High | 16.54 ± 0.6764 | 3.928 ± 0.2047 |
2.2. Liver Weight
2.3. Mortality
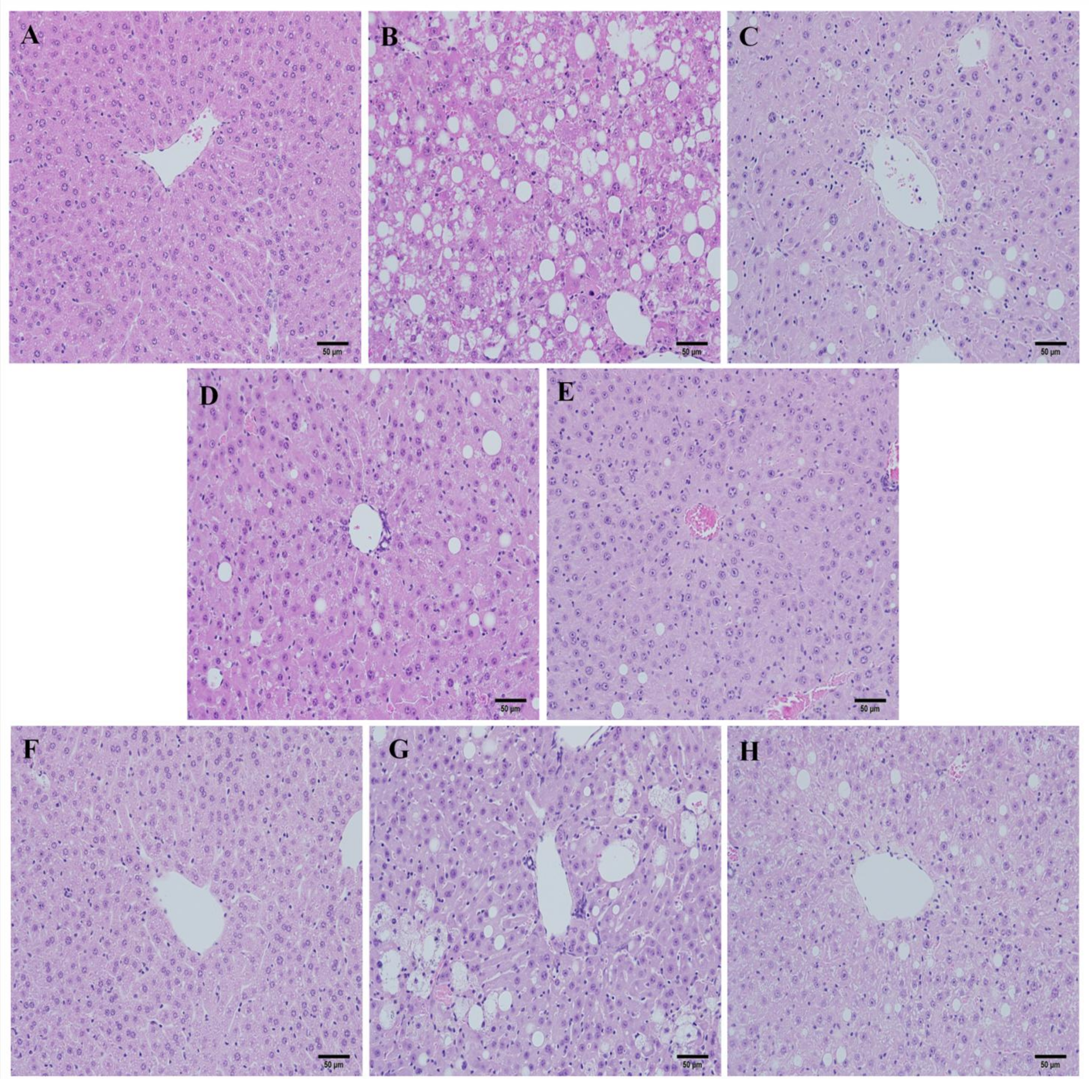
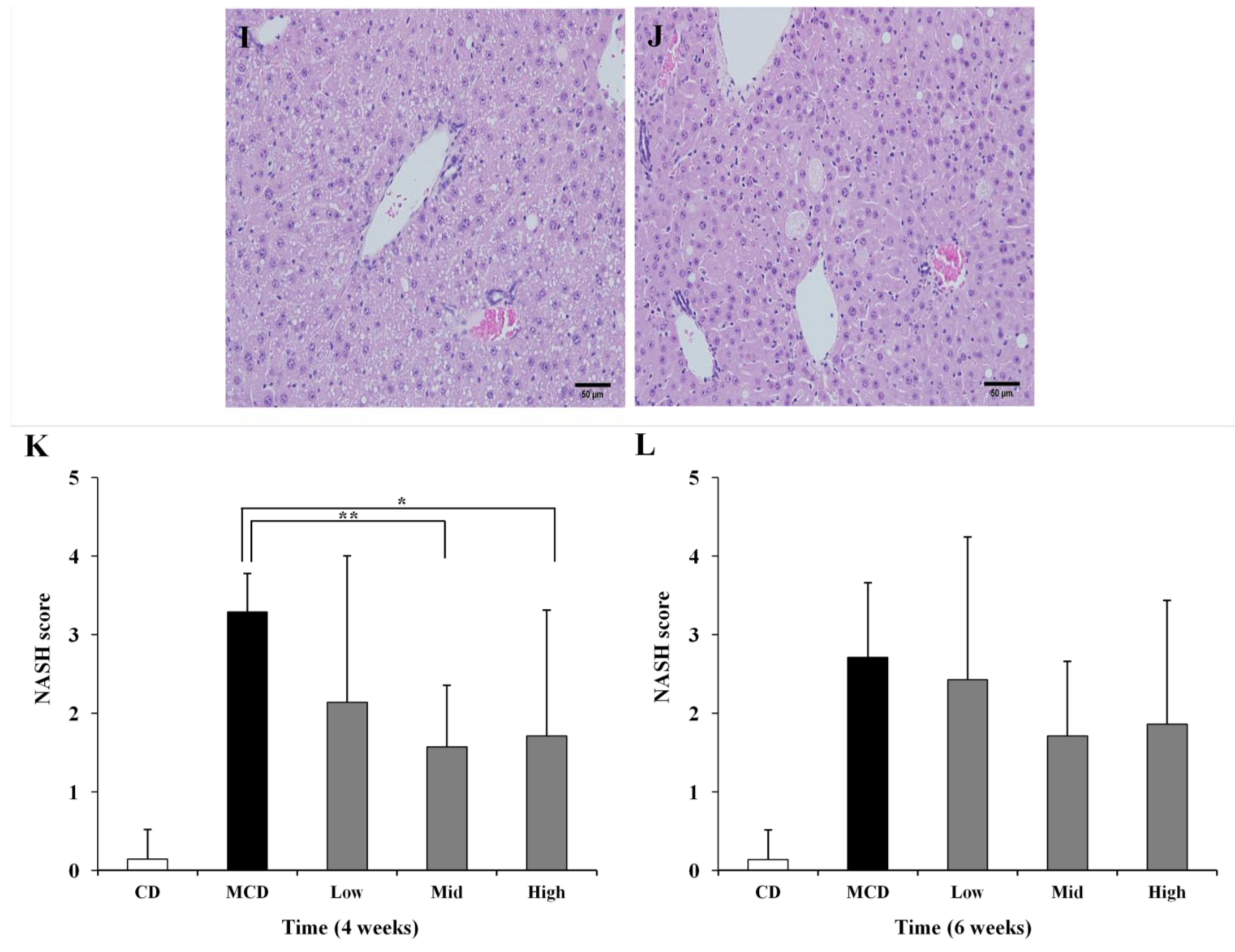
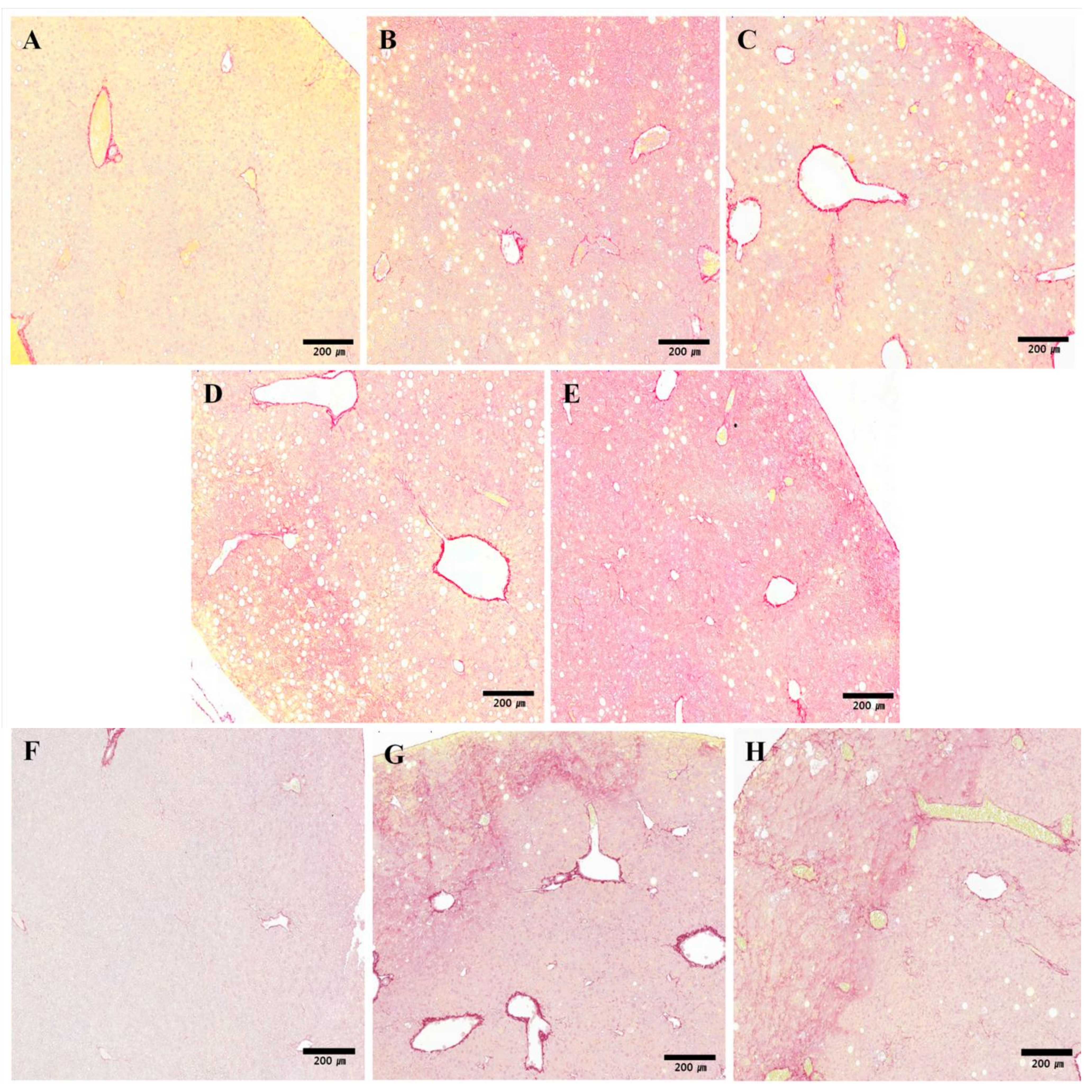
2.4. Histopathology Analysis




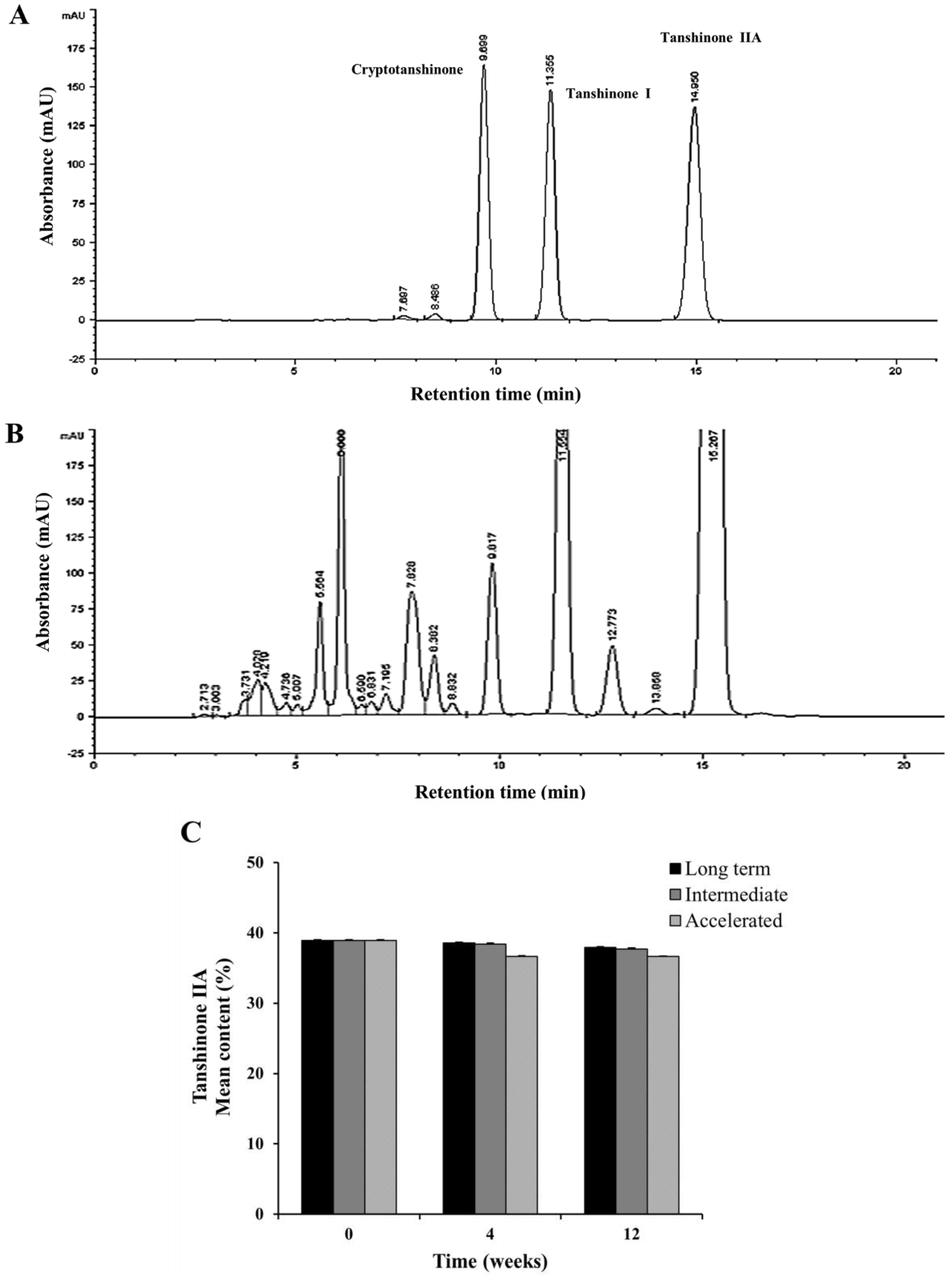
2.5. Stability Test of Tanshinone IIA

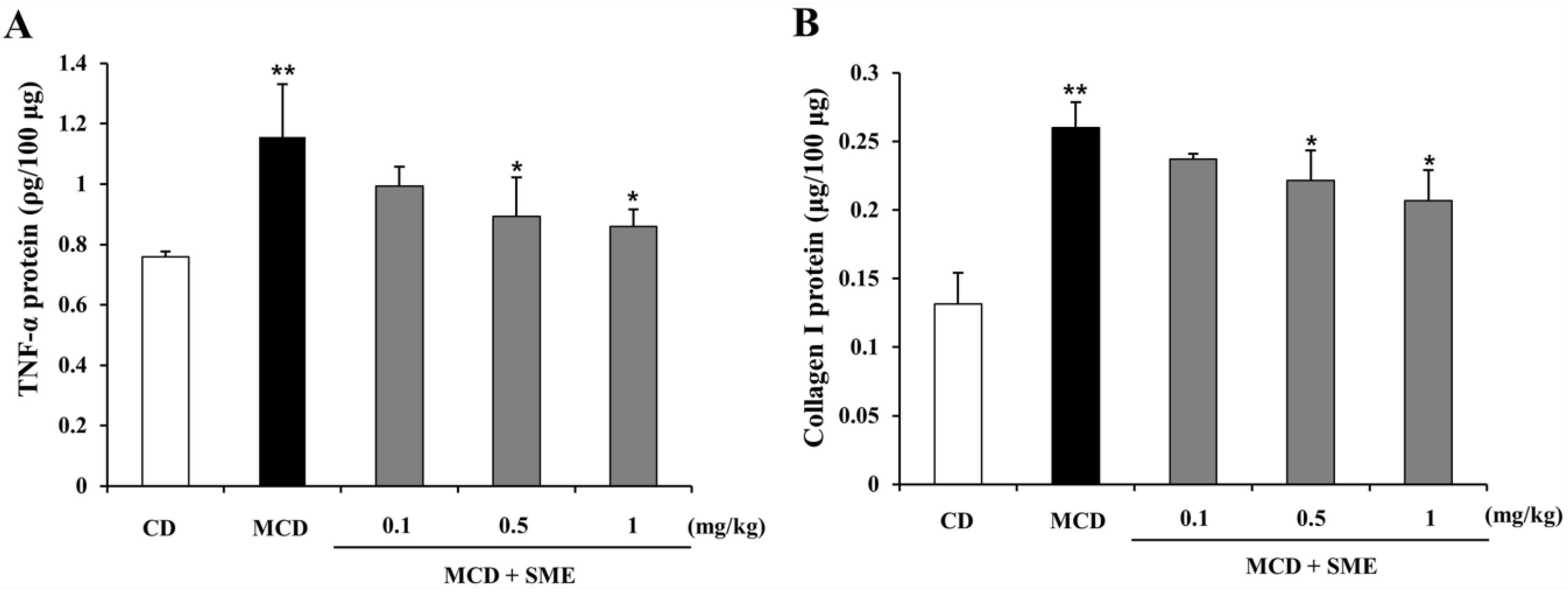
2.6. Effect of SME on Hepatic TNF-α and Collagen I Expression in NASH Induced by the MCD Diet in Mice

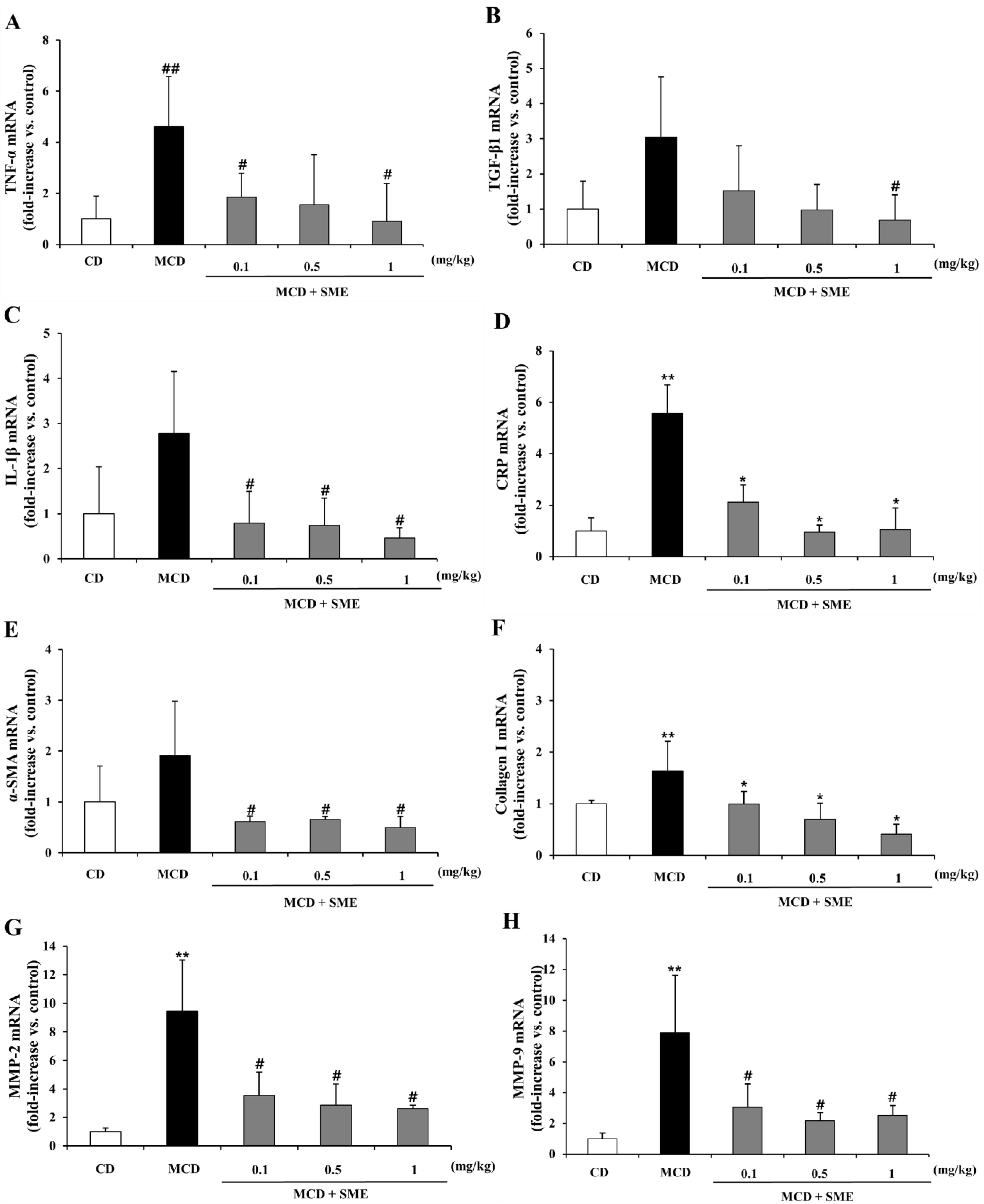
2.7. Effect of SME on Hepatic mRNA Expression of NASH-Related Specific Genes in the MCD Dietary Mouse Model

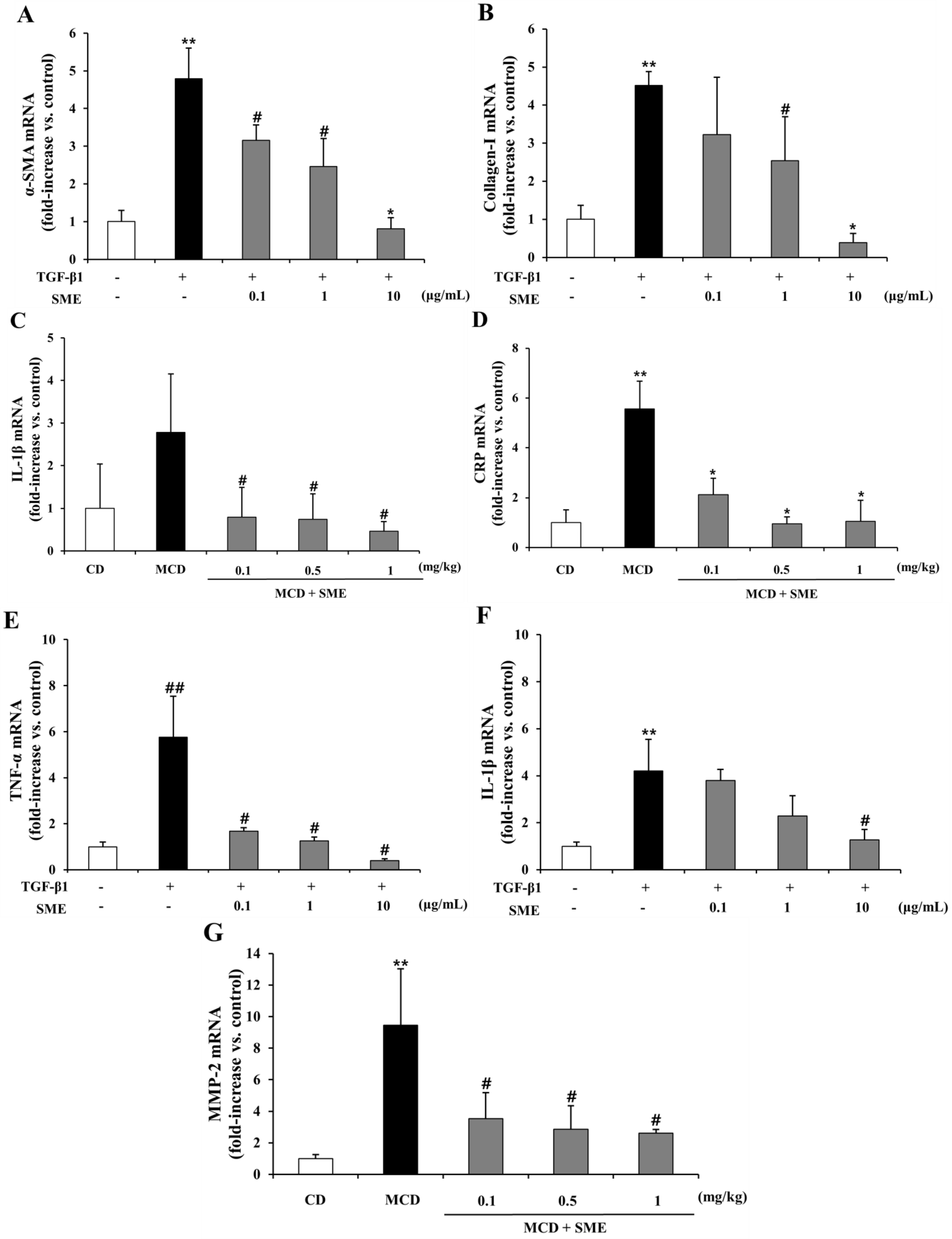
2.8. Effect of SME on mRNA Expression of NASH-Related Specific Genes in HSCs Induced by TGF-β1

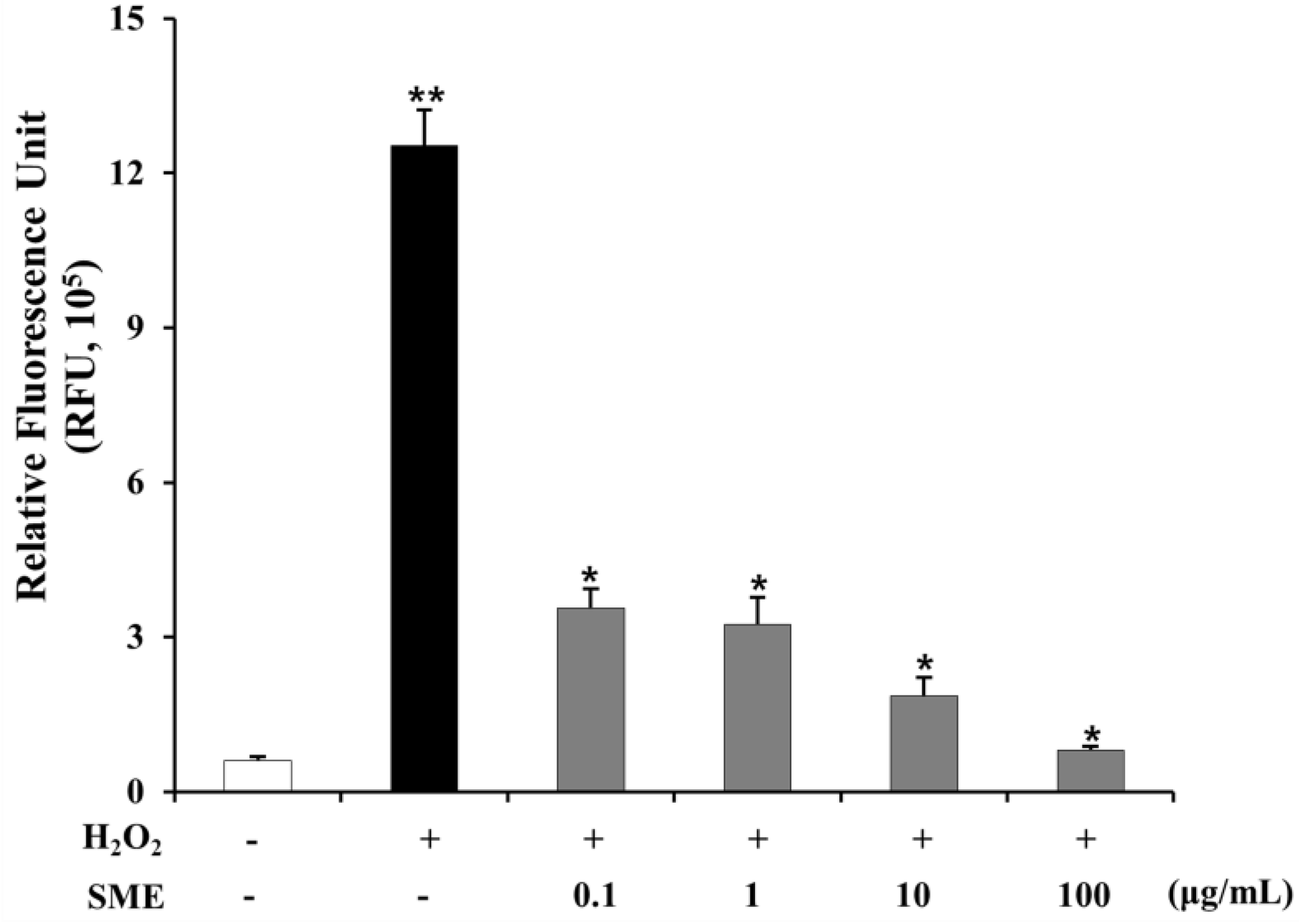
2.9. Effect of SME on HSCs Induced by Oxidative Stress

3. Experimental
3.1. Preparation of Standardized Extract of Salvia Miltiorrhiza
3.2. Stability Analysis of Tanshinone IIA by HPLC
3.3. Animals and Experimental Protocol
3.4. Liver Histopathology
| Diet | Test Article | Group | Test Article Administration Period (weeks) | Dose Level | Number of Animals (male) |
|---|---|---|---|---|---|
| (mg/kg) | |||||
| Normal Diet | Normal | 1. Normal | 4 | 0 | 7 |
| 2. Normal | 6 | 7 | |||
| MCD Diet | Control | 3. Control | 4 | 0 | 7 |
| 4. Control | 6 | 7 | |||
| Low dose | 5. MCD+Low dose | 4 | 0.1 | 7 | |
| 6. MCD+Low dose | 6 | 7 | |||
| Mid dose | 7. MCD+Mid dose | 4 | 0.5 | 7 | |
| 8. MCD+Mid dose | 6 | 7 | |||
| High Dose | 9. MCD+High dose | 4 | 1 | 7 | |
| 10. MCD+High dose | 6 | 7 |
3.4.1. NASH (NAFLD Activity Score) Evaluation
3.4.2. Fibrosis Evaluation and Measurement of Collagen Distribution
3.5. Cell Culture & Reagents
3.6. Preparation of Liver Lysate
3.7. RNA Isolation and Quantitative PCR
| Target Gene | Primer (Forward/Reverse) | Reference | ||
|---|---|---|---|---|
| Human | ||||
| α-SMA | F: ctg ttc cag cca tcc ttc at | R: tca tga tgc tgt tgt agg tgg t | ENSG00000107796 | |
| MMP-2 | F: ccc caa aac gga caa aga g | R: ctt cag cac aaa acg gtt gc | ENSG00000087245 | |
| MMP-9 | F: tct tcc ctg gag acc tga ga | R: gag tgt aac cat agc ggt aca gg | ENSG00000100985 | |
| Collagen I | F: ggg att ccc tgg acc taa ag | R: gga aca cct cgc tct cca | ENSG00000108821 | |
| TNF-α | F: cag cct ctt ctc ctt cct gat | R: gcc aga ggg ctg att aga ga | ENSG00000232810 | |
| CRP | F: cca gct gtg ggt cct gaa | R: cac agc ccc aca agg ttc | ENSG00000132693 | |
| IL-1β | F: agc tga tgg ccc taa aca ga | R: gtc gga gat tcg tag ctg ga | ENSG00000125538 | |
| Mouse | ||||
| α-SMA | F: ctc tct tcc agc cat ctt tca t | R: tat agg tgg ttt cgt gga tgc | ENSMUSG00000035783 | |
| MMP-2 | F: aac ttt gag aag gat ggc aag t | R: tgc cac cca tgg taa aca a | ENSMUSG00000031740 | |
| MMP-9 | F: acg aca tag acg gca tcc a | R: gct gtg gtt cag ttg tgg tg | ENSMUSG00000017737 | |
| Collagen I | F: ctg gtc ctg ctg gct ttg | R: acc acg atc gcc att ctt | ENSMUSG00000001506 | |
| TNF-α | F: cca gac cct cac act caca a | R: ttg aga tcc atg ccg ttg | ENSMUSG00000024401 | |
| CRP | F: ctc gga ctt ttg gtc atg aag | R: aaa ggt gtt cag tgg ctt ctt t | ENSMUSG00000037942 | |
| IL-1β | F: agt tga cgg acc cca aaa g | R: ttt gaa gct gga tgc tct cat | ENSMUSG00000027398 | |
| TGF-β1 | F: tgg agc aac atg tgg aac tc | R: gtc agc agc cgg tta cca | ENSMUSG00000002603 | |
3.8. Enzyme-Linked Immunosorbent Assay (ELISA)
3.9. Intracellular ROS Activity
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- De Alwis, N.M.; Day, C.P. Non-alcoholic fatty liver disease: The mist gradually clears. J. Hepatol. 2008, 48, S104–S112. [Google Scholar] [CrossRef]
- Marchesini, G.; Brizi, M.; Morselli-Labate, A.M.; Bianchi, G.; Bugianesi, E.; McCullough, A.J.; Forlani, G.; Melchionda, N. Association of nonalcoholic fatty liver disease with insulin resistance. Am. J. Med. 1999, 107, 450–455. [Google Scholar] [CrossRef]
- Browning, J.D.; Horton, J.D. Molecular mediators of hepatic steatosis and liver injury. J. Clin. Invest. 2004, 114, 147–152. [Google Scholar] [CrossRef]
- Schwimmer, J.B.; Behling, C.; Newbury, R.; Deutsch, R.; Nievergelt, C.; Schork, N.J.; Lavine, J.E. Histopathology of pediatric nonalcoholic fatty liver disease. Hepatology 2005, 42, 641–649. [Google Scholar] [CrossRef]
- Jansen, P.L. Non-alcoholic steatohepatitis. Eur. J. Gastroenterol. Hepatol. 2004, 16, 1079–1085. [Google Scholar] [CrossRef]
- Wanless, I.R.; Shiota, K. The pathogenesis of nonalcoholic steatohepatitis and other fatty liver diseases: A four-step model including the role of lipid release and hepatic venular obstruction in the progression to cirrhosis. Semin. Liver Dis. 2004, 24, 99–106. [Google Scholar] [CrossRef]
- Harrison, S.A.; Kadakia, S.; Lang, K.A.; Schenker, S. Nonalcoholic steatohepatitis: What we know in the new millennium. Am. J. Gastroenterol. 2002, 97, 2714–2724. [Google Scholar]
- Cortez-Pinto, H.; de Moura, M.C.; Day, C.P. Non-alcoholic steatohepatitis: From cell biology to clinical practice. J. Hepatol. 2006, 44, 197–208. [Google Scholar] [CrossRef]
- Anstee, Q.M.; Goldin, R.D. Mouse models in non-alcoholic fatty liver disease and steatohepatitis research. Int. J. Exp. Pathol. 2006, 87, 1–16. [Google Scholar] [CrossRef]
- Leclercq, I.A.; Farrell, G.C.; Field, J.; Bell, D.R.; Gonzalez, F.J.; Robertson, G.R. Cyp2e1 and cyp4a as microsomal catalysts of lipid peroxides in murine nonalcoholic steatohepatitis. J. Clin. Invest. 2000, 105, 1067–1075. [Google Scholar]
- Oliveira, C.P.; da Costa Gayotto, L.C.; Tatai, C.; Della Bina, B.I.; Janiszewski, M.; Lima, E.S.; Abdalla, D.S.; Lopasso, F.P.; Laurindo, F.R.; Laudanna, A.A. Oxidative stress in the pathogenesis of nonalcoholic fatty liver disease, in rats fed with a choline-deficient diet. J. Cell Mol. Med. 2002, 6, 399–406. [Google Scholar] [CrossRef]
- Tilg, H.; Diehl, A.M. Cytokines in alcoholic and nonalcoholic steatohepatitis. N. Engl. J. Med. 2000, 343, 1467–1476. [Google Scholar] [CrossRef]
- Polyzos, S.A.; Kountouras, J.; Zavos, C. Nonalcoholic fatty liver disease: The pathogenetic roles of insulin resistance and adipocytokines. Curr. Mol. Med. 2009, 9, 299–314. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R. Evolution of inflammation in nonalcoholic fatty liver disease: The multiple parallel hits hypothesis. Hepatology 2010, 52, 1836–1846. [Google Scholar] [CrossRef]
- Ikejima, K.; Takei, Y.; Honda, H.; Hirose, M.; Yoshikawa, M.; Zhang, Y.J.; Lang, T.; Fukuda, T.; Yamashina, S.; Kitamura, T.; et al. Leptin receptor-mediated signaling regulates hepatic fibrogenesis and remodeling of extracellular matrix in the rat. Gastroenterology 2002, 122, 1399–1410. [Google Scholar] [CrossRef]
- Bissell, D.M.; Roulot, D.; George, J. Transforming growth factor beta and the liver. Hepatology 2001, 34, 859–867. [Google Scholar] [CrossRef]
- Knittel, T.; Muller, L.; Saile, B.; Ramadori, G. Effect of tumour necrosis factor-alpha on proliferation, activation and protein synthesis of rat hepatic stellate cells. J. Hepatol. 1997, 27, 1067–1080. [Google Scholar] [CrossRef]
- Han, Y.P.; Tuan, T.L.; Hughes, M.; Wu, H.; Garner, W.L. Transforming growth factor-beta-and tumor necrosis factor-alpha -mediated induction and proteolytic activation of mmp-9 in human skin. J. Biol. Chem. 2001, 276, 22341–22350. [Google Scholar]
- Arthur, M.J. Fibrogenesis, II. Metalloproteinases and their inhibitors in liver fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G245–G249. [Google Scholar]
- Murphy, G.; Docherty, A.J. The matrix metalloproteinases and their inhibitors. Am. J. Respir. Cell Mol. Biol. 1992, 7, 120–125. [Google Scholar] [CrossRef]
- Arthur, M.J.; Friedman, S.L.; Roll, F.J.; Bissell, D.M. Lipocytes from normal rat liver release a neutral metalloproteinase that degrades basement membrane (type iv) collagen. J. Clin. Invest. 1989, 84, 1076–1085. [Google Scholar] [CrossRef]
- Knittel, T.; Mehde, M.; Kobold, D.; Saile, B.; Dinter, C.; Ramadori, G. Expression patterns of matrix metalloproteinases and their inhibitors in parenchymal and non-parenchymal cells of rat liver: Regulation by tnf-alpha and tgf-beta1. J. Hepatol. 1999, 30, 48–60. [Google Scholar] [CrossRef]
- Han, J.Y.; Fan, J.Y.; Horie, Y.; Miura, S.; Cui, D.H.; Ishii, H.; Hibi, T.; Tsuneki, H.; Kimura, I. Ameliorating effects of compounds derived from salvia miltiorrhiza root extract on microcirculatory disturbance and target organ injury by ischemia and reperfusion. Pharmacol. Ther. 2008, 117, 280–295. [Google Scholar] [CrossRef]
- Jang, S.I.; Kim, H.J.; Kim, Y.J.; Jeong, S.I.; You, Y.O. Tanshinone iia inhibits lps-induced nf-kappab activation in raw 264.7 cells: Possible involvement of the nik-ikk, erk1/2, p38 and jnk pathways. Eur. J. Pharmacol. 2006, 542, 1–7. [Google Scholar] [CrossRef]
- Ng, T.B.; Liu, F.; Wang, Z.T. Antioxidative activity of natural products from plants. Life Sci. 2000, 66, 709–723. [Google Scholar] [CrossRef]
- Sung, H.J.; Choi, S.M.; Yoon, Y.; An, K.S. Tanshinone iia, an ingredient of salvia miltiorrhiza bunge, induces apoptosis in human leukemia cell lines through the activation of caspase-3. Exp. Mol. Med. 1999, 31, 174–178. [Google Scholar] [CrossRef]
- Fan, G.W.; Gao, X.M.; Wang, H.; Zhu, Y.; Zhang, J.; Hu, L.M.; Su, Y.F.; Kang, L.Y.; Zhang, B.L. The anti-inflammatory activities of tanshinone iia, an active component of tcm, are mediated by estrogen receptor activation and inhibition of inos. J. Steroid Biochem. Mol. Biol. 2009, 113, 275–280. [Google Scholar] [CrossRef]
- Yin, H.Q.; Kim, Y.S.; Choi, Y.J.; Kim, Y.C.; Sohn, D.H.; Ryu, S.Y.; Lee, B.H. Effects of tanshinone iia on the hepatotoxicity and gene expression involved in alcoholic liver disease. Arch. Pharm. Res. 2008, 31, 659–665. [Google Scholar]
- Song, H.Y.; Zhang, L.; Pan, J.L.; Yang, L.L.; Ji, G. Bioactivity of five components of Chinese herbal formula jiangzhi granules against hepatocellular steatosis. J. Integr. Med. 2013, 11, 262–268. [Google Scholar] [CrossRef]
- Fang, Z.Y.; Lin, R.; Yuan, B.X.; Liu, Y.; Zhang, H. Tanshinone iia inhibits atherosclerotic plaque formation by down-regulating mmp-2 and mmp-9 expression in rabbits fed a high-fat diet. Life Sci. 2007, 81, 1339–1345. [Google Scholar] [CrossRef]
- Vizzutti, F.; Provenzano, A.; Galastri, S.; Milani, S.; Delogu, W.; Novo, E.; Caligiuri, A.; Zamara, E.; Arena, U.; Laffi, G.; et al. Curcumin limits the fibrogenic evolution of experimental steatohepatitis. Lab. Invest. 2010, 90, 104–115. [Google Scholar] [CrossRef]
- Kulinski, A.; Vance, D.E.; Vance, J.E. A choline-deficient diet in mice inhibits neither the cdp-choline pathway for phosphatidylcholine synthesis in hepatocytes nor apolipoprotein b secretion. J. Biol. Chem. 2004, 279, 23916–23924. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, J.; Liu, Q.; Harnish, D.C. Farnesoid x receptor agonist way-362450 attenuates liver inflammation and fibrosis in murine model of non-alcoholic steatohepatitis. J. Hepatol. 2009, 51, 380–388. [Google Scholar] [CrossRef]
- Gao, D.; Wei, C.; Chen, L.; Huang, J.; Yang, S.; Diehl, A.M. Oxidative DNA damage and DNA repair enzyme expression are inversely related in murine models of fatty liver disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G1070–G1077. [Google Scholar] [CrossRef]
- Gebhardt, R. Oxidative stress, plant-derived antioxidants and liver fibrosis. Planta Med. 2002, 68, 289–296. [Google Scholar] [CrossRef]
- Pociask, D.A.; Sime, P.J.; Brody, A.R. Asbestos-derived reactive oxygen species activate tgf-beta1. Lab. Invest. 2004, 84, 1013–1023. [Google Scholar] [CrossRef]
- Albright, C.D.; Salganik, R.I.; Craciunescu, C.N.; Mar, M.H.; Zeisel, S.H. Mitochondrial and microsomal derived reactive oxygen species mediate apoptosis induced by transforming growth factor-beta1 in immortalized rat hepatocytes. J. Cell Biochem. 2003, 89, 254–261. [Google Scholar] [CrossRef]
- Kayanoki, Y.; Fujii, J.; Suzuki, K.; Kawata, S.; Matsuzawa, Y.; Taniguchi, N. Suppression of antioxidative enzyme expression by transforming growth factor-beta 1 in rat hepatocytes. J. Biol. Chem. 1994, 269, 15488–15492. [Google Scholar]
- Brunt, E.M.; Kleiner, D.E.; Wilson, L.A.; Belt, P.; Neuschwander-Tetri, B.A.; Network, N.C.R. Nonalcoholic fatty liver disease (nafld) activity score and the histopathologic diagnosis in nafld: Distinct clinicopathologic meanings. Hepatology 2011, 53, 810–820. [Google Scholar] [CrossRef]
- Sample Availability: Samples of Standardized Salvia miltiorrhiza extract (SME) are available from the authors.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, H.S.; Son, W.-C.; Ryu, J.-E.; Koo, B.A.; Kim, Y.S. Standardized Salvia miltiorrhiza Extract Suppresses Hepatic Stellate Cell Activation and Attenuates Steatohepatitis Induced by a Methionine-Choline Deficient Diet in Mice. Molecules 2014, 19, 8189-8211. https://doi.org/10.3390/molecules19068189
Lee HS, Son W-C, Ryu J-E, Koo BA, Kim YS. Standardized Salvia miltiorrhiza Extract Suppresses Hepatic Stellate Cell Activation and Attenuates Steatohepatitis Induced by a Methionine-Choline Deficient Diet in Mice. Molecules. 2014; 19(6):8189-8211. https://doi.org/10.3390/molecules19068189
Chicago/Turabian StyleLee, Hak Sung, Woo-Chan Son, Jae-Eun Ryu, Bon Am Koo, and Yeong Shik Kim. 2014. "Standardized Salvia miltiorrhiza Extract Suppresses Hepatic Stellate Cell Activation and Attenuates Steatohepatitis Induced by a Methionine-Choline Deficient Diet in Mice" Molecules 19, no. 6: 8189-8211. https://doi.org/10.3390/molecules19068189