Peptides Derived from Rhopilema esculentum Hydrolysate Exhibit Angiotensin Converting Enzyme (ACE) Inhibitory and Antioxidant Abilities

Abstract
:1. Introduction
2. Results and Discussion
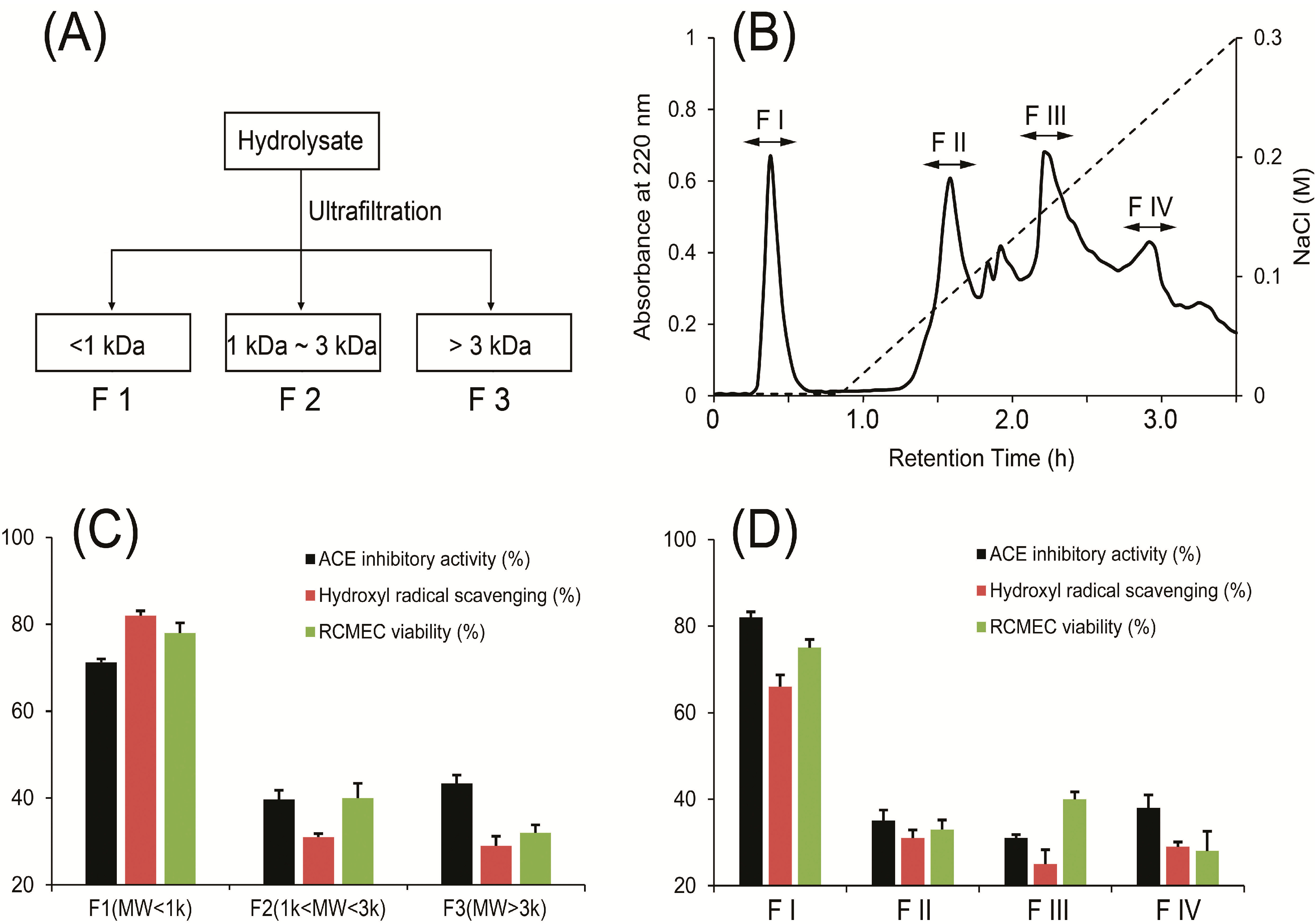
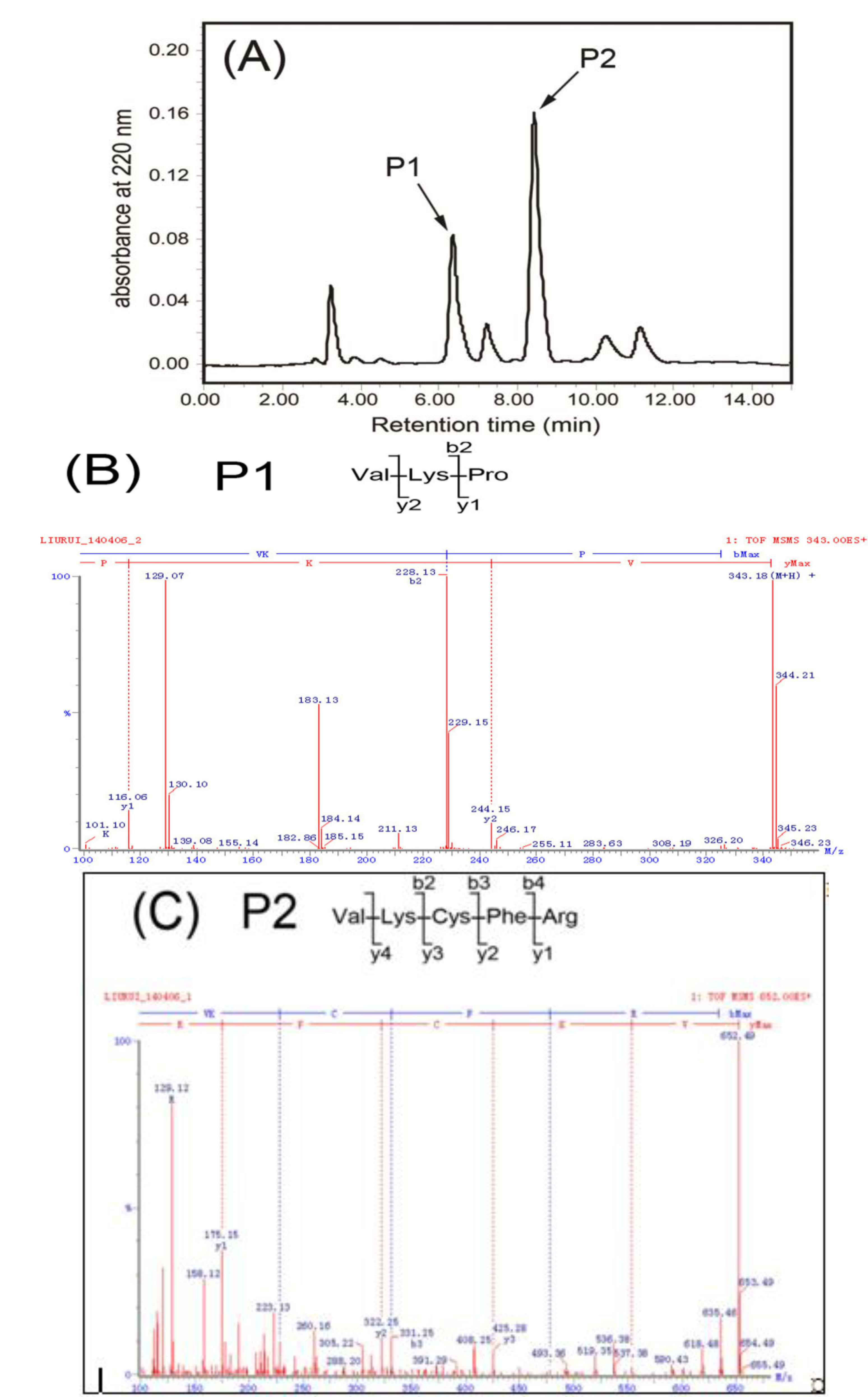
2.1. Isolation, Purification and Characterization of Peptides


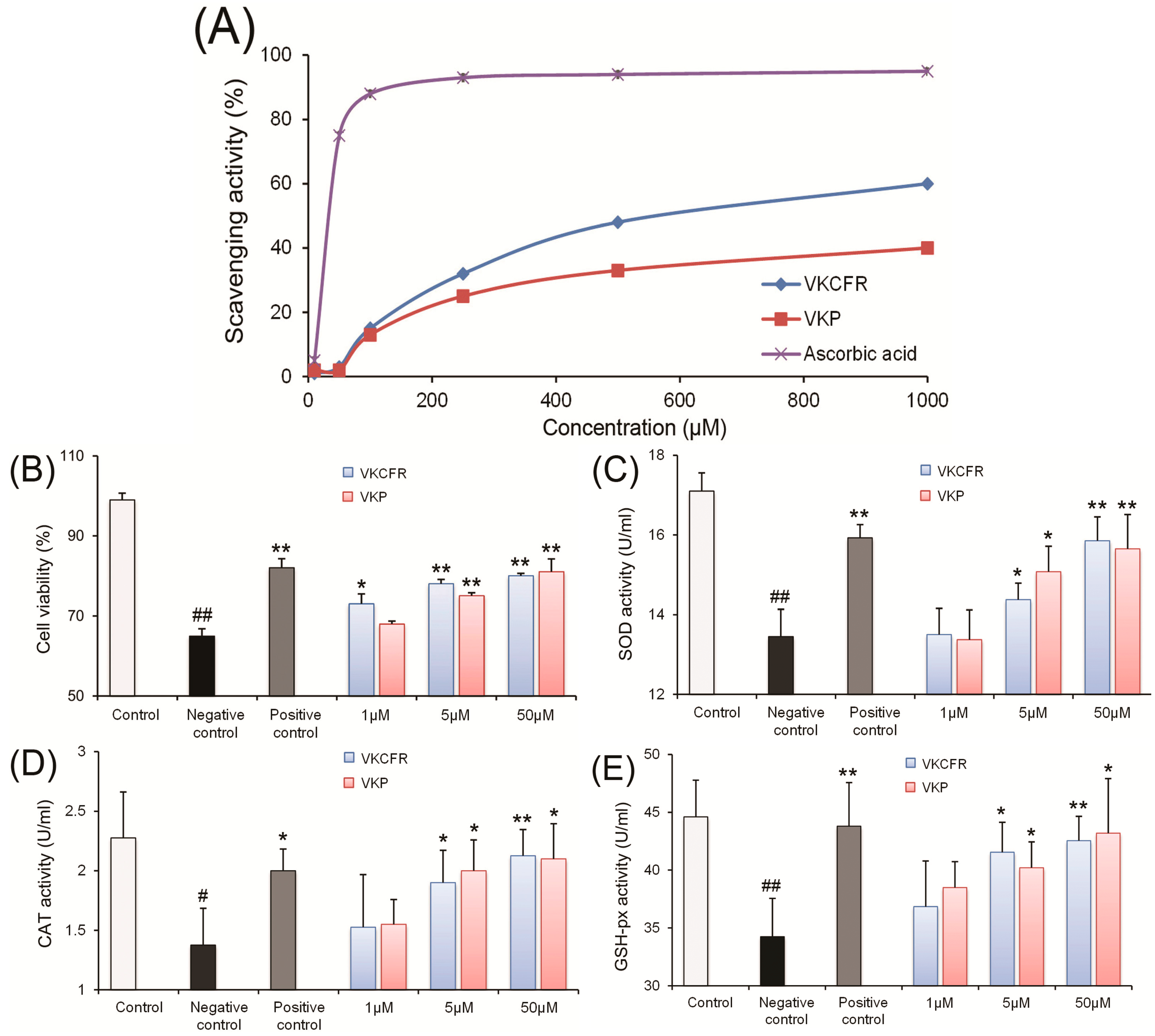
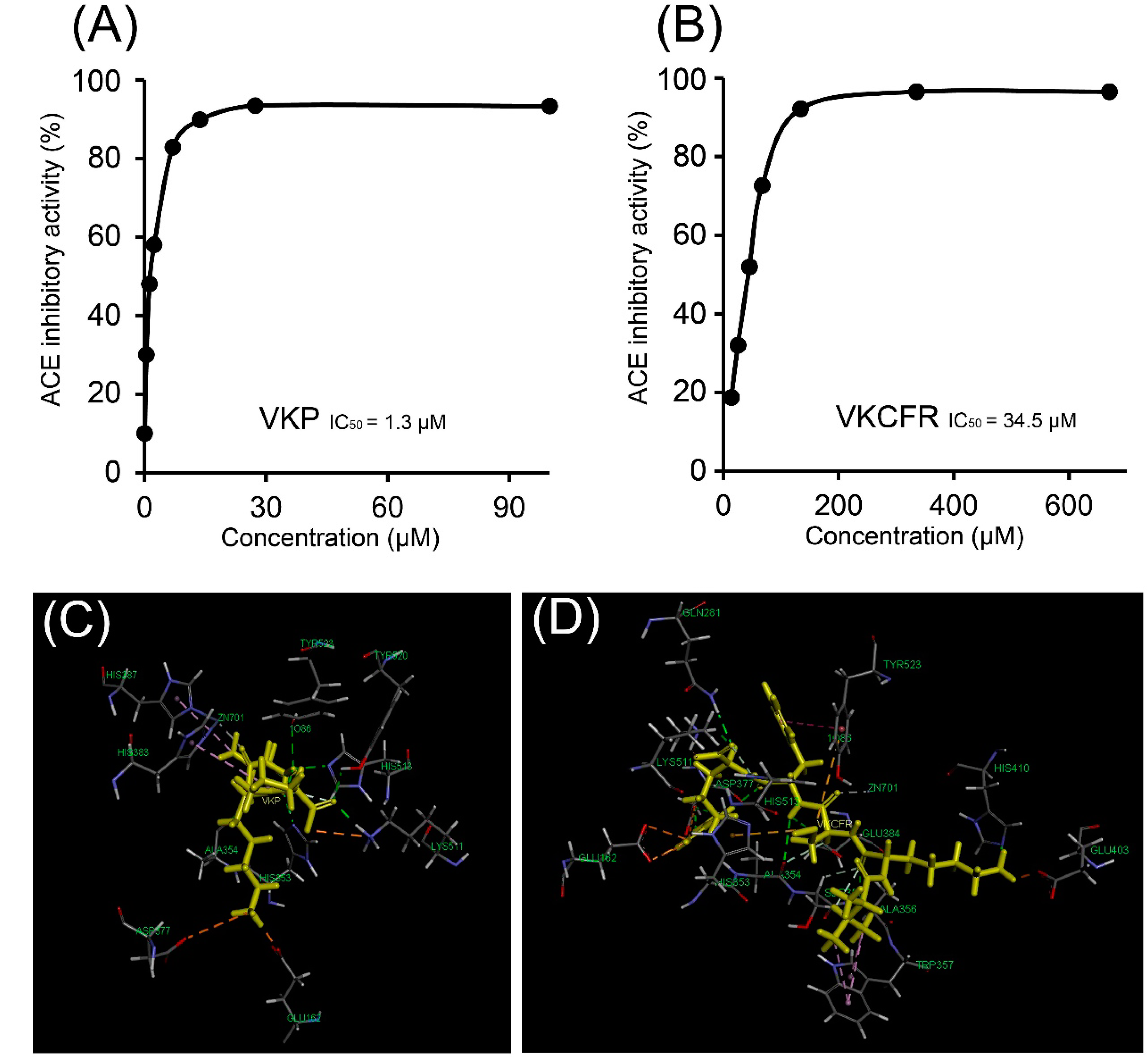
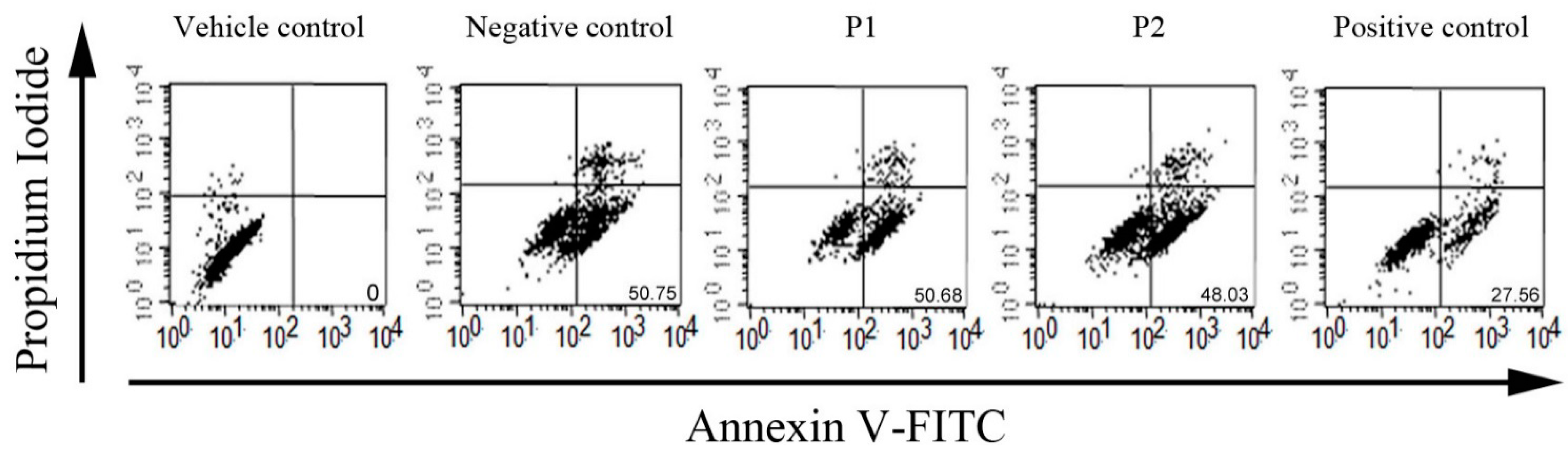
2.2. Antioxidant and ACE Inhibitory Activities of Purified Peptides


2.3. Molecular Docking of ACE and Peptides

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | Van der Waals Energy | Electrostatic Energy | Potential Energy | Bond Energy | CHARMm Energy |
|---|---|---|---|---|---|
| VKP | −8.347 | −52.149 | −26.028 | 1.296 | −26.028 |
| VKCFR | −17.823 | −134.961 | −132.409 | 1.988 | −132.409 |
3. Experimental Section
3.1. Materials
3.2. Preparation of Hydrolysate
3.3. Assay for ACE Inhibitory Activity

3.4. Determination of Antioxidant Activities
3.4.1. Hydroxyl Radical Scavenging Assay
3.4.2. Cell Culture and Cytotoxicity Determination
3.4.3. Determination of Antioxidant Enzyme Activity
3.5. Purification of Bioactive Peptides
3.5.1. Ultrafiltration
3.5.2. Ion-Exchange Chromatography
3.5.3. RP-HPLC
3.6. Characterization of Purified Peptides
3.7. Peptide Synthesis
3.8. Molecular Docking Analysis
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Abbreviation
| RCMEC | rat cerebral microvascular endothelial cell |
| ACE | angiotensin-I-converting enzyme |
| SOD | superoxide dismutase |
| CAT | catalase |
| GSH-px | glutathione peroxidase |
| BHA | butylated hydroxyanisole |
| BHT | butylated hydroxytoluene |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| HHL | hippuryl-histidyl-leucine |
| HA | hippuric acid |
Appendix
Detection of Apoptotic Cells by Flow Cytometry

Conflicts of Interest
References
- Zhuang, Y.; Hou, H.; Zhao, X.; Zhang, Z.; Li, B. Effects of collagen and collagen hydrolysate from jellyfish (Rhopilema esculentum) on mice skin photoaging induced by UV irradiation. J. Food Sci. 2009, 74, H183–H188. [Google Scholar]
- Yu, H.H.; Liu, X.G.; Xing, R.E.; Liu, S.; Guo, Z.Y.; Wang, P.B.; Li, C.P.; Li, P.C. In vitro determination of antioxidant activity of proteins from jellyfish Rhopilema esculentum. Food Chem. 2006, 95, 123–130. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, M.; Zhang, C.; Liu, C. Angiotensin converting enzyme (ACE) inhibitory, antihypertensive and antihyperlipidaemic activities of protein hydrolysates from Rhopilema esculentum. Food Chem. 2012, 134, 2134–2140. [Google Scholar]
- Xiao, L.; Liu, S.; He, Q.; Wang, Q.; Ye, X.; Liu, G.; Nie, F.; Zhao, J.; Zhang, L. The Acute Toxicity and Hematological Characterization of the Effects of Tentacle-Only Extract from the Jellyfish Cyanea capillata. Mar. Drugs 2011, 9, 526–534. [Google Scholar]
- Murray, B.A.; FitzGerald, R.J. Angiotensin converting enzyme inhibitory peptides derived from food proteins: Biochemistry, bioactivity and production. Curr. Pharm. Des. 2007, 13, 773–791. [Google Scholar] [CrossRef]
- Liu, R.; Wang, M.; Duan, J.A.; Guo, J.M.; Tang, Y.P. Purification and identification of three novel antioxidant peptides from Cornu Bubali (water buffalo horn). Peptides 2010, 31, 786–793. [Google Scholar]
- Shahidi, F.; Han, X.Q.; Synowiecki, J. Production and characteristics of protein hydrolysates from capelin (Mallotus villosus). Food Chem. 1995, 53, 285–293. [Google Scholar]
- Salampessy, J.; Phillips, M.; Seneweera, S.; Kailasapathy, K. Release of antimicrobial peptides through bromelain hydrolysis of leatherjacket (Meuchenia sp.) insoluble proteins. Food Chem. 2010, 120, 556–560. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Antioxidant and angiotensin converting enzyme-inhibitory properties of a flaxseed protein-derived high Fischer ratio peptide mixture. J. Agric. Food Chem. 2010, 58, 4762–4768. [Google Scholar]
- Vaštag, Z.; Popović, L.; Popović, S.; Peričin-Starčević, I.; Krimer-Malešević, V. In vitro study on digestion of pumpkin oil cake protein hydrolysate: Evaluation of impact on bioactive properties. Int. J. Food Sci. Nutr. 2013, 64, 452–460. [Google Scholar] [CrossRef]
- Ngo, D.H.; Ryu, B.; Kim, S.K. Active peptides from skate (Okamejei kenojei) skin gelatin diminish angiotensin-I converting enzyme activity and intracellular free radical-mediated oxidation. Food Chem. 2014, 143, 246–255. [Google Scholar]
- Lun, H.H.; Lan, C.X.; Yun, S.C.; Zhong, Z.Y.; Cheng, Z.B. Analysis of novel angiotensin-I-converting enzyme inhibitory peptides from protease-hydrolyzed marine shrimp Acetes chinensis. J. Pept. Sci. 2006, 12, 726–733. [Google Scholar]
- Tsai, J.S.; Chen, J.L.; Pan, B.S. ACE-inhibitory peptides identified from the muscle protein hydrolysate of hard clam (Meretrix lusoria). Process Biochem. 2008, 43, 743–747. [Google Scholar] [CrossRef]
- Harrison, D.G.; Gongora, M.C. Oxidative stress and hypertension. Med. Clin. N. Am. 2009, 93, 621–635. [Google Scholar]
- Muhammad, S.; Bierhaus, A.; Schwaninger, M. Reactive oxygen species in diabetes-induced vascular damage, stroke, and Alzheimer’s disease. J. Alzheimers Dis. 2009, 16, 775–785. [Google Scholar]
- Rao, K.S. Free radical induced oxidative damage to DNA: Relation to brain aging and neurological disorders. Indian J. Biochem. Biophys. 2009, 46, 9–15. [Google Scholar]
- Kuhlen, J.L.; Forcucci, J. Angiotensin-converting enzyme inhibitor-induced unilateral tongue angioedema. Am. J. Med. Sci. 2012, 344, 416–417. [Google Scholar]
- Saleh, A.S.; Zhang, Q.; Shen, Q. Recent research in antihypertensive activity of food protein-derived hydrolyzates and peptides. Crit. Rev. Food Sci. Nutr. 2014. [Google Scholar] [CrossRef]
- Amado, I.R.; Vázquez, J.A.; González, P.; Esteban-Fernández, D.; Carrera, M.; Piñeiro, C. Identification of the major ACE-inhibitory peptides produced by enzymatic hydrolysis of a protein concentrate from cuttlefish wastewater. Mar. Drugs 2014, 12, 1390–1405. [Google Scholar]
- Pan, D.D.; Cao, J.X.; Guo, H.Q.; Zhao, B. Studies on purification and the molecular mechanism of a novel ACE inhibitory peptide from whey protein hydrolysate. Food Chem. 2012, 130, 121–126. [Google Scholar] [CrossRef]
- Vercruysse, L.; van Camp, J.; Smagghe, G. ACE inhibitory peptides derived from enzymatic hydrolysates of animal muscle protein: A review. J. Agric. Food Chem. 2005, 53, 8106–8115. [Google Scholar]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of Angiotensin I-converting enzyme inhibitory peptides: Quantitative structure-activity relationship study of di- and tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar]
- Tavares, T.; Sevilla, M.Á.; Montero, M.J.; Carrón, R.; Malcata, F.X. Acute effect of whey peptides upon blood pressure of hypertensive rats, and relationship with their angiotensin-converting enzyme inhibitory activity. Mol. Nutr. Food Res. 2012, 56, 316–324. [Google Scholar]
- Hernández-Ledesma, B.; del Mar Contreras, M.; Recio, I. Antihypertensive peptides: Production, bioavailability and incorporation into foods. Adv. Colloid Interface Sci. 2011, 165, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.P.; Hu, J.E.; Cui, J.Z.; Bai, X.F.; Du, Y.G.; Miyaguchi, Y.; Lin, B.C. Purification and identification of a ACE inhibitory peptide from oyster proteins hydrolysate and the antihypertensive effect of hydrolysate in spontaneously hypertensive rats. Food Chem. 2008, 111, 302–308. [Google Scholar] [CrossRef]
- Balti, R.; Nedjar-Arroume, N.; Adjé, E.Y.; Guillochon, D.; Nasri, M. Analysis of novel angiotensin I-converting enzyme inhibitory peptides from enzymatic hydrolysates of cuttlefish (Sepia officinalis) muscle proteins. J. Agric. Food Chem. 2010, 58, 3840–3846. [Google Scholar] [CrossRef]
- Sundaram, A.; Siew Keah, L.; Sirajudeen, K.N.; Singh, H.J. Upregulation of catalase and downregulation of glutathione peroxidase activity in the kidney precede the development of hypertension in pre-hypertensive SHR. Hypertens. Res. 2013, 36, 213–218. [Google Scholar]
- Rodrigo, R.; González, J.; Paoletto, F. The role of oxidative stress in the pathophysiology of hypertension. Hypertens. Res. 2011, 34, 431–440. [Google Scholar]
- Jimsheena, V.K.; Gowda, L.R. Arachin derived peptides as selective angiotensin I-converting enzyme (ACE) inhibitors: Structure-activity relationship. Peptides 2010, 31, 1165–1176. [Google Scholar]
- Gentilucci, L.; Tolomelli, A.; de Marco, R.; Artali, R. Molecular docking of opiates and opioid peptides, a tool for the design of selective agonists and antagonists, and for the investigation of atypical ligand-receptor interactions. Curr. Med. Chem. 2012, 19, 1587–1601. [Google Scholar] [CrossRef]
- Qian, Z.J.; Jung, W.K.; Kim, S.K. Free radical scavenging activity of a novel antioxidative peptide purified from hydrolysate of bullfrog skin, Rana catesbeiana Shaw. Bioresource Technol. 2008, 99, 1690–1698. [Google Scholar]
- Arechabala, B.; Coiffard, C.; Rivalland, P.; Coiffard, L.J.; de Roeck-Holtzhauer, Y. Comparison of cytotoxicity of various surfactants tested on normal human fibroblast cultures using the neutral red test, MTT assay and LDH release. J. Appl. Toxicol. 1999, 19, 163–165. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar]
- Crystal Structure of Human Angiotensin Converting Enzyme in complex with Lisinopril. Available online: http://www.rcsb.org/pdb/explore.do?pdbId=1O86 (accessed on 4 April 2014).
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, J.; Li, Q.; Li, J.; Zhou, B. Peptides Derived from Rhopilema esculentum Hydrolysate Exhibit Angiotensin Converting Enzyme (ACE) Inhibitory and Antioxidant Abilities. Molecules 2014, 19, 13587-13602. https://doi.org/10.3390/molecules190913587
Li J, Li Q, Li J, Zhou B. Peptides Derived from Rhopilema esculentum Hydrolysate Exhibit Angiotensin Converting Enzyme (ACE) Inhibitory and Antioxidant Abilities. Molecules. 2014; 19(9):13587-13602. https://doi.org/10.3390/molecules190913587
Chicago/Turabian StyleLi, Jun, Qian Li, Jingyun Li, and Bei Zhou. 2014. "Peptides Derived from Rhopilema esculentum Hydrolysate Exhibit Angiotensin Converting Enzyme (ACE) Inhibitory and Antioxidant Abilities" Molecules 19, no. 9: 13587-13602. https://doi.org/10.3390/molecules190913587