Matrilin-2 Is Proteolytically Cleaved by ADAMTS-4 and ADAMTS-5

Abstract
:1. Introduction
2. Results and Discussion
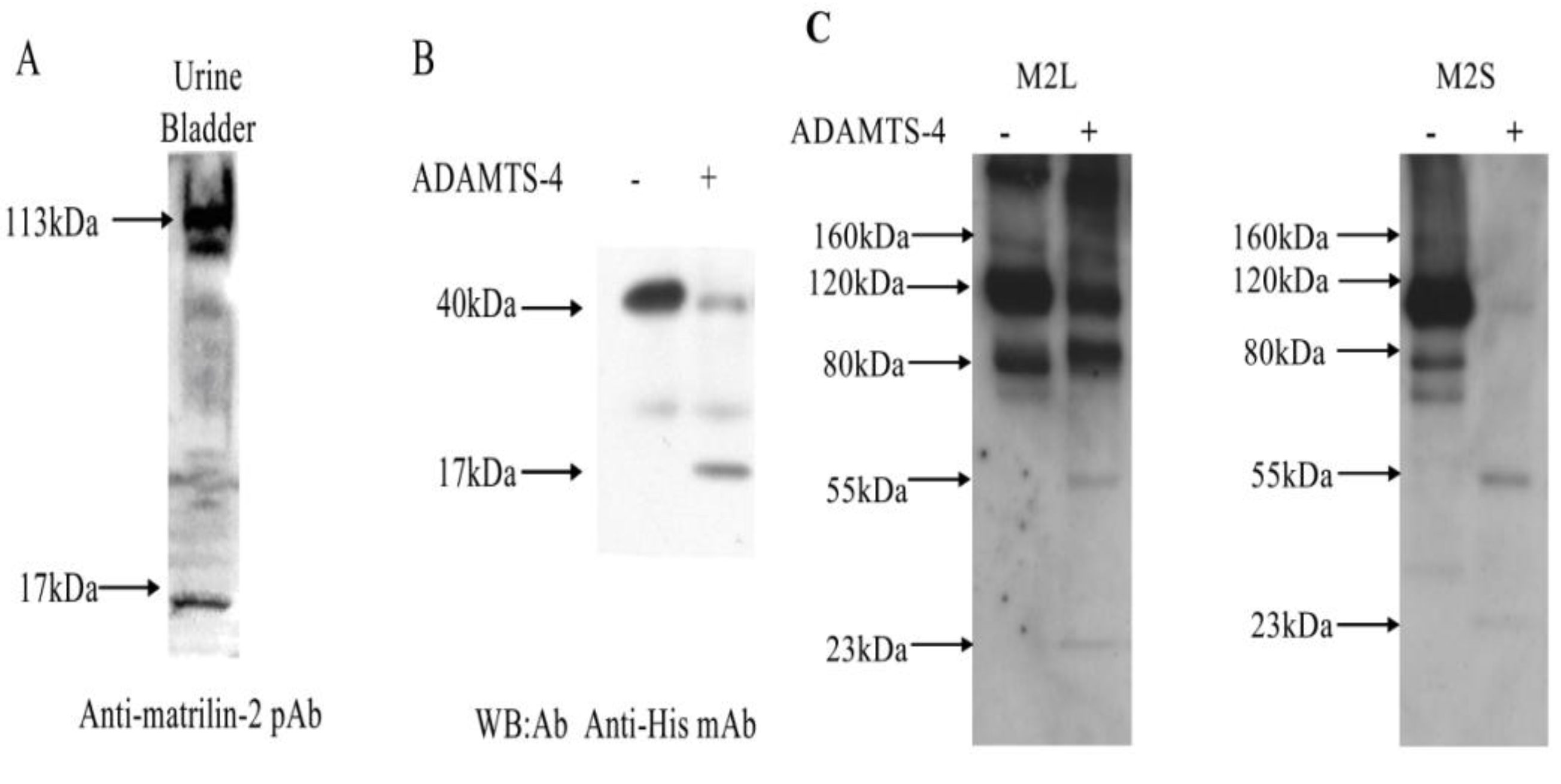
2.1. Proteolytic Processing of Matrilin-2
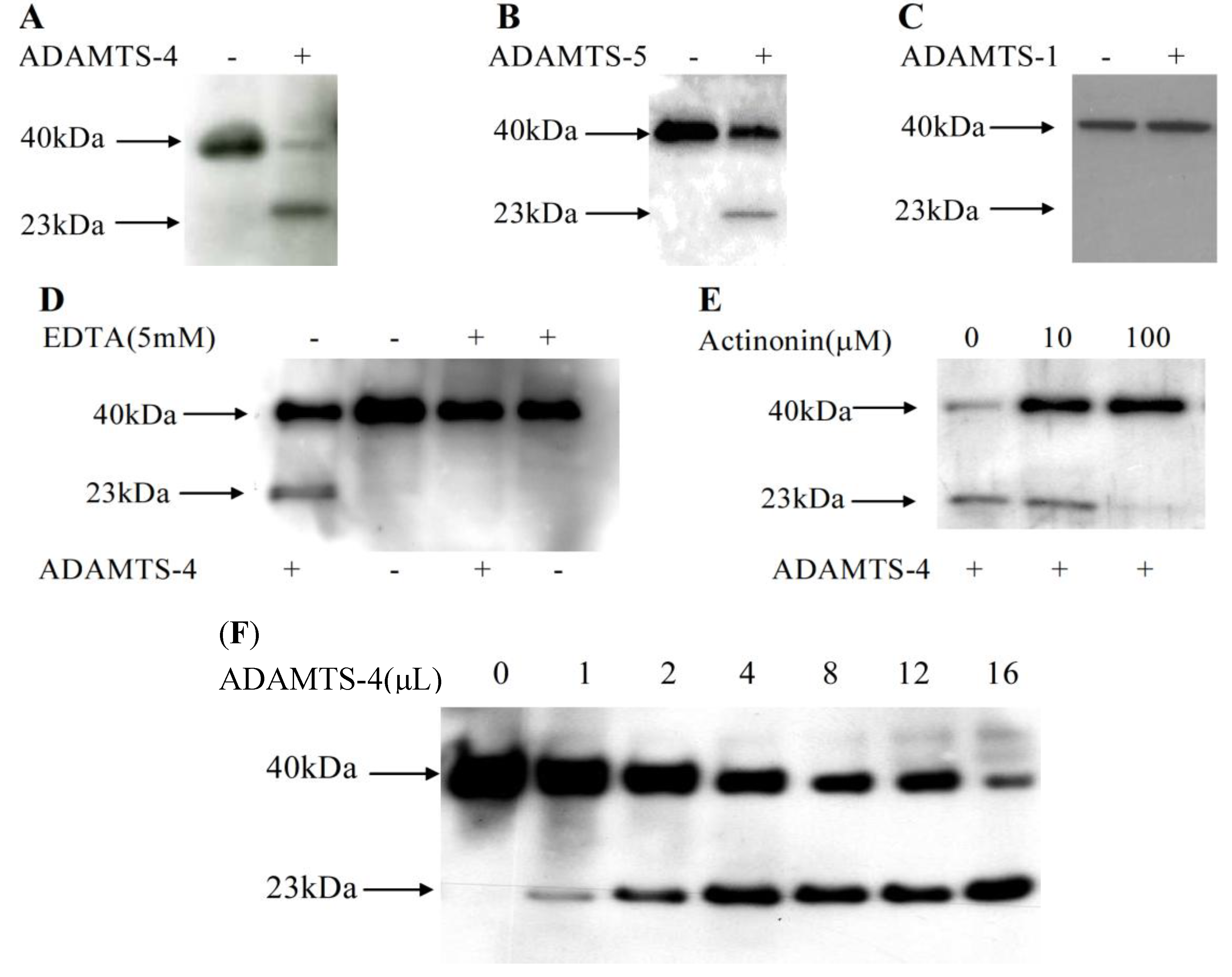
2.2. Matrilin-2 Was Cleaved by ADAMTS-4 and ADAMTS-5 but Not ADAMTS-1


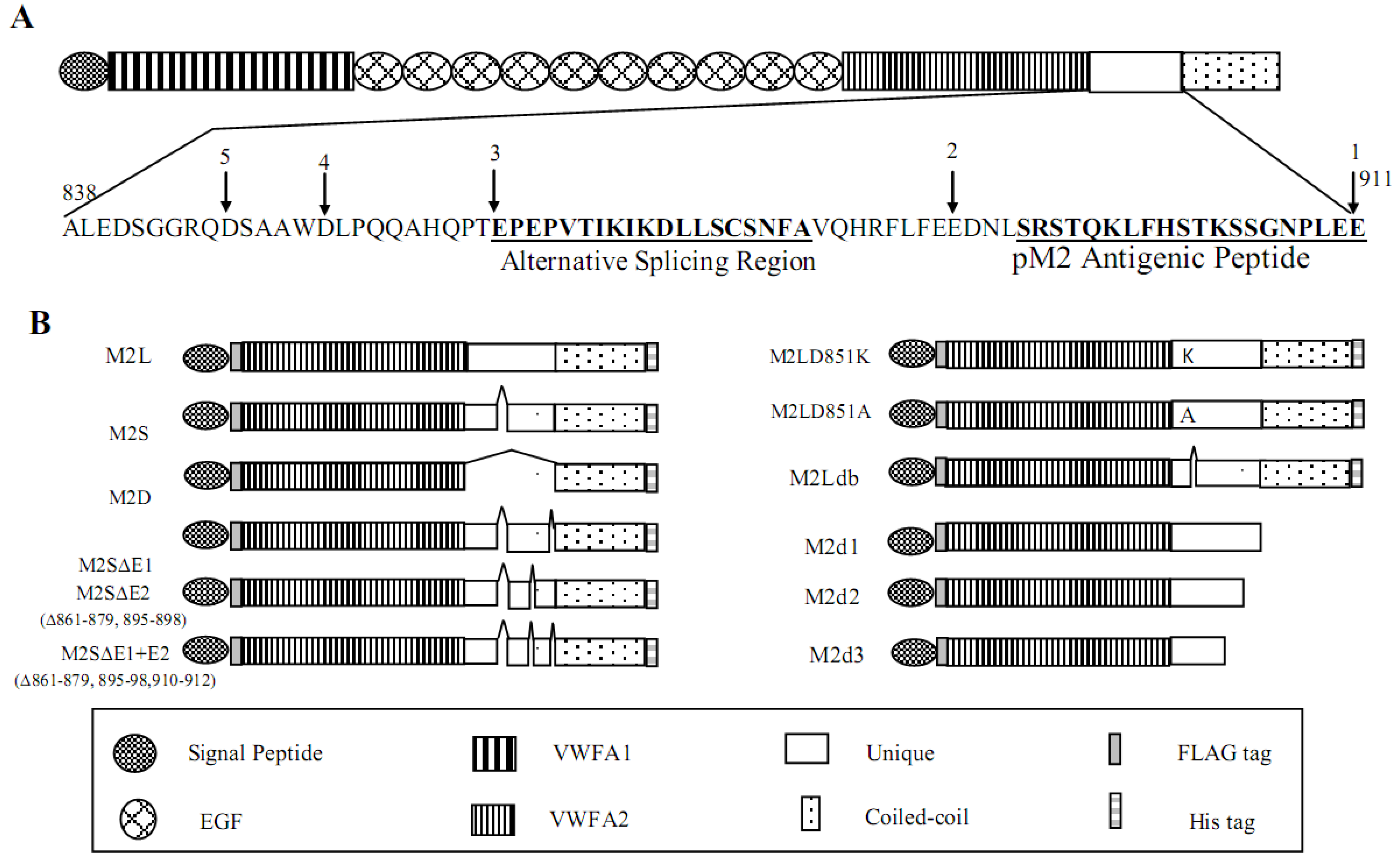
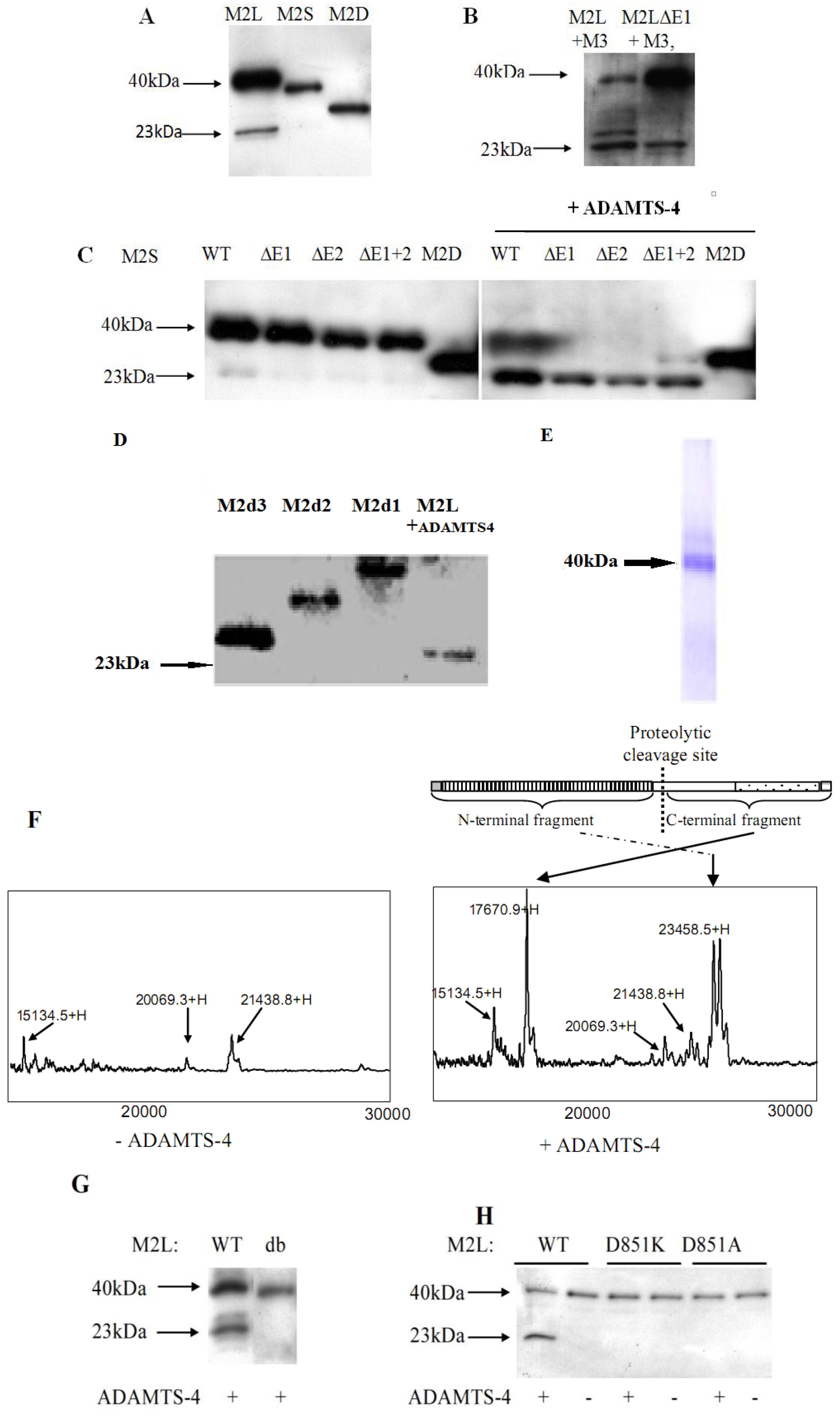
2.3. Identification of the Proteolytic Site in the Unique Domain of Matrilin-2


2.4. Matrilin-2 Proteolytic Cleavage in Vivo and the Proteolytic Susceptibility of the Splicing Variants

3. Experimental
3.1. Cloning and Construction of Matrilin-2 cDNAs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequences (5'–>3') | PCR Purpose |
|---|---|---|
| T7 | T AATACGACTCACTATAGGG | Cloning into pcDNA3.1/V5-His |
| BGH | CTAGAAGGCAACAGTCGAGG | |
| Mat2flgas | GCATCTTTTGTCATCATCGTCCTT | Adding a FLAG tag in matrilin-2 |
| Mat2flgs | GATGACAAAAAGAGATGCACTGAAGGC | |
| M2d1R | CGCTCGAGTCATTCTTCCAAAGGGTTCC | Creating M2d1 deletion mutant |
| M2d2R | CGCTCGAGTCATTCTTCAAACAGGAATC | Creating M2d2 deletion mutant |
| M2d3R | CGCTCGAGTCATTCTGTTGGCTGGTGGC | Creating M2d3 deletion mutant |
| M2Ldb2F | GACTCAGCAGCACAGCAGGCCCACCA | Creating M2Ldb mutant deleted 850WDLP853 |
| M2Ldb2R | GTGGGCCTGCTGTGCTGCTGAGTCCT | |
| M2LD851KF | AGCATGGAAAGCTGCCACA | Creating M2LD851K mutant |
| M2LD851KR | TGTGGCAGCTTCCATGCT | |
| M2LD851AF | AGCATGGGCCCTGCCACAGCAG | Creating M2LD851A mutant |
| M2LD851AR | CCTGCTGTGGCAGGGCCCATGCT | |
| DETA3F | TTCCTGTTTAATCTTTCACGGTCTACACA | M2SΔE2 (ΔGlu887 GluAsp889), M2SΔE1 + E2(ΔGlu887 GluAsp889, ΔGlu910 GluSer912) |
| DETA3R | GAAAAGTTAAACAGGAATCTATGTTGCA | |
| DETA4F | CCCTTTGCAGGACCAATGCAAATGTGA | Creating M2SΔE1 (ΔGlu910 GluSer912), M2SΔE1 + E2 (ΔGlu887 GluAsp889, ΔGlu910 GluSer912) |
| DETA4R | GTCCTGCAAAGGGTTTCCTGAAGATTT |
3.2. Transfection of Matrilin cDNAs
3.3. SDS-Polyacrylamide Gel Electrophoresis and Western Blot
3.4. In Vitro Analysis of Matrilin-2 Proteolysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Deak, F.; Wagener, R.; Kiss, I.; Paulsson, M. The matrilins: A novel family of oligomeric extracellular matrix proteins. Matrix Biol. 1999, 18, 55–64. [Google Scholar] [CrossRef]
- Wagener, R.; Ehlen, H.W.; Ko, Y.P.; Kobbe, B.; Mann, H.H.; Sengle, G.; Paulsson, M. The matrilins-adaptor proteins in the extracellular matrix. FEBS Lett. 2005, 579, 3323–3329. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, Y.; Johnson, D.M.; Goetinck, P.F. Assembly of a novel cartilage matrix protein filamentous network: Molecular basis of differential requirement of von Willebrand factor A domains. Mol. Biol. Cell 1999, 10, 2149–2162. [Google Scholar] [CrossRef]
- Wagener, R.; Kobbe, B.; Paulsson, M. Primary structure of matrilin-3, a new member of a family of extracellular matrix proteins related to cartilage matrix protein (matrilin-1) and von Willebrand factor. FEBS Lett. 1997, 413, 129–134. [Google Scholar] [CrossRef]
- Belluoccio, D.; Schenkera, T.; Baicib, A.; Trueba, B. Characterization of human matrilin-3 (MATN3). Genomics 1998, 53, 391–439. [Google Scholar] [CrossRef]
- Deak, F.; Piecha, D.; Bachrati, C.; Paulsson, M.; Kiss, I. Primary structure and expression of matrilin-2, the closest relative of cartilage matrix protein within the von Willebrand factor type A-like module superfamily. J. Biol. Chem. 1997, 272, 9268–9274. [Google Scholar]
- Klatt, A.R.; Nitsche, D.P.; Kobbe, B.; Macht, M.; Paulsson, M.; Wagener, R. Molecular structure, processing, and tissue distribution of matrilin-4. J. Biol. Chem. 2001, 276, 17267–17275. [Google Scholar]
- Wagener, R.; Kobbe, B.; Paulsson, M. Matrilin-4, a new member of the matrilin family of extracellular matrix proteins. FEBS Lett. 1998, 436, 123–127. [Google Scholar] [CrossRef]
- Klatt, A.R.; Paulsson, M.; Wagener, R. Expression of matrilins during maturation of mouse skeletal tissues. Matrix Biol. 2002, 21, 289–296. [Google Scholar] [CrossRef]
- Klatt, A.R.; Becker, A.K.; Neacsu, C.D.; Paulsson, M.; Wagener, R. The matrilins: Modulators of extracellular matrix assembly. Int. J. Biochem. Cell Biol. 2011, 43, 320–330. [Google Scholar] [CrossRef]
- Piecha, D.; Wiberg, C.; Mörgelin, M.; Reinhardt, D.P.; Deák, F.; Maurer, P.; Paulsson, M. Matrilin-2 interacts with itself and with other extracellular matrix proteins. Biochem. J. 2002, 367, 715–721. [Google Scholar] [CrossRef]
- Malin, D.; Sonnenberg-Riethmacher, E.; Guseva, D.; Wagener, R.; Aszódi, A.; Irintchev, A.; Riethmacher, D. The extracellular-matrix protein matrilin 2 participates in peripheral nerve regeneration. J. Cell Sci. 2009, 122, 995–1004. [Google Scholar] [CrossRef]
- Piecha, D.; Hartmann, K.; Kobbe, B.; Haase, I.; Mauch, C.; Krieg, T.; Paulsson, M. Expression of matrilin-2 in human skin. J. Investig. Dermatol. 2002, 119, 38–43. [Google Scholar] [CrossRef]
- Ichikawa, T.; Suenaga, Y.; Koda, T.; Ozaki, T.; Nakagawara, A. DeltaNp63/BMP-7-dependent expression of matrilin-2 is involved in keratinocyte migration in response to wounding. Biochem. Biophys. Res. Commun. 2008, 369, 994–1000. [Google Scholar] [CrossRef]
- Tortorella, M.D.; Burn, T.C.; Pratta, M.A.; Abbaszade, I.; Hollis, J.M.; Liu, R.; Rosenfeld, S.A.; Copeland, R.A.; Decicco, C.P.; Wynn, R.; et al. Purification and cloning of aggrecanase-1: A member of the ADAMTS family of proteins. Science 1999, 284, 1664–1666. [Google Scholar]
- Tortorella, M.D.; Pratta, M.; Liu, R.Q.; Austin, J.; Ross, O.H.; Abbaszade, I.; Burn, T.; Arner, E. Sites of aggrecan cleavage by recombinant human aggrecanase-1 (ADAMTS-4). J Biol. Chem. 2000, 275, 18566–18573. [Google Scholar]
- Curtis, C.L.; Rees, S.G.; Little, C.B.; Flannery, C.R.; Hughes, C.E.; Wilson, C.; Dent, C.M.; Otterness, I.G.; Harwood, J.L.; Caterson, B.; et al. Pathologic indicators of degradation and inflammation in human osteoarthritic cartilage are abrogated by exposure to n-3 fatty acids. Arthritis Rheumatol. 2002, 46, 1544–1553. [Google Scholar] [CrossRef]
- Arner, E.C. Aggrecanase-mediated cartilage degradation. Curr. Opin. Pharmacol. 2002, 2, 322–329. [Google Scholar] [CrossRef]
- Sandy, J.D.; Westling, J.; Kenagy, R.D.; Iruela-Arispe, M.L.; Verscharen, C.; Rodriguez-Mazaneque, J.C.; Zimmermann, D.R.; Lemire, J.M.; Fischer, J.W.; Wight, T.N.; et al. Versican V1 proteolysis in human aorta in vivo occurs at the Glu441-Ala442 bond, a site that is cleaved by recombinant ADAMTS-1 and ADAMTS-4. J. Biol. Chem. 2001, 276, 13372–13378. [Google Scholar] [CrossRef]
- Flannery, C.R.; Zeng, W.; Corcoran, C.; Collins-Racie, L.A.; Chockalingam, P.S.; Hebert, T.; Mackie, S.A.; McDonagh, T.; Crawford, T.K.; Tomkinson, K.N.; et al. Autocatalytic cleavage of ADAMTS-4 (Aggrecanase-1) reveals multiple glycosaminoglycan-binding sites. J. Biol. Chem. 2002, 277, 42775–42780. [Google Scholar] [CrossRef]
- Tortorella, M.; Pratta, M.; Liu, R.-Q.; Abbaszade, I.; Ross, H.; Burn, T.; Arner, E. The thrombospondin motif of aggrecanase-1 (ADAMTS-4) is critical for aggrecan substrate recognition and cleavage. J. Biol. Chem. 2000, 275, 25791–25797. [Google Scholar]
- Makihira, S.; Yan, W.; Ohno, S.; Kawamoto, T.; Fujimoto, K.; Okimura, A.; Yoshida, E.; Noshiro, M.; Hamada, T.; Kato, Y.; et al. Enhancement of cell adhesion and spreading by a cartilage-specific noncollagenous protein, cartilage matrix protein (CMP/Matrilin-1), via integrin alpha1beta1. J. Biol. Chem. 1999, 274, 11417–11423. [Google Scholar]
- Winterbottom, N.; Tondravi, M.M.; Harrington, T.L.; Klier, F.G.; Vertel, B.M.; Goetinck, P.F. Cartilage matrix protein is a component of the collagen fibril of cartilage. Dev. Dyn. 1992, 193, 266–276. [Google Scholar] [CrossRef]
- Chen, Q.; Johnson, D.M.; Haudenschild, D.R.; Tondravi, M.M.; Goetinck, P.F. Cartilage matrix protein forms a type II collagen-independent filamentous network: Analysis in primary cell cultures with a retrovirus expression system. Mol. Biol. Cell 1995, 6, 1743–1753. [Google Scholar] [CrossRef]
- Paulsson, M.; Heinegard, D. Matrix proteins bound to associatively prepared proteoglycans from bovine cartilage. Biochem. J. 1979, 183, 539–545. [Google Scholar]
- Wiberg, C.; Klatt, A.R.; Wagener, R.; Paulsson, M.; Bateman, J.F.; Heinegård, D.; Mörgelin, M. Complexes of matrilin-1 and biglycan or decorin connect collagen VI microfibrils to both collagen II and aggrecan. J. Biol. Chem. 2003, 278, 37698–37704. [Google Scholar]
- Wu, J.J.; Eyre, D.R. Matrilin-3 forms disulfide-linked oligomers with matrilin-1 in bovine epiphyseal cartilage. J. Biol. Chem. 1998, 273, 17433–17438. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Q. Changes of matrilin forms during endochondral ossification. Molecular basis of oligomeric assembly. J. Biol. Chem. 2000, 275, 32628–32634. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.-K.; Luo, J.-M.; Kanbe, K.; Chen, Q. Multiple functions of the von Willebrand Factor A domain in matrilins: Secretion, assembly, and proteolysis. J. Orthop. Surg. Res. 2008, 3, 21. [Google Scholar] [CrossRef]
- Hills, R.; Mazzarella, R.; Fok, K.; Liu, M.; Nemirovskiy, O.; Leone, J.; Zack, M.D.; Arner, E.C.; Viswanathan, M.; Abujoub, A.; et al. Identification of an ADAMTS-4 cleavage motif using phage display leads to the development of fluorogenic peptide substrates and reveals matrilin-3 as a novel substrate. J. Biol. Chem. 2007, 282, 11101–11109. [Google Scholar] [CrossRef]
- Ehlen, H.W.; Sengle, G.; Klatt, A.R.; Talke, A.; Müller, S.; Paulsson, M.; Wagener, R. Proteolytic processing causes extensive heterogeneity of tissue matrilin forms. J. Biol. Chem. 2009, 284, 21545–21556. [Google Scholar]
- Li, L.; Zhang, L.; Shao, Y.; Wang, G.; Gong, R.; Wang, Z.; Peng, J.; Wang, S.; Genochio, D.; Zhao, B.; et al. Distinct roles of two alternative splice variants of matrilin-2 in protein oligomerization and proteolysis. Mol. Med. Rep. 2012, 6, 1204–1210. [Google Scholar]
- Hughes, C.E.; Little, C.B.; Büttner, F.H.; Bartnik, E.; Caterson, B. Differential expression of aggrecanase and matrix metalloproteinase activity in chondrocytes isolated from bovine and porcine articular cartilage. J. Biol. Chem. 1998, 273, 30576–30582. [Google Scholar] [CrossRef]
- Nakamura, H.; Fujii, Y.; Inoki, I.; Sugimoto, K.; Tanzawa, K.; Matsuki, H.; Miura, R.; Yamaguchi, Y.; Okada, Y. Brevican is degraded by matrix metalloproteinases and aggrecanase-1 (ADAMTS4) at different sites. J. Biol. Chem. 2000, 275, 38885–38890. [Google Scholar] [CrossRef]
- Westling, J.; Gottschall, P.E.; Thompson, V.P.; Cockburn, A.; Perides, G.; Zimmermann, D.R.; Sandy, J.D. ADAMTS4 (aggrecanase-1) cleaves human brain versican V2 at Glu405-Gln406 to generate glial hyaluronate binding protein. Biochem. J. 2004, 377, 787–795. [Google Scholar] [CrossRef]
- Matthews, R.T.; Gary, S.C.; Zerillo, C.; Pratta, M.; Solomon, K.; Arner, E.C.; Hockfield, S. Brain-enriched hyaluronan binding (BEHAB)/brevican cleavage in a glioma cell line is mediated by a disintegrin and metalloproteinase with thrombospondin motifs (ADAMTS) family member. J. Biol. Chem. 2000, 275, 22695–22703. [Google Scholar]
- Tortorella, M.D.; Arner, E.C.; Hills, R.; Easton, A.; Korte-Sarfaty, J.; Fok, K.; Wittwer, A.J.; Liu, R.Q.; Malfait, A.M. α-Macroglobulin is a novel substrate for ADAMTS-4 and ADAMTS-5and represents an endogenous inhibitor of these enzymes. J. Biol. Chem. 2004, 279, 17554–17561. [Google Scholar] [CrossRef]
- Westling, J.; Fosang, A.J.; Last, K.; Thompson, V.P.; Tomkinson, K.N.; Hebert, T.; McDonagh, T.; Collins-Racie, L.A.; LaVallie, E.R.; Morris, E.A.; et al. ADAMTS4 cleaves at the aggrecanase site (Glu373-Ala374) and secondarily at the matrix metalloproteinase site (Asn341–Phe342) in the aggrecan interglobular domain. J. Biol. Chem. 2002, 277, 16059–16066. [Google Scholar] [CrossRef]
- Piecha, D.; Kiss, I.; Paulsson, M.; Deák, F.; Hauser, N.; Studer, D.; Muratoglu, S.; Mörgelin, M. Matrilin-2, a large, oligomeric matrix protein, is expressed by a great variety of cells and forms fibrillar networks. J. Biol. Chem. 1999, 274, 13353–13361. [Google Scholar]
- Jayasuriya, C.T.; Goldring, M.B.; Terek, R.; Chen, Q. Matrilin-3 induction of IL-1 receptor antagonist Is required for up-regulating collagen II and aggrecan and down-regulating ADAMTS-5 gene expression. Arthritis Res. Ther. 2012. [Google Scholar] [CrossRef]
- Lefebvre, V.; Garofalo, S.; de Crombrugghe, B. Type X collagen gene expression in mouse chondrocytes immortalized by a temperature-sensitive simian virus 40 large tumor antigen. J. Cell Biol. 1995, 128, 239–245. [Google Scholar] [CrossRef]
- Chen, Q.; Johnson, D.M.; Haudenschild, D.R.; Goetinck, P.F. Progression and recapitulation of the chondrocyte differentiation program: Cartilage matrix protein is a marker for cartilage maturation. Dev. Biol. 1995, 172, 293–306. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Q. The noncollagenous domain 1 of type X collagen. A novel motif for trimer and higher order multimer formation without a triple helix. J. Biol. Chem. 1999, 274, 22409–22413. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds and enzymes are ordered from the companies and are not available from the authors.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Z.; Luo, J.; Iwamoto, S.; Chen, Q. Matrilin-2 Is Proteolytically Cleaved by ADAMTS-4 and ADAMTS-5. Molecules 2014, 19, 8472-8487. https://doi.org/10.3390/molecules19068472
Wang Z, Luo J, Iwamoto S, Chen Q. Matrilin-2 Is Proteolytically Cleaved by ADAMTS-4 and ADAMTS-5. Molecules. 2014; 19(6):8472-8487. https://doi.org/10.3390/molecules19068472
Chicago/Turabian StyleWang, Zhengke, Junming Luo, Satori Iwamoto, and Qian Chen. 2014. "Matrilin-2 Is Proteolytically Cleaved by ADAMTS-4 and ADAMTS-5" Molecules 19, no. 6: 8472-8487. https://doi.org/10.3390/molecules19068472