Effect of Chlorogenic Acid on Melanogenesis of B16 Melanoma Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
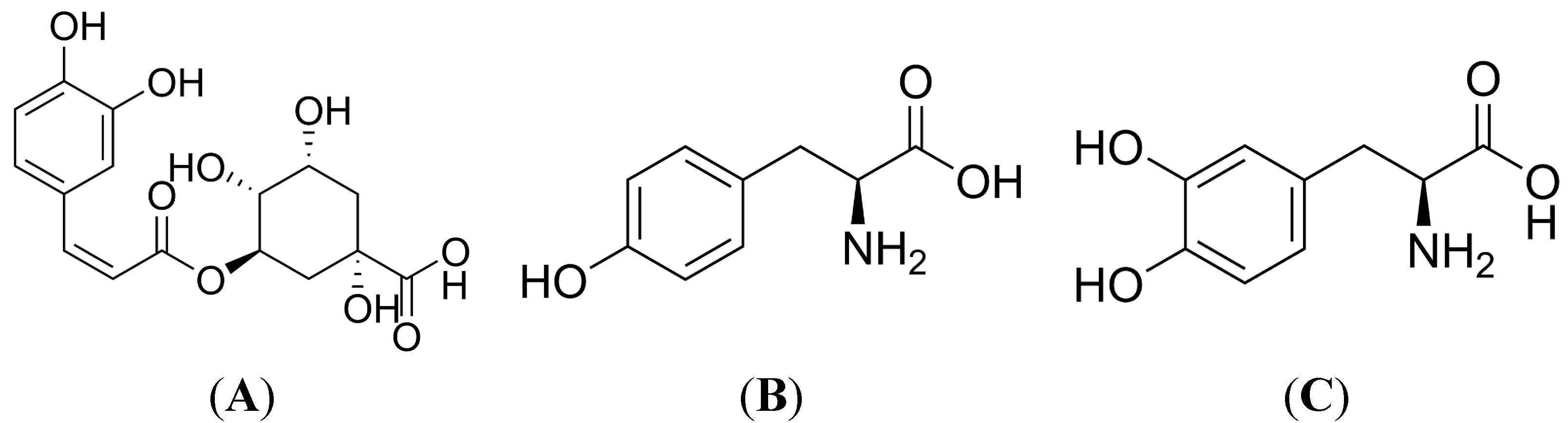
:1. Introduction

2. Results and Discussion
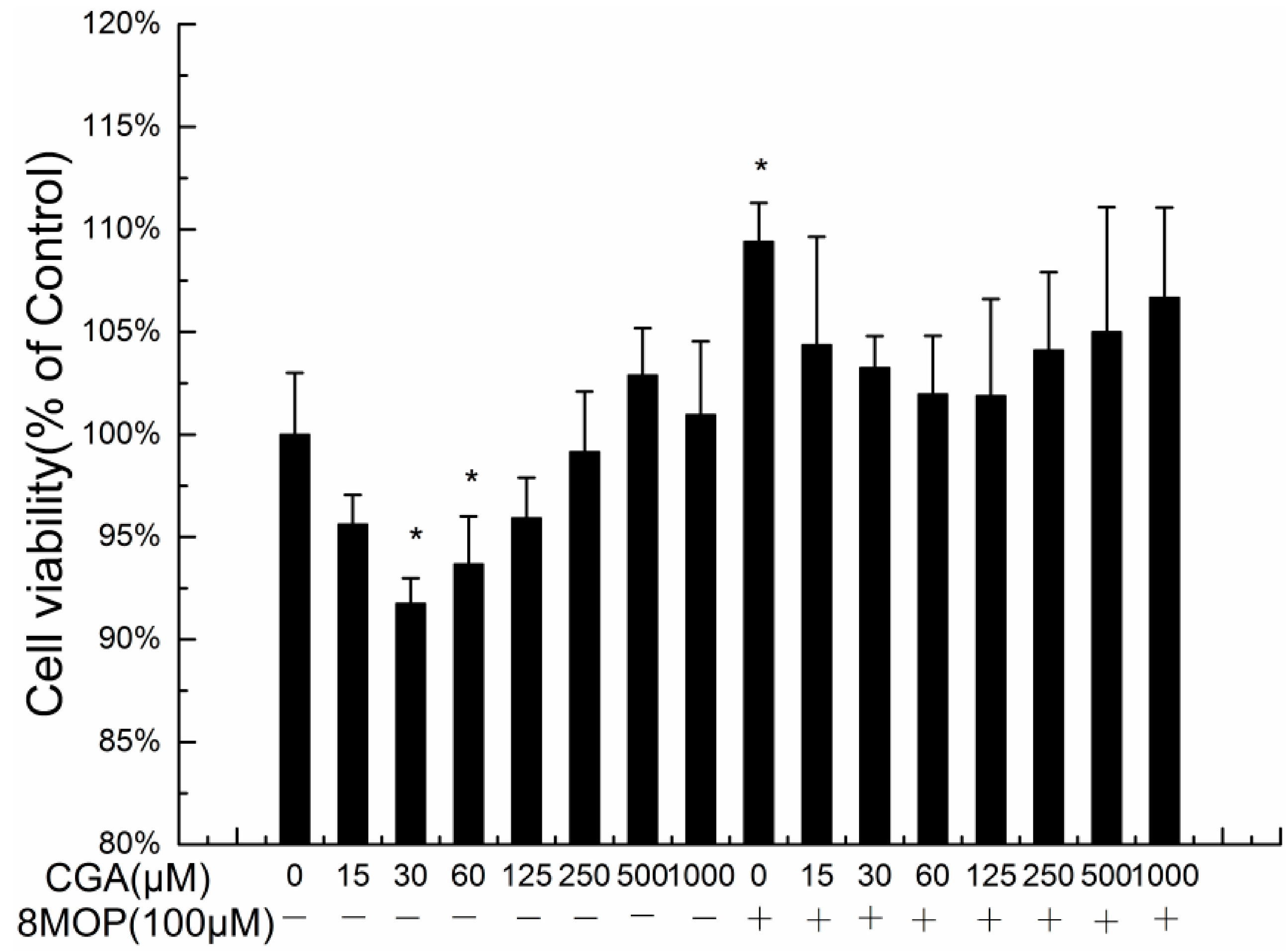
2.1. Effects of CGA on the Proliferation of B16 Melanoma Cells

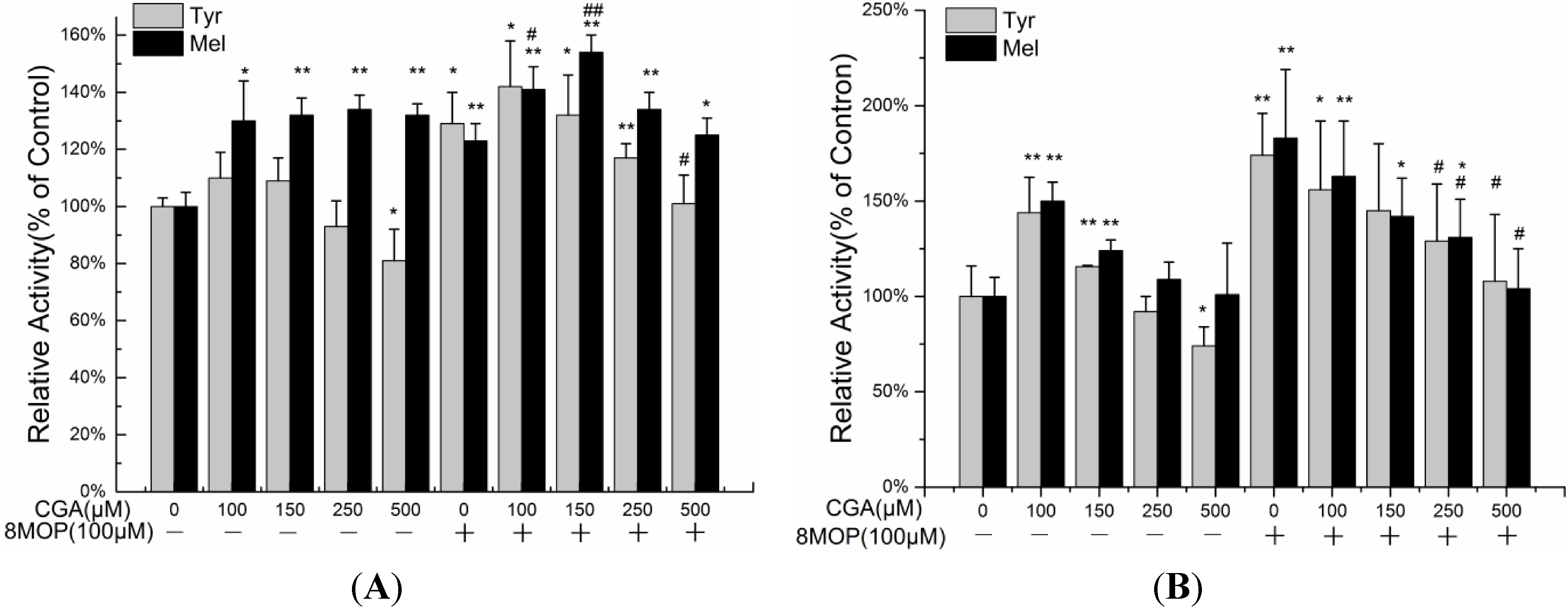
2.2. Effects of CGA on Melanin Production and Tyrosinase Activity

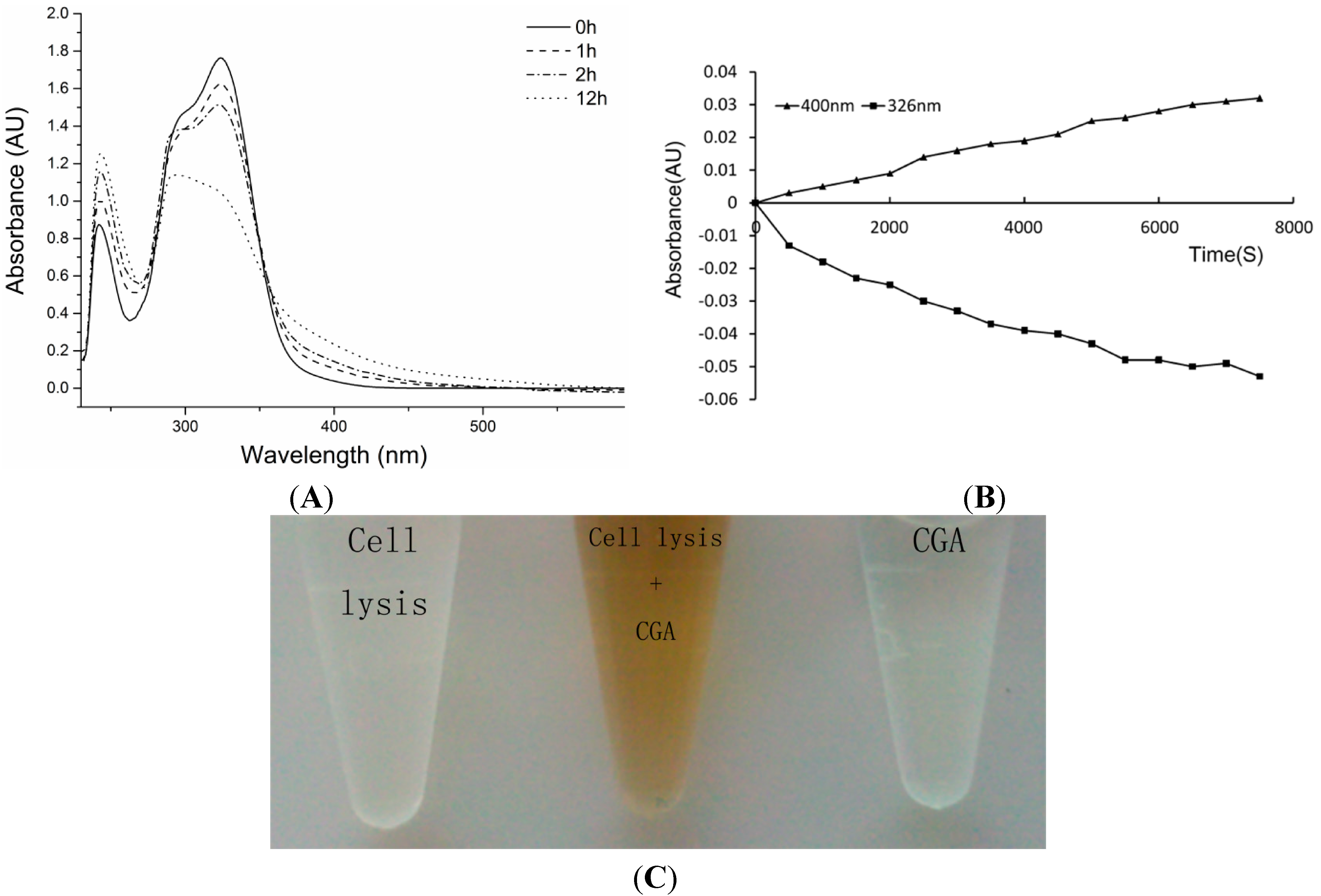
2.3. Effects of CGA on Cells Lysis Solution Enzymatic Reaction Assay

3. Experimental Section
3.1. Materials
3.2. Cell Culture and MTT Assay
3.3. Tyrosinase Activity
3.4. Melanin Content
3.5. Enzymatic Reaction
3.6. General Procedures
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Upadhyay, R.; Mohan Rao, L.J. An outlook on chlorogenic acids—Occurrence, chemistry, technology, and biological activities. Crit. Rev. Food Sci. Nutr. 2013, 53, 968–984. [Google Scholar] [CrossRef]
- Scharffetter-Kochanek, K.; Wlaschek, M.; Brenneisen, P.; Schauen, M.; Blaudschun, R.; Wenk, J. UV-induced reactive oxygen species in photocarcinogenesis and photoaging. Biol. Chem. 1997, 378, 1247–1257. [Google Scholar]
- Gaudout, D.; Megard, D.; Inisan, C.; Esteve, C.; Lejard, F. Phloridzin-Rich Phenolic Fraction and Use Thereof as a Cosmetic, Dietary or Nutraceutical Agent. U.S. Patent 7427418 B2, 23 September 2008. [Google Scholar]
- Gaudout, D.; Megard, D.; Lejard, F. Use of a Dihydrochalcone-Rich Phenolic Fraction in a Cosmetic Treatment. U.S. Patent 7285298, 23 October 2007. [Google Scholar]
- Kim, Y.-J.; Uyama, H. Tyrosinase inhibitors from natural and synthetic sources: Structure, inhibition mechanism and perspective for the future. Cell. Mol. Life Sci. CMLS 2005, 62, 1707–1723. [Google Scholar] [CrossRef]
- Lee, B.W.; Schwartz, R.A.; Hercogová, J.; Valle, Y.; Lotti, T.M. Vitiligo road map. Dermatol. Ther. 2012, 25, S44–S56. [Google Scholar] [CrossRef]
- Coates, C.J.; Nairn, J. Hemocyanin-derived phenoloxidase activity: A contributing factor to hyperpigmentation in Nephrops norvegicus. Food Chem. 2013, 140, 361–369. [Google Scholar] [CrossRef]
- Parrish, J.A.; Fitzpatrick, T.B.; Shea, C.; Pathak, M.A. Photochemotherapy of vitiligo: Use of orally administered psoralens and a high-intensity long-wave ultraviolet light system. Arch. Dermatol. 1976, 112, 1531–1534. [Google Scholar] [CrossRef]
- Kao, C.-H.; Yu, H.-S. Comparison of the effects of 8-methoxypsoralen (8-MOP) plus UVA (PUVA) on human melanocytes in vitiligo vulgaris and in vitro. J. Investig. Dermatol. 1992, 98, 734–740. [Google Scholar]
- Lerner, A.B.; Denton, C.R.; Fitzpatrick, T.B. Clinical and experimental studies with 8-methoxypsoralen in vitiligo. J. Investig. Dermatol. 1953, 20, 299–314. [Google Scholar]
- Nylander, K.; Bourdon, J.C.; Bray, S.E.; Gibbs, N.K.; Kay, R.; Hart, I.; Hall, P.A. Transcriptional activation of tyrosinase and TRP-I by p53 links UV irradiation to the protective tanning response. J. Pathol. 2000, 190, 39–46. [Google Scholar] [CrossRef]
- Galibert, M.D.; Carreira, S.; Goding, C.R. The Usf-1 transcription factor is a novel target for the stress-responsive p38 kinase and mediates UV-induced tyrosinase expression. EMBO J. 2001, 20, 5022–5031. [Google Scholar] [CrossRef]
- Deeni, Y.Y.; Ibbotson, S.H.; Woods, J.A.; Wolf, C.R.; Smith, G. Cytochrome P450 CYP1B1 interacts with 8-methoxypsoralen (8-MOP) and influences psoralen-ultraviolet A (PUVA) sensitivity. PLoS One 2013, 8, e75494. [Google Scholar]
- Koenigs, L.L.; Trager, W.F. Mechanism-based inactivation of cytochrome P450 2B1 by 8-methoxypsoralen and several other furanocoumarins. Biochemistry 1998, 37, 13184–13193. [Google Scholar] [CrossRef]
- Lei, T.C.; Virador, V.; Yasumoto, K.; Vieira, W.D.; Toyofuku, K.; Hearing, V.J. Stimulation of melanoblast pigmentation by 8-methoxypsoralen: The involvement of microphthalmia-associated transcription factor, the protein kinase A signal pathway, and proteasome-mediated degradation. J. Investig. Dermatol. 2002, 119, 1341–1349. [Google Scholar] [CrossRef]
- Hearing, V.J.; Tsukamoto, K. Enzymatic control of pigmentation in mammals. FASEB J. 1991, 5, 2902–2909. [Google Scholar]
- Pawelek, J.M. After dopachrome? Pigm. Cell Res. 1991, 4, 53–62. [Google Scholar] [CrossRef]
- Chen, Y.-S.; Lee, S.-M.; Lin, Y.-J.; Chiang, S.-H.; Lin, C.-C. Effects of Danshensu and Salvianolic Acid B from Salvia miltiorrhiza Bunge (Lamiaceae) on cell proliferation and collagen and melanin production. Molecules 2014, 19, 2029–2041. [Google Scholar] [CrossRef]
- Aroca, P.; Garcia-Borron, J.C.; Solano, F.; Lozano, J.A. Regulation of mammalian melanogenesis I: Partial purification and characterization of a dopachrome converting factor: Dopachrome tautomerase. BBA-Gen. Subj. 1990, 1035, 266–275. [Google Scholar] [CrossRef]
- Alaluf, S.; Atkins, D.; Barrett, K.; Blount, M.; Carter, N.; Heath, A. Ethnic variation in melanin content and composition in photoexposed and photoprotected human skin. Pigm. Cell Res. 2002, 15, 112–118. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef]
- Iwai, K.; Kishimoto, N.; Kakino, Y.; Mochida, K.; Fujita, T. In vitro antioxidative effects and tyrosinase inhibitory activities of seven hydroxycinnamoyl derivatives in green coffee beans. J. Agric. Food Chem. 2004, 52, 4893–4898. [Google Scholar] [CrossRef]
- Kong, J.-N.; Lee, H.-J.; Jo, D.-H.; Kong, K.-H. Characterization of human tyrosinase ectodomain expressed in Escherichia coli. Protein Peptide Lett. 2010, 17, 1026–1030. [Google Scholar] [CrossRef]
- Murata, M.; Sugiura, M.; Sonokawa, Y.; Shimamura, T.; Homma, S. Properties of chlorogenic acid quinone: Relationship between browning and the formation of hydrogen peroxide from a quinone solution. Biosci. Biotechnol. Biochem. 2002, 66, 2525–2530. [Google Scholar] [CrossRef]
- Munoz, J.; Garcia-Molina, F.; Varon, R.; Rodriguez-Lopez, J.; Garcia-Ruiz, P.; Garcia-Canovas, F.; Tudela, J. Kinetic characterization of the oxidation of chlorogenic acid by polyphenol oxidase and peroxidase. Characteristics of the o-quinone. J. Agric. Food Chem. 2007, 55, 920–928. [Google Scholar] [CrossRef]
- Ryu, H.W.; Jeong, S.H.; Curtis-Long, M.J.; Jung, S.; Lee, J.W.; Wo, H.S.; Cho, J.K.; Park, K.H. Inhibition effects of mangosenone F from garcinia mangostana on melanin formation in B16F10 cells. J. Agric. Food Chem. 2012, 60, 8372–8378. [Google Scholar] [CrossRef]
- Shirasugi, I.; Sakakibara, Y.; Yamasaki, M.; Nishiyama, K.; Matsui, T.; Liu, M.-C.; Suiko, M. Novel screening method for potential skin-whitening compounds by a luciferase reporter assay. Biosci. Biotechnol. Biochem. 2010, 74, 2253–2258. [Google Scholar] [CrossRef]
- Kuijpers, T.F.; Narváez-Cuenca, C.-E.; Vincken, J.-P.; Verloop, A.J.; van Berkel, W.J.; Gruppen, H. Inhibition of enzymatic browning of chlorogenic acid by sulfur-containing compounds. J. Agric. Food Chem. 2012, 60, 3507–3514. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds chlorogenic acid are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, H.-R.; Habasi, M.; Xie, L.-Z.; Aisa, H.A. Effect of Chlorogenic Acid on Melanogenesis of B16 Melanoma Cells. Molecules 2014, 19, 12940-12948. https://doi.org/10.3390/molecules190912940
Li H-R, Habasi M, Xie L-Z, Aisa HA. Effect of Chlorogenic Acid on Melanogenesis of B16 Melanoma Cells. Molecules. 2014; 19(9):12940-12948. https://doi.org/10.3390/molecules190912940
Chicago/Turabian StyleLi, Hao-Rong, Maidina Habasi, Lian-Zhen Xie, and Haji Akber Aisa. 2014. "Effect of Chlorogenic Acid on Melanogenesis of B16 Melanoma Cells" Molecules 19, no. 9: 12940-12948. https://doi.org/10.3390/molecules190912940