Cytotoxic and Antiproliferative Effect of Tepary Bean Lectins on C33-A, MCF-7, SKNSH, and SW480 Cell Lines


 , , , and
, , , and 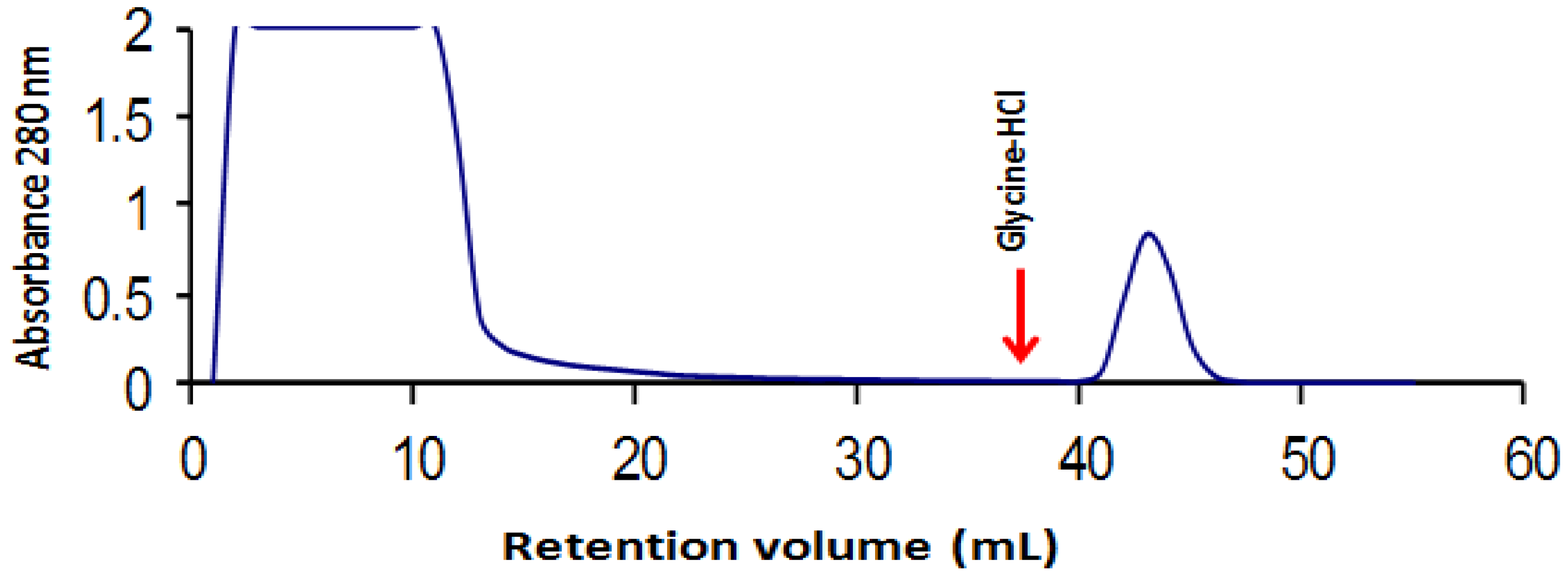{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Tepary Bean Lectin Purification

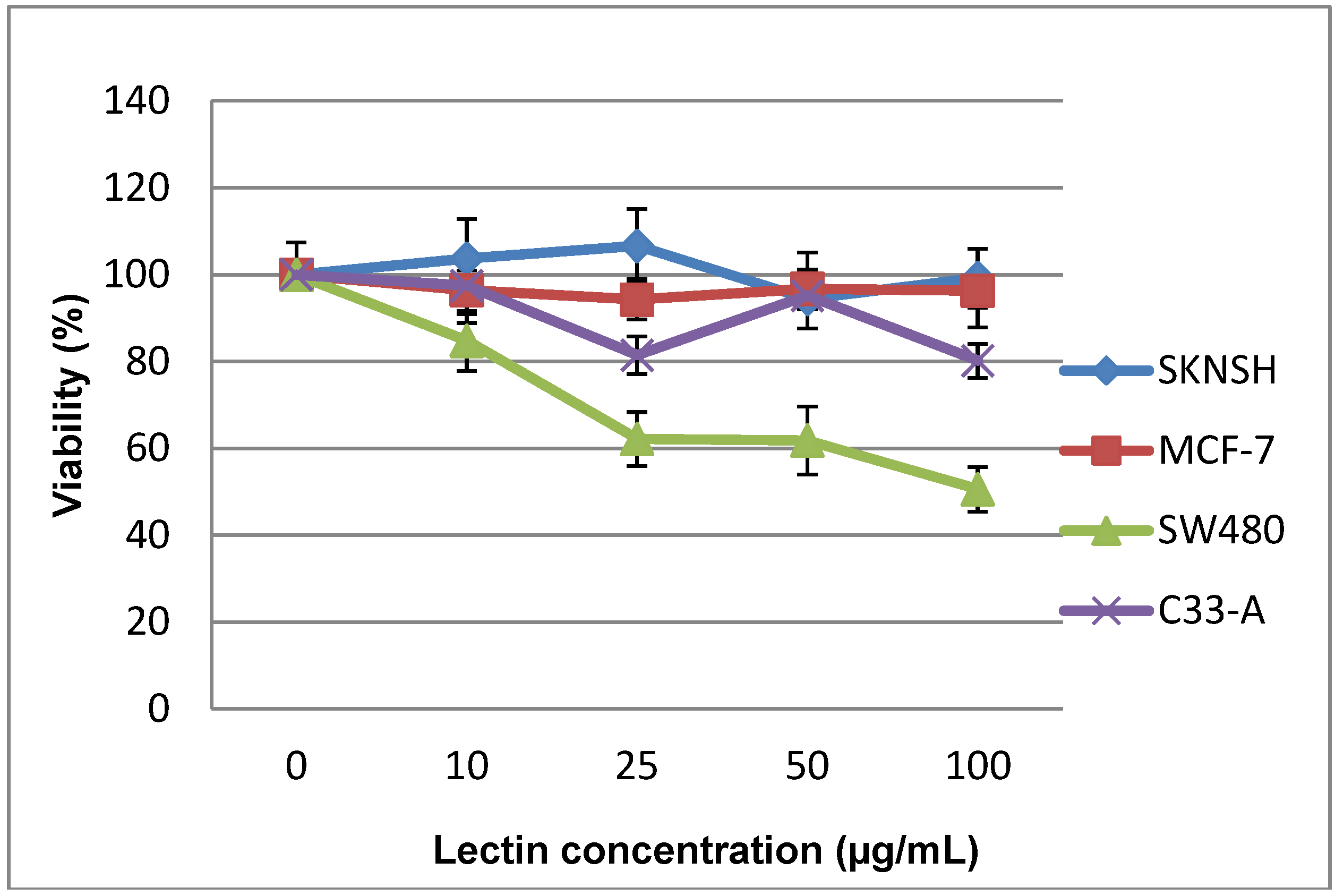
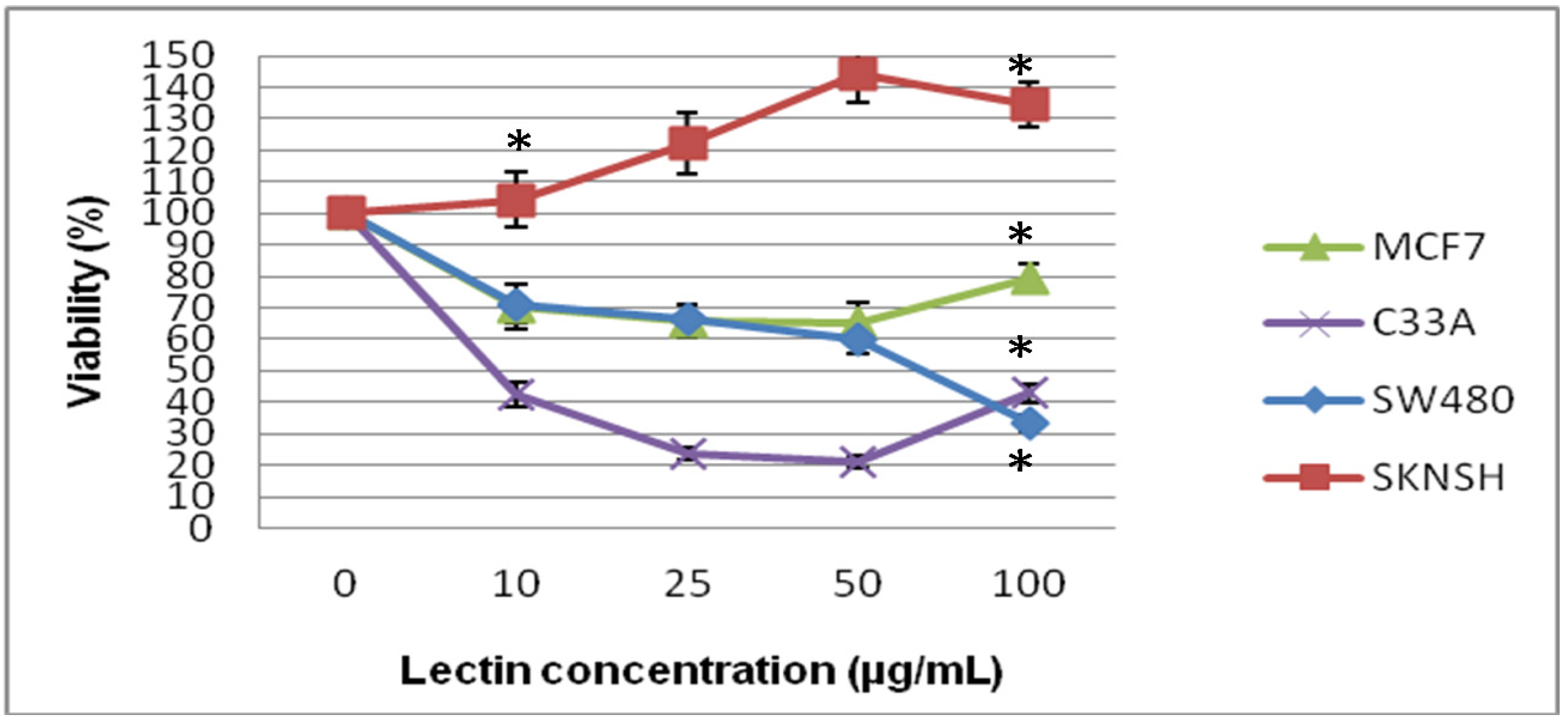
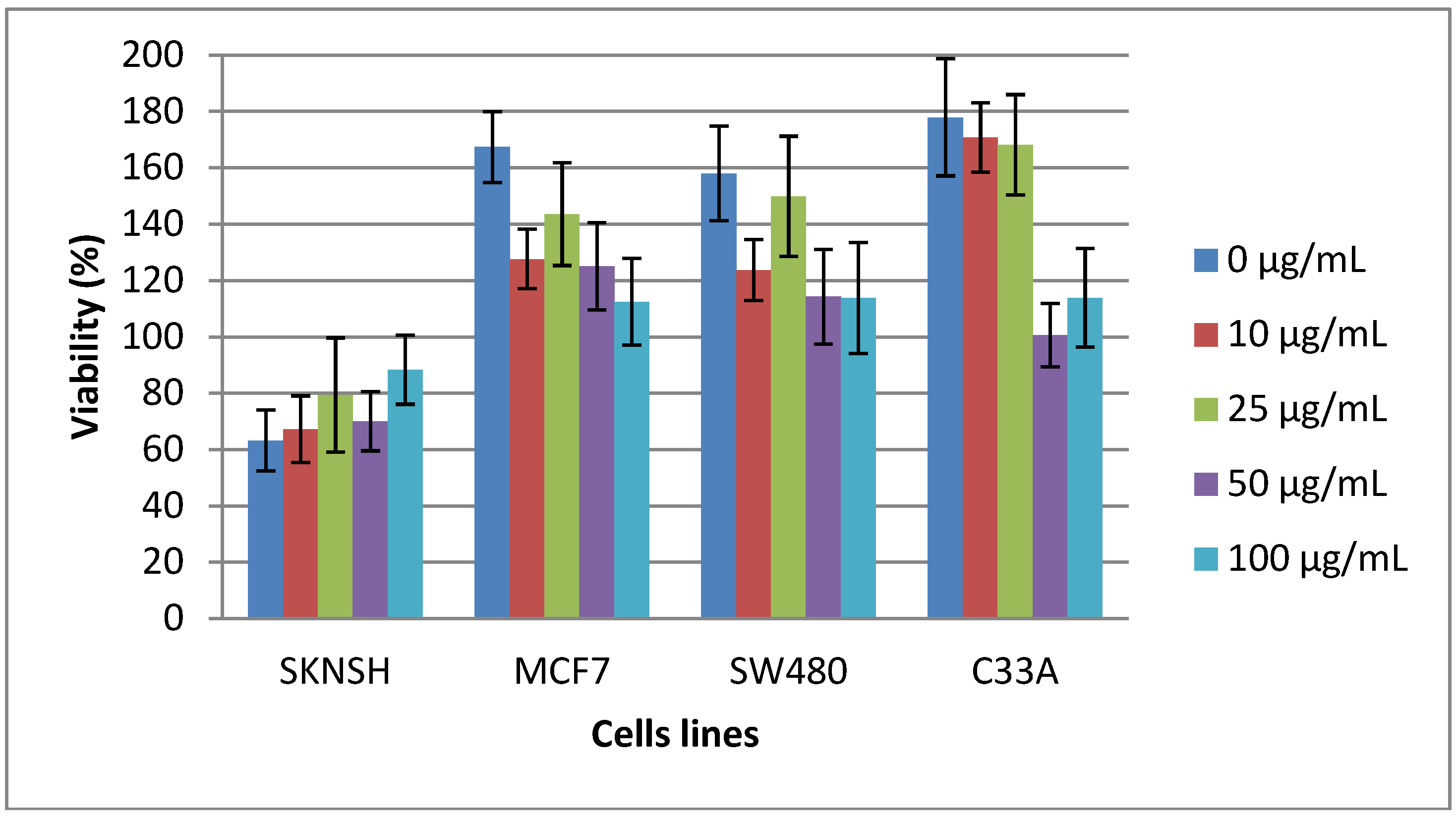
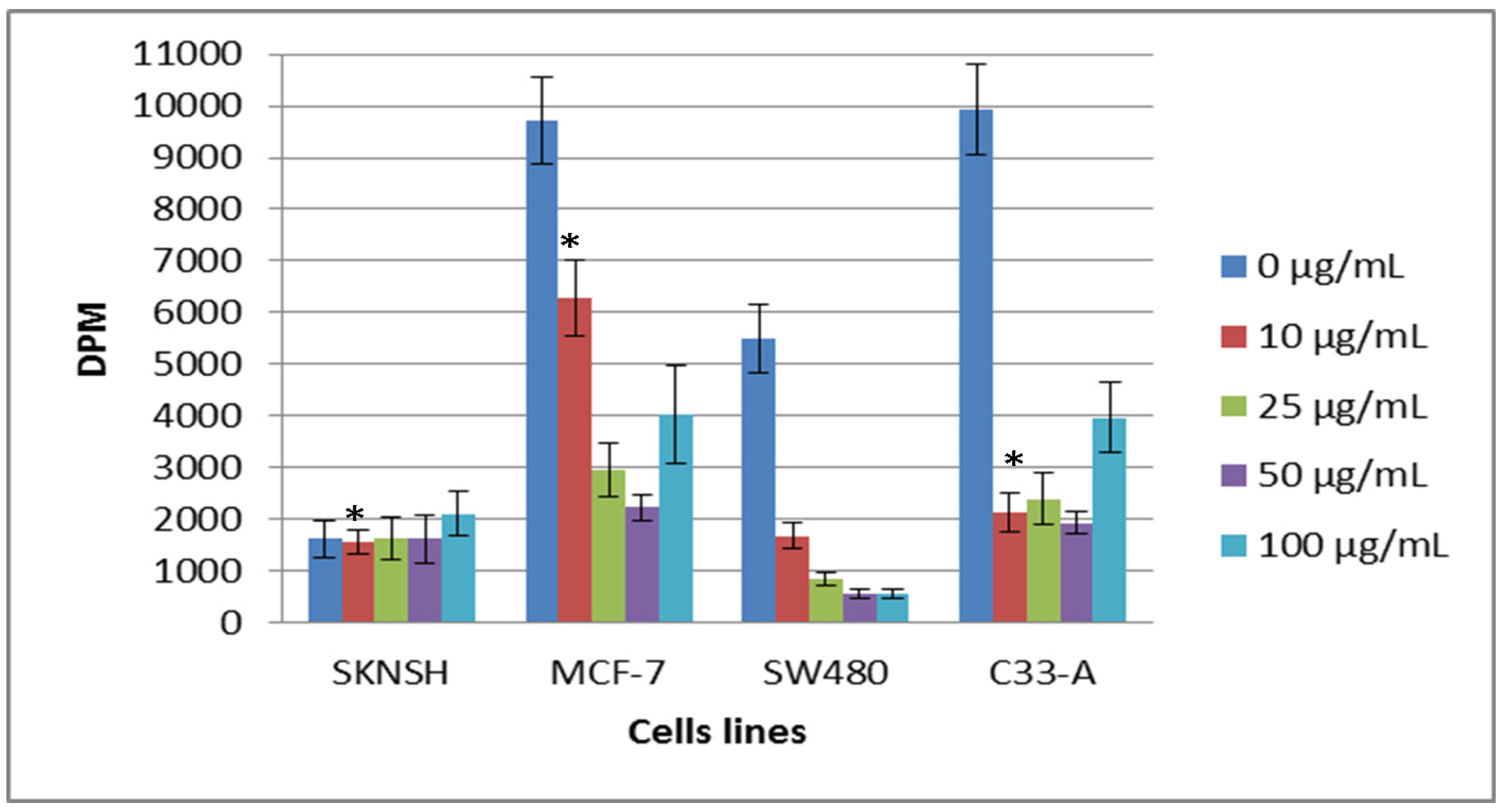
2.2. Cytotoxic and Antiproliferative Effect of the Tepary Bean Lectin (TBL)






3. Discussion
4. Experimental Section
4.1. Extraction and Purification of Lectins
4.2. Cell Culture
4.3. Cytotoxic and Antiproliferative Assay
4.3.1. MTT Assay
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Conflicts of Interest
Acknowledgements
References
- Grassi, D.; Desideri, G.; di Giosia, P.; de Feo, M.; Fellini, E.; Cheli, P.; Ferri, L.; Ferri, C. Tea, flavonoids, and cardiovascular health: Endothelial protection. Am. J. Clin. Nutr. 2013, 98, 1660S–1666S. [Google Scholar]
- Jansen, R.J.; Robinson, D.P.; Stolzenberg-Solomon, R.Z.; Bamlet, W.R.; de Andrade, M.; Oberg, A.L.; Rabe, K.G.; Anderson, K.E.; Olson, J.E.; Sinha, R.; et al. Nutrients from fruit and vegetable consumption reduce the risk of pancreatic cancer. J. Gastrointest. Cancer 2013, 44, 152–161. [Google Scholar] [CrossRef]
- Behzad, S.; Pirani, A.; Mosaddegh, M. Cytotoxic activity of some medicinal plants from hamedan district of Iran. Iran. J. Pharm. Res. 2014, 13, 199–205. [Google Scholar]
- Castillo-Villanueva, A.; Abdullaev, F. Plant lectins and their effects on cancer. Rev. Invest. Clin. 2005, 57, 55–64. [Google Scholar]
- De Mejía, E.G.; Prisecaru, V.I. Lectins as bioactive plant proteins: A potential in cancer treatment. Crit. Rev. Food Sci. Nutr. 2005, 45, 425–445. [Google Scholar] [CrossRef]
- Vargas-Albores, F.; de la Fuente, G.; Agundis, C.; Córdoba, F. Purification and characterization of a lectin from Phaseolus acutifolius var. Latifolius. Prep. Biochem. 1987, 17, 379–396. [Google Scholar]
- Valadez-Vega, C.; Guzmán-Partida, A.M.; Soto-Cordova, F.J.; Alvarez-Manilla, G.; Morales-González, J.A.; Madrigal-Santillán, E.; Villagómez-Ibarra, J.R.; Zúñiga-Pérez, C.; Gutiérrez-Salinas, J.; Becerril-Flores, M.A. Purification, biochemical characterization, and bioactive properties of a lectin purified from the seeds of whitetepary bean (phaseolus acutifolius variety latifolius). Molecules 2011, 21, 2561–2582. [Google Scholar]
- Leite, J.F.; Assreuy, A.M.; Mota, M.R.; Bringel, P.H.; E Lacerda, R.R.; Gomes Vde, M.; Cajazeiras, J.B.; do Nascimento, K.S.; Pessôa Hde, L.; Gadelha, C.A.; et al. Antinociceptive and Anti-inflammatory Effects of a Lectin-Like Substance from Clitoria fairchildiana R. Howard Seeds. Molecules 2012, 17, 3277–3290. [Google Scholar] [CrossRef]
- Pinto-Júnior, V.R.; de Santiago, M.Q.; Osterne, V.J.; Correia, J.L.; Pereira-Júnior, F.N.; Cajazeiras, J.B.; de Vasconcelos, M.A.; Teixeira, E.H.; do Nascimento, A.S.; Miguel, T.B.; et al. Purification, partial characterization and immobilization of a mannose-specific lectin from seeds of Dioclea lasiophylla mart. Molecules 2013, 18, 10857–10869. [Google Scholar] [CrossRef]
- Alborés, S.; Mora, P.; Bustamante, M.J.; Cerdeiras, M.P.; Franco Fraguas, L. Purification and applications of a lectin from the mushroom Gymnopilus spectabilis. Appl. Biochem. Biotechnol. 2014, 172, 2081–2090. [Google Scholar] [CrossRef]
- Campo, E.; Condom, E.; Palacín, A.; Quesada, E.; Cardesa, A. Lectin binding patterns in normal and neoplastic colonic mucosa. A study of Dolichos biflorus agglutinin, peanut agglutinin, and wheat germ agglutinin. Dis. Colon Rectum. 1988, 31, 892–899. [Google Scholar]
- Langkilde, N.C.; Wolf, H.; Orntoft, T.F. Binding of wheat and peanut lectins to human transitional cell carcinomas. Correlation with histopathologic grade, invasion, and DNA ploidy. Cancer 1989, 64, 849–853. [Google Scholar] [CrossRef]
- Zhang, S.L.; Liang, Y.R.; Li, J.L.; Dai, Y.R.; Huang, D. Preliminary studies of serum glycoconjugates in patients with cancer using the enzyme-linked lectin assay. Cancer Biochem. Biophys. 1990, 11, 211–216. [Google Scholar]
- Wong, S.E.; Winbanks, C.E.; Samuel, C.S.; Hewitson, T.D. Lectin histochemistry for light and electron microscopy. Methods Mol. Biol. 2010, 611, 103–114. [Google Scholar] [CrossRef]
- Goldstein, I.J.; Hughes, C.; Monsigny, M.; Osawa, T.; Sharon, N. What should be called a lectin? Nature 1980, 285, 66. [Google Scholar]
- Willy, J.P.; van Damme, E.J. Lectins as PIant Defense Proteins. Plant. Physiol. 1995, 109, 347–352. [Google Scholar]
- Sharon, N.; Lis, H. How proteins bind carbohydrates: Lessons from legume lectins. J. Agric. Food Chem. 2002, 50, 6586–6591. [Google Scholar] [CrossRef]
- Huang, L.H.; Yan, Q.J.; Kopparapu, N.K.; Jiang, Z.Q.; Sun, Y. Astragalus membranaceus lectin (AML) induces caspase-dependent apoptosis in human leukemia cells. Cell. Prolif. 2012, 45, 15–21. [Google Scholar] [CrossRef]
- Kabir, S.R.; Reza, M.A. Antibacterial activity of kaempferia rotunda rhizome lectin and its induction of apoptosis in ehrlich ascites carcinoma cells. Appl. Biochem. Biotechnol. 2014, 172, 2866–2876. [Google Scholar] [CrossRef]
- Hivrale, A.; Ingale, A. Plant as a plenteous reserve of lectin. Plant Signal. Behav. 2013, 8, e26595. [Google Scholar] [CrossRef]
- Regente, M.; Taveira, G.B.; Pinedo, M.; Elizalde, M.M.; Ticchi, A.J.; Diz, M.S.; Carvalho, A.O.; de la Canal, L.; Gomes, V.M. A sunflower Lectin with Antifungal Properties and Putative Medical Mycology Applications. Curr. Microbiol. 2014. [Google Scholar] [CrossRef]
- Reynoso-Camacho, R.; González de Mejía, E.; Loarca-Piña, G. Purification and acute toxicity of a lectin extracted from tepary bean (Phaseolus acutifolius). Food Chem. Toxicol. 2003, 41, 21–27. [Google Scholar] [CrossRef]
- Mody, R.; Joshi, S.; Chaney, W. Use of lectins as diagnostic and therapeutic tools for cancer. Pharmacol. Toxicol. Methods 1995, 33, 1–10. [Google Scholar] [CrossRef]
- Gabius, H.; Kaltner, H. Lectin-based therapy applications from the laboratory to practice. Berl. Munch Tierarztl. Wochenschr. 1994, 107, 376–381. [Google Scholar]
- Wu, L.; Yang, X.; Duan, X.; Cui, L.; Li, G. Exogenous expression of marine lectins DlFBL and SpRBL induces cancer cell apoptosis possibly through PRMT5-E2F-1 pathway. Sci. Rep. 2014, 4, 4505. [Google Scholar]
- Lis, H.; Sharon, N. Review. Protein glycosylation, Structural and functional aspects. Eur. J. Biochem. 1993, 218, 1–27. [Google Scholar]
- Sharon, N.; Lis, H. Carbohydrates in cell recognition. Sci. Am. 1993, 268, 82–89. [Google Scholar] [CrossRef]
- Varki, A.; Freeze, H.H.; Vacquier, V.D. Glycans in development and systemic physiology. In Essentials of Glycobiology; Varki, A., Cummings, R., Esko, J., Freeze, H., Hart, G., Marth, J., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009. [Google Scholar]
- Spiro, R.G. Protein glycosylation: Nature, distribution, enzymatic formation, and disease implications of glycopeptide bonds. Glycobiology 2002, 12, 43R–56R. [Google Scholar] [CrossRef]
- Milde-Langosch, K.; Karn, T.; Schmidt, M.; Zu Eulenburg, C.; Oliveira-Ferrer, L.; Wirtz, R.M.; Schumacher, U.; Witzel, I.; Schütze, D.; Müller, V. Prognostic relevance of glycosylation-associated genes in breast cancer. Breast Cancer Res. Treat. 2014. [Google Scholar] [CrossRef]
- Varki, A.; Kannagi, R.; Toole, B.P. Glycosylation changes in cancer. In Essentials of Glycobiology; Varki, A., Cummings, R., Esko, J., Freeze, H., Hart, G., Marth, J., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009. [Google Scholar]
- Tuccillo, F.M.; de Laurentiis, A.; Palmieri, C.; Fiume, G.; Bonelli, P.; Borrelli, A.; Tassone, P.; Scala, I.; Buonaguro, F.M.; Quinto, I.; Scala, G. Aberrant Glycosylation as Biomarker for Cancer: Focus on CD43. Biomed. Res. Int. 2014, 2014, 742831. [Google Scholar]
- Nagata, Y. Function and structure of fungal lectins. Chem. Biol. 2000, 38, 368–373. [Google Scholar]
- Tucker-Burden, C.; Chappa, P.; Krishnamoorthy, M.; Gerwe, B.A.; Scharer, C.D.; Heimburg-Molinaro, J.; Harris, W.; Usta, S.N.; Eilertson, C.D.; Hadjipanayis, C.G.; et al. Lectins Identify Glycan Biomarkers on Glioblastoma Derived Cancer Stem Cells. Stem Cells Dev. 2012, 21, 2374. [Google Scholar]
- Peiris, D.; Markiv, A.; Curley, G.P.; Dwek, M.V. A novel approach to determining the affinity of protein-carbohydrate interactions employing adherent cancer cells grown on a biosensor surface. Biosens. Bioelectron. 2012, 35, 160. [Google Scholar] [CrossRef]
- Rambaruth, N.D.; Greenwell, P.; Dwek, M.V. The lectin Helix pomatia agglutinin recognises O-GlcNAc containing glycoproteins in human breast cancer. Glycobiology 2012, 22, 839–848. [Google Scholar] [CrossRef]
- Hernández, P.; Pérez, E.; Martínez, L.; Ortiz, B.; Martínez, G. Las lectinas vegetales como modelo de estudio de las interacciones proteína-carbohidrato. REB 2005, 24, 21–27. [Google Scholar]
- Brooks, S.A.; Hall, D.M.; Buley, I. GalNAc glycoprotein expression by breast cell lines, primary breast cancer and normal breast epithelial membrane. Br. J. Cancer. 2001, 85, 1014–1022. [Google Scholar] [CrossRef]
- Chrispeels, M.J. Glycobiology of plant cells. In Essential of Glycobiology, 2nd ed.; Varki, A., Cummings, R., Esko, J., Freeze, H., Hart, G., Marth, J., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1999. [Google Scholar]
- Nishimura, H.; Nishimura, M.; Oda, R.; Yamanaka, K.; Matsubara, T.; Ozaki, Y.; Sekiya, K.; Hamada, T.; Kato, Y. Lectins induce resistance to proteases and/or mechanical stimulus in all examined cells—Including bone marrow mesenchymal stem cells—On various scaffolds. Exp. Cell. Res. 2004, 15, 119–27. [Google Scholar]
- Kuwahara, I.; Ikebuchi, K.; Hamada, H.; Niitsu, Y.; Miyazawa, K.; Ohyashiki, K.; Fujisawa, H.; Furukawa, K. Changes in N-glycosylation of human stromal cells by telomerase expression. Biochem. Biophys. Res. Commun. 2003, 301, 293–297. [Google Scholar]
- Lyu, S.Y.; Choi, S.H.; Park, W.B. Korean mistletoe lectin-induced apoptosis in hepatocarcinoma cells is associated with inhibition of telomerase via mitochondrial controlled pathway independent of p53. Arch. Pharm. Res. 2002, 25, 93–101. [Google Scholar] [CrossRef]
- Kidd, P. A New Approach to Metastatic Cancer Prevention: Modified Citrus Pectin that Blocks Cell Surface Lectins. Altern. Med. Rev. 1996, 1, 4–10. [Google Scholar]
- Sehgal, P.; Kumar, O.; Kameswararao, M.; Ravindran, J.; Khan, M.; Sharma, S.; Vijayaraghavan, R.; Prasad, G.B. Differential toxicity profile of ricin isoforms correlates with their glycosylation levels. Toxicology 2011, 282, 56–67. [Google Scholar] [CrossRef]
- Zhou, W.; Gao, Y.; Xu, S.; Yang, Z.; Xu, T. Purification of a mannose-binding lectin Pinellia ternata agglutinin and its induction of apoptosis in Bel-7404 cells. Protein Expr. Purif. 2014, 93, 11–17. [Google Scholar]
- Pryme, I.F.; Bardocz, S. Anti-cancer therapy: Diversion of polyamines in the gut. Eur. J. Gastroenterol. Hepatol. 2001, 13, 1041–1046. [Google Scholar] [CrossRef]
- Abdullaev, F.I.; de Mejia, E.G. Antitumor effect of plant lectins. Nat. Toxins. 1997, 5, 157–163. [Google Scholar]
- Valadez-Vega, C.; Alvarez-Manilla, G.; Riverón-Negrete, L.; García-Carrancá, A.; Morales-González, J.A.; Zuñiga-Pérez, C.; Madrigal-Santillán, E.; Esquivel-Soto, J.; Esquivel-Chirino, C.; Villagómez-Ibarra, R.; et al. Detection of cytotoxic activity of lectin on human colon adenocarcinoma (Sw480) and epithelial cervical carcinoma (C33-A). Molecules 2011, 16, 2107–2118. [Google Scholar]
- De Mejia, E.G.; Valadez-Vega, M.; Reynoso-Camacho, R.; Loarca-Pina, G. Tannins, trypsin inhibitors and lectin cytotoxicity in Tepary (Phaseolus acutifolius) and Common (Phaseolus vulgaris) Beans. Plant. Foods Hum. Nutrit. 2005, 60, 137–145. [Google Scholar] [CrossRef]
- García-Gasca, T.; García-Cruz, M.; Hernandez-Rivera, E.; López-Matínez, J.; Castañeda-Cuevas, A.L.; Yllescas-Gasca, L.; Rodríguez-Méndez, A.J.; Mendiola-Olaya, E.; Castro-Guillén, J.L.; Blanco-Labra, A. Effects of Tepary bean (Phaseolus acutifolius) protease inhibitor and semipure lectin fractions on cancer cells. Nutr. Cancer 2012, 64, 1269–1278. [Google Scholar]
- López-Sánchez, C.; López-Martínez, F.J.; Castañeda-Cuevas, A.L.; Yllescas-Gasca, L.; Ferriz-Martínez, R.A.; Torres-Arteaga, I.C.; García-Gasca, T.; Gallegos-Corona, M.A.; Rodríguez-Méndez, A.J.; Mendiola-Olaya, E.; et al. Evaluation of in vitro and in vivo toxicity of lectins tepary bean. CIENCIA UAQ 2010, 3, 3–13. [Google Scholar]
- Zhao, Y.; Ahmad, A.M.; Randy Cheung, C.F.; Ng, T.B. Isolation and Characterization of a Lectin from Japanese Mottled Beans. Protein Pept. Lett. 2014, 21, 696–704. [Google Scholar]
- Ang, A.S.; Cheung, R.C.; Dan, X.; Chan, Y.S.; Pan, W.; Ng, T.B. Purification and characterization of a glucosamine-binding antifungal lectin from Phaseolus vulgaris cv. Chinese pinto beans with antiproliferative activity towards nasopharyngeal carcinoma cells. Appl. Biochem. Biotechnol. 2014, 172, 672–686. [Google Scholar]
- Osterne, V.J.; Santiago, M.Q.; Pinto-Junior, V.R.; Cajazeiras, J.B.; Correia, J.L.; Leitão, C.C.; Carneiro, R.F.; Pereira-Junior, F.N.; Vasconcelos, M.A.; Rocha, B.A.; et al. Purification, Partial Characterization, and CNBr-Sepharose Immobilization of a Vasorelaxant Glucose/Mannose Lectin from Canavalia virosa Seeds. Appl. Biochem. Biotechnol. 2014, 172, 3342–3353. [Google Scholar]
- Sehgal, P.; Khan, M.; Kumar, O.; Vijayaraghavan, R. Purification, characterization and toxicity profile of ricin isoforms from castor beans. Food Chem. Toxicol. 2010, 48, 3171–3176. [Google Scholar] [CrossRef]
- Pusztai, A.; Watt, W.; Stewart, J.C. Erythro and Lymphoagglutinins of Phaseolus Acutifolius. Phytochemistry 1987, 26, 1009–1013. [Google Scholar] [CrossRef]
- Pusztai, A.; Watt, W.B. Isolectins of Phaseolus vulgaris. A comprehensive study of fractionation. Biochim. Biophys. Acta 1974, 13, 57–71. [Google Scholar] [CrossRef]
- Kumar, P.; Patil, D.N.; Chaudhary, A.; Tomar, S.; Yernool, D.; Singh, N.; Dasauni, P.; Kundu, S.; Kumar, P. Purification and biophysical characterization of an 11S globulin from Wrightia tinctoria exhibiting hemagglutinating activity. Protein Pept. Lett. 2013, 20, 499–509. [Google Scholar] [CrossRef]
- Li, T.; Yin, X.; Liu, D.; Ma, X.; Lv, H.; Sun, S. Isolation and characterization of a novel lectin with antifungal and antiproliferative activities from Sophora alopecuroides seeds. Acta Biochim. Biophys. Sin. (Shanghai) 2012, 44, 606–613. [Google Scholar] [CrossRef]
- Stanley, P.; Carver, P. Lectin receptors and lectin resistance in Chinese hamster ovary cells. Adv. Exp. Med. Biol. 1977, 84, 265–284. [Google Scholar] [CrossRef]
- Li, W.P.; Zuber, C.; Roth, J. Use of Phaseolus vulgaris leukoagglutinating lectin in histochemical and blotting techniques: A comparison of Digoxigenin and biotin labeled lectins. Histochemistry 1993, 100, 347–354. [Google Scholar]
- Cumming, R.D.; Kornfeld, S. Characterization of the structural determinants required for the High affinity interaction of asparagine-linked oligosaccharides with immobilized Phaseolus vulgaris leukoagglutinating and erythroagglutinating lectins. J. Biol. Chem. 1982, 257, 11230–11234. [Google Scholar]
- Lotan, R.; Beattie, G.; Hubbell, W.; Nicholson, G.L. Activities of lectins and their immobilized derivatives in detergent solutions. Implication on the use lectin affinity chromatography for the purification of membrane glycoproteins. Biochemistry 1991, 16, 1787–1794. [Google Scholar]
- Bantel, H.; Engels, I.H.; Voelter, W.; Schulze-Osthoff, K.; Wesselborg, S. Mistletoes lectins activates caspase-8/FLICE independently of death receptor signaling and enhances anticancer drug-inducen apoptosis. Cancer Res. 1999, 59, 2083–2090. [Google Scholar]
- Büssing, A.; Stein, G.M.; Pfüller, U.; Schietzel, M. Differential binding of toxic lectins from Viscum album L., ML I and ML III, to human lymphocytes. Anticancer Res. 1999, 19, 5095–5099. [Google Scholar]
- Liu, B.; Xu, X.C.; Cheng, Y.; Huang, J.; Liu, Y.H.; Liu, Z.; Min, M.W.; Bian, H.J.; Chen, J.; Bao, J.K. Apoptosis-inducing effect and structural basis of Polygonatum cyrtonema lectin and chemical modification properties on its mannose-binding sites. BMB Rep. 2008, 41, 369–375. [Google Scholar] [CrossRef]
- Gong, F.; Ma, Y.; Ma, A.; Yu, Q.; Zhang, J.; Nie, H.; Chen, X.; Shen, B.; Li, N.; Zhang, D. A lectin from Chinese mistletoe increases gammadelta T cell-mediated cytotoxicity through induction of caspase-dependent apoptosis. Acta Biochim. Biophys. Sin. 2007, 39, 445–452. [Google Scholar]
- Joshi, S.; Tomes, D.; Pirruccello, S.; Gabius, H. Galactoside specific VAA lectin-induced apoptosis in human leukemia cells. Proc. Am. Assn. Cancer Res. 1994, 35, 6. [Google Scholar]
- Hansen, M.B.; Nielsen, S.E.; Berg, K. Re-examination and further development of a precise and rapid dye method for measuring cell growth/cell kill. J. Immunol. Methods 1989, 119, 203–210. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are not available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valadez-Vega, C.; Morales-González, J.A.; Sumaya-Martínez, M.T.; Delgado-Olivares, L.; Cruz-Castañeda, A.; Bautista, M.; Sánchez-Gutiérrez, M.; Zuñiga-Pérez, C. Cytotoxic and Antiproliferative Effect of Tepary Bean Lectins on C33-A, MCF-7, SKNSH, and SW480 Cell Lines. Molecules 2014, 19, 9610-9627. https://doi.org/10.3390/molecules19079610
Valadez-Vega C, Morales-González JA, Sumaya-Martínez MT, Delgado-Olivares L, Cruz-Castañeda A, Bautista M, Sánchez-Gutiérrez M, Zuñiga-Pérez C. Cytotoxic and Antiproliferative Effect of Tepary Bean Lectins on C33-A, MCF-7, SKNSH, and SW480 Cell Lines. Molecules. 2014; 19(7):9610-9627. https://doi.org/10.3390/molecules19079610
Chicago/Turabian StyleValadez-Vega, Carmen, José A. Morales-González, María Teresa Sumaya-Martínez, Luis Delgado-Olivares, Areli Cruz-Castañeda, Mirandeli Bautista, Manuel Sánchez-Gutiérrez, and Clara Zuñiga-Pérez. 2014. "Cytotoxic and Antiproliferative Effect of Tepary Bean Lectins on C33-A, MCF-7, SKNSH, and SW480 Cell Lines" Molecules 19, no. 7: 9610-9627. https://doi.org/10.3390/molecules19079610