Tocotrienol-Rich Fraction, [6]-Gingerol and Epigallocatechin Gallate Inhibit Proliferation and Induce Apoptosis of Glioma Cancer Cells

Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Bioactives on the Viability of Glioma Cells

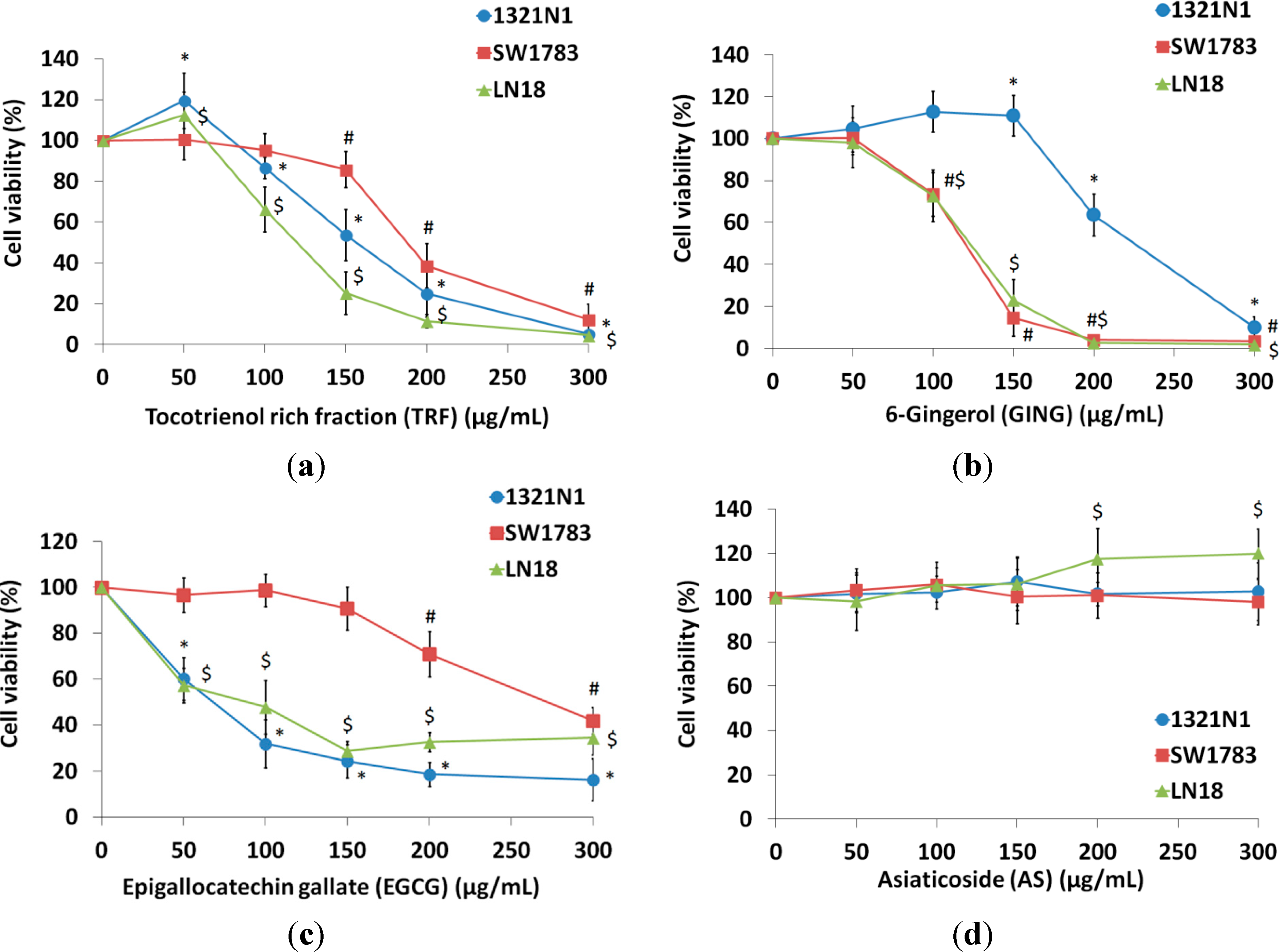{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Compound | IC50 Value (µg/mL) | Viability (% Cells) a |
|---|---|---|---|
| Grade II 1321N1 | Tocotrienol rich fraction 70 (TRF) | 171.5 ± 11.43 | 5.1 ± 1.1 |
| Epigallocatechin gallate (EGCG) | 82.0 ± 10.31 | 16.3 ± 9.2 | |
| [6]-gingerol (GING) | 243.0 ± 11.6 | 10.0 ± 5.0 | |
| Asiaticoside (AS) | n/a | 102.8 ± 11.1 | |
| Grade III SW1783 | Tocotrienol rich fraction 70 (TRF) | 202.0 ± 6.02 | 12.3 ± 7.5 |
| Epigallocatechin gallate (EGCG) | 300.0 ± 9.10 | 41.9 ± 5.7 | |
| [6]-gingerol (GING) | 132 ± 4.51 | 3.5 ± 2.0 | |
| Asiaticoside (AS) | n/a | 98.1 ± 10.3 | |
| Grade IV LN18 | Tocotrienol rich fraction 70 (TRF) | 142.0 ± 5.03 | 4.5 ± 1.9 |
| Epigallocatechin gallate (EGCG) | 134.0 ± 11.36 | 34.4 ± 7.4 | |
| [6]-gingerol (GING) | 132.5 ± 10.11 | 1.8 ± 0.9 | |
| Asiaticoside (AS) | n/a | 120.1 ± 11.2 |

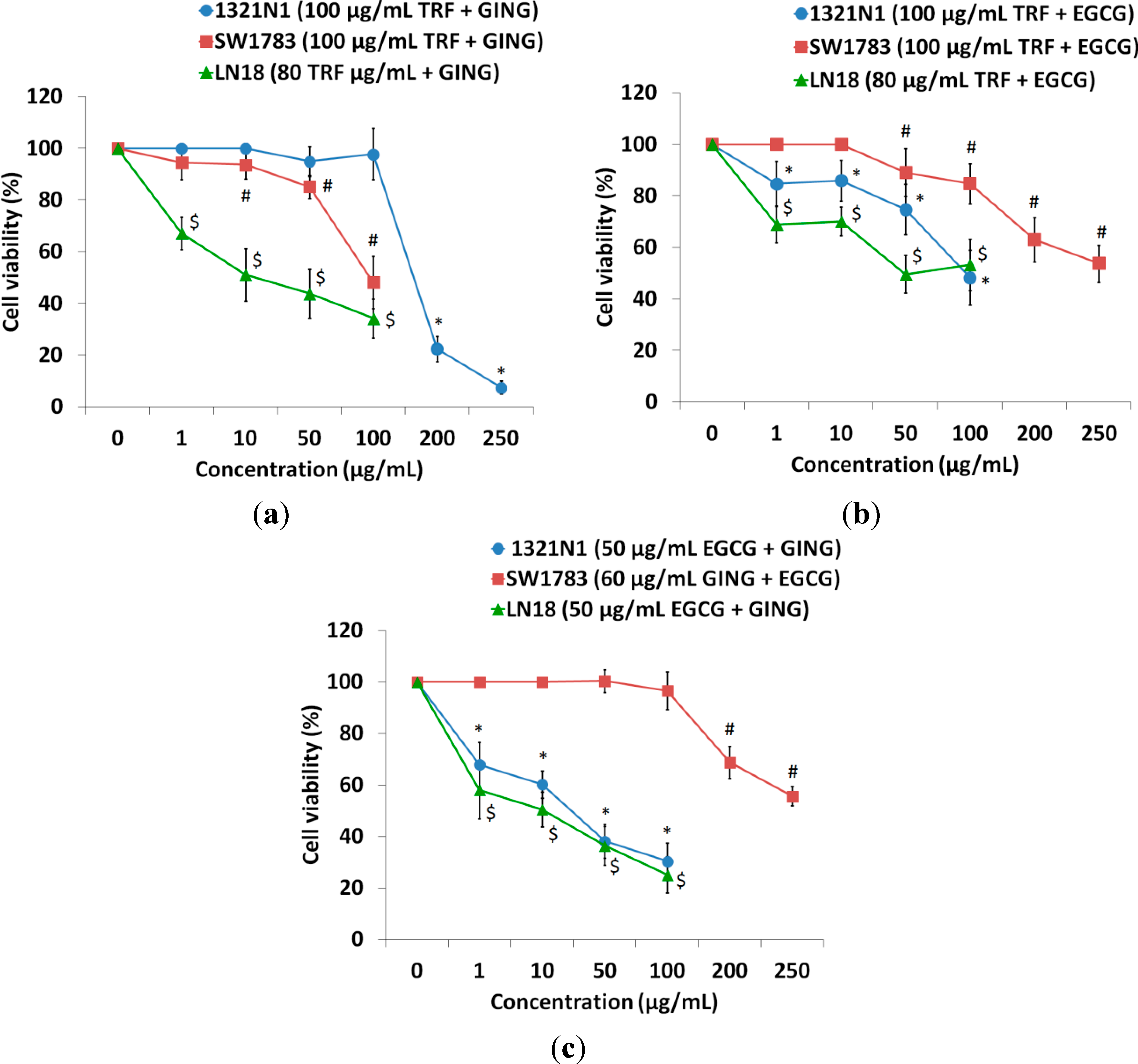
2.2. Isobologram Analysis of the Interaction between Bioactive Compounds

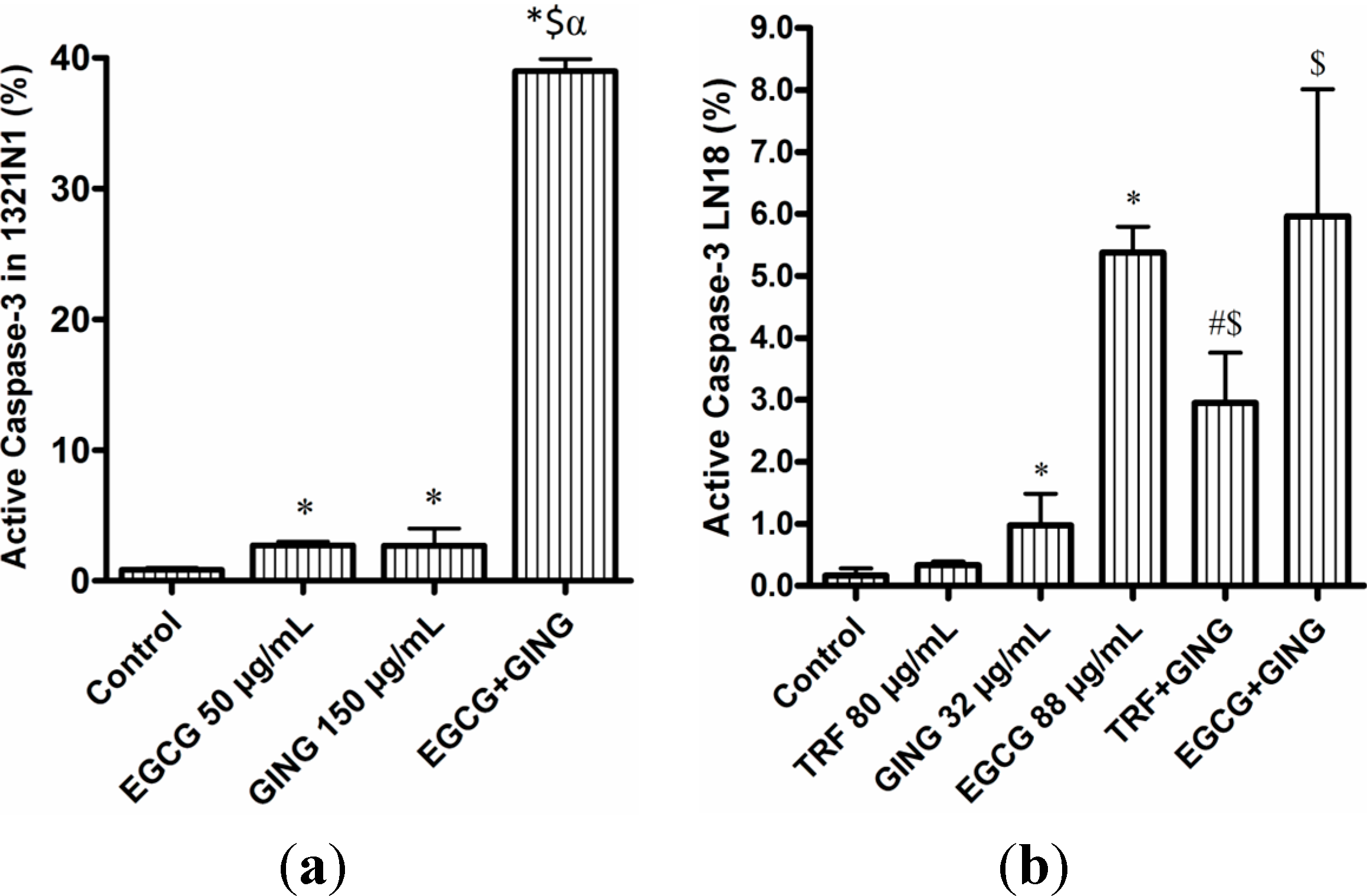
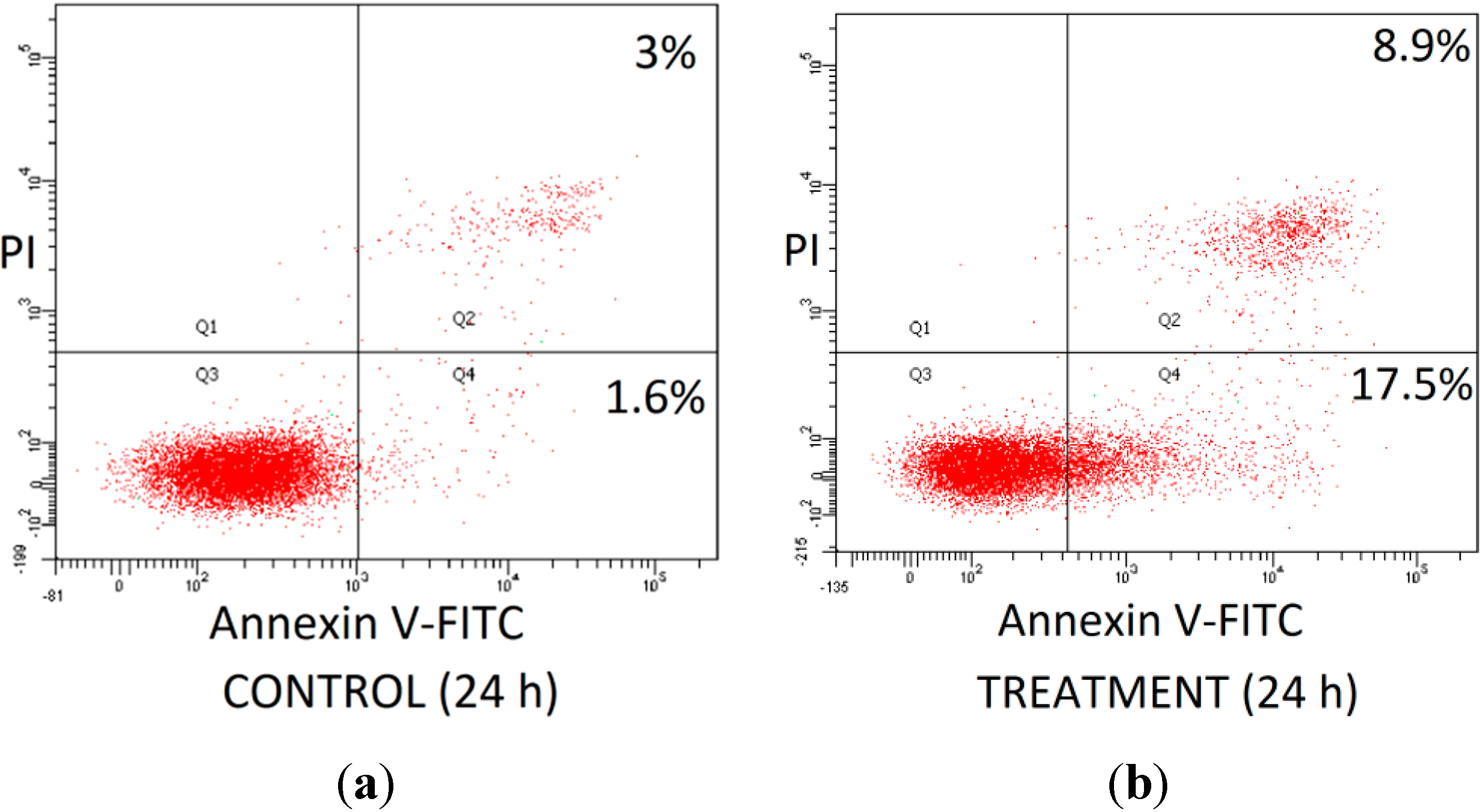
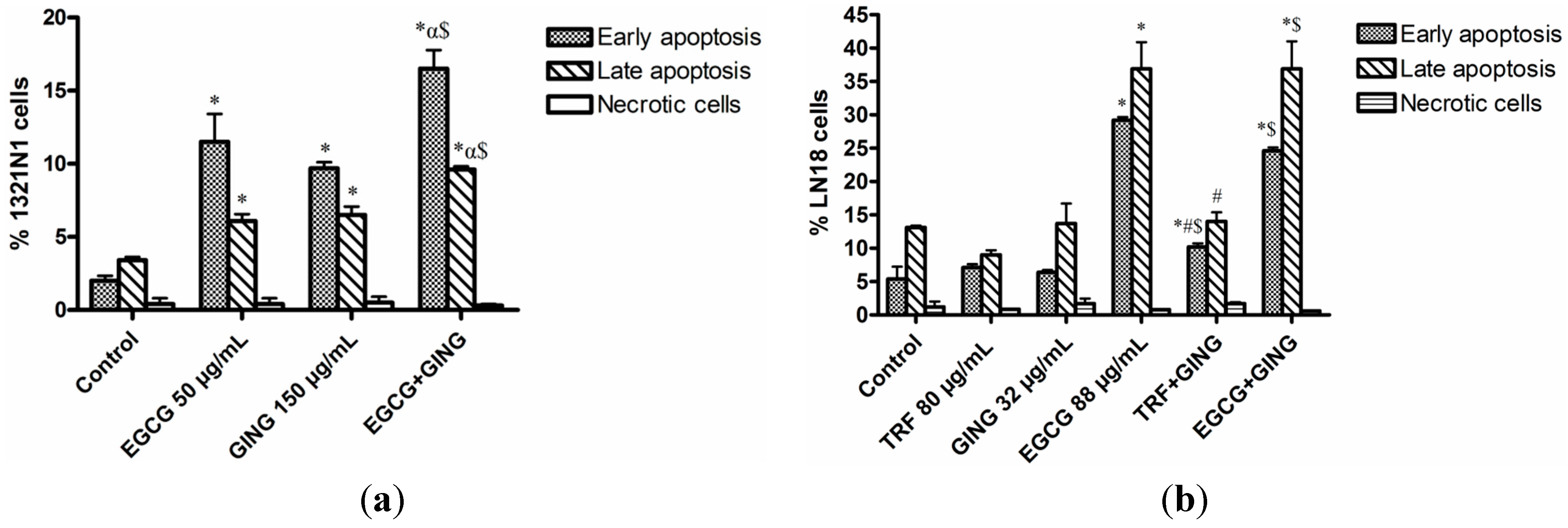
2.3. Effect of Combined Bioactives on Apoptosis
| Type of Cell Line | TRF:GING | IC50 a (µg/mL) | TRF b (µg/mL) | GING b (µg/mL) | Combination Index c (CI) |
| 1321N1 | 5:8 | 160 ± 4.36 | 171.5 ± 11.43 | 243 ± 11.60 | 1.24 |
| SW1783 | 10:11 | 110 ± 8.89 | 202 ± 6.02 | 132 ± 4.51 | 1.29 |
| LN18 | 5:2 | 32 ± 10.21 | 142 ± 5.03 | 132.5 ± 10.11 | 0.80 |
| Type of Cell Line | TRF:EGCG | IC50 a (µg/mL) | TRF b (µg/mL) | EGCG b (µg/mL) | Combination Index c (CI) |
| 1321N1 | 1:1 | 100 ± 9.5 | 171.5 ± 11.43 | 82 ± 10.31 | 1.80 |
| SW1783 | 10:27 | 270 ± 4.16 | 202 ± 6.02 | 300 ± 9.10 | 1.39 |
| LN18 | 10:11 | 88 ± 11.14 | 142 ± 5.03 | 134 ± 11.36 | 1.22 |
| Type of Cell Line | EGCG:GING | IC50 a (µg/mL) | EGCG b (µg/mL) | GING b (µg/mL) | Combination Index c (CI) |
| 1321N1 | 5:4 | 40 ± 8.62 | 82 ± 10.31 | 243 ± 11.60 | 0.77 |
| SW1783 | 9:2 | 60 ± 5.6 | 300 ± 9.10 | 132 ± 4.51 | 1.35 |
| LN18 | 2:1 | 24 ± 2.65 | 134 ± 11.36 | 132.5 ± 10.11 | 0.55 |



3. Experimental Section
3.1. Reagents and Chemicals
3.2. Cell Line and Culture Condition
3.3. Treatments with Natural Compounds
3.4. Determination of Cell Viability
3.5. Active Caspase-3 Apoptosis Assay
3.6. Annexin V-Propidium Iodide Staining Apoptosis Assay
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Furnari, F.B.; Fenton, T.; Bachoo, R.M.; Mukasa, A.; Stommel, J.M.; Stegh, A.; Hahn, W.C.; Ligon, K.L.; Louis, D.N.; Brennan, C.; et al. Malignant astrocytic glioma: Genetics, biology, and paths to treatment. Gene. Dev. 2007, 21, 2683–2710. [Google Scholar] [CrossRef]
- Wagner, H.; Ulrich-Merzenich, G. Synergy research: Approaching a new generation of phytopharmaceuticals. Phytomedicine 2009, 16, 97–110. [Google Scholar] [CrossRef]
- Straetemans, R.; O’Brien, T.; Wouters, L.; van Dun, J.; Janicot, M.; Bijnens, L.; Burzykowski, T.; Aerts, M. Design and analysis of drug combination experiments. Biom. J. 2005, 47, 299–308. [Google Scholar] [CrossRef]
- Imming, P.; Sinning, C.; Meyer, A. Drugs, their targets and the nature and number of drug targets. Nat. Rev. Drug Discov. 2006, 5, 821–834. [Google Scholar] [CrossRef]
- Ulrich-Merzenich, G.; Panek, D.; Zeitler, H.; Vetter, H.; Wagner, H. Drug development from natural products: Exploiting synergistic effects. Indian J. Exp. Biol. 2010, 48, 208–219. [Google Scholar]
- Ong, F.B.; wan Ngah, W.Z.; Marzuki, A.; Kadir, K.A.; Abdullah, N.; Top, A.G.M.; Shamaan, N.A. Effect of vitamin E supplementation on the immune response during chemically induced hepatocarcinogenesis in the rat. J. Clin. Biochem. Nutr. 1994, 17, 161–169. [Google Scholar] [CrossRef]
- Nigam, N.; George, J.; Srivastava, S.; Roy, P.; Bhui, K.; Singh, M.; Shukla, Y. Induction of apoptosis by [6]-gingerol associated with the modulation of p53 and involvement of mitochondrial signaling pathway in B[a]P-induced mouse skin tumorigenesis. Cancer Chemoth. Pharmacol. 2010, 65, 687–696. [Google Scholar] [CrossRef]
- Meeran, S.M.; Ahmed, A.; Tollefsbol, T.O. Epigenetic targets of bioactive dietary components for cancer prevention and therapy. Clin. Epigenet. 2010, 1, 101–116. [Google Scholar] [CrossRef]
- Bunpo, P.; Kataoka, K.; Arimochi, H.; Nakayama, H.; Kuwahara, T.; Bando, Y.; Izumi, K.; Vinitketkumnuen, U.; Ohnishi, Y. Inhibitory effects of Centella asiatica on azoxymethane-inducedaberrant crypt focus formation and carcinogenesis in the intestines of F344 rats. Food Chem. Toxicol. 2004, 42, 1987–1997. [Google Scholar] [CrossRef]
- Viola, V.; Pilolli, F.; Piroddi, M.; Pierpaoli, E.; Orlando, F.; Provinciali, M.; Betti, M.; Mazzini, F.; Galli, F. Why tocotrienols work better: Insights into the in vitro anti-cancer mechanism of vitamin E. Gene. Nutr. 2012, 7, 29–41. [Google Scholar] [CrossRef]
- Nigam, N.; Bhui, K.; Prasad, S.; George, J.; Shukla, Y. [6]-Gingerol induces reactive oxygen species regulated mitochondrial cell death pathway in human epidermoid carcinoma A431 cells. Chem. Biol. Interact. 2009, 181, 77–84. [Google Scholar] [CrossRef]
- Al-Saeedi, F.J.; Bitar, M.; Pariyani, S. Effect of asiaticoside on 99mTc-tetrofosmin and 99mTc-sestamibi uptake in MCF-7 cells. J. Nucl. Med. Technol. 2011, 39, 279–283. [Google Scholar] [CrossRef]
- Leaver, K.R.; Allbutt, H.N.; Creber, N.J.; Kassiou, M.; Henderson, J.M. Oral pre-treatment with epigallocatechin gallate in 6-OHDA lesioned rats produces subtle symptomatic relief but not neuroprotection. Brain Res. Bull. 2009, 80, 397–402. [Google Scholar] [CrossRef]
- Hwang, J.T.; Ha, J.; Park, I.J.; Lee, S.K.; Baik, H.W.; Kim, Y.M.; Park, O.J. Apoptotic effect of EGCG in HT-29 colon cancer cells via AMPK signal pathway. Cancer Lett. 2007, 247, 115–121. [Google Scholar] [CrossRef]
- Park, Y.J.; Wen, J.; Bang, S.; Park, S.W.; Song, S.Y. [6]-Gingerol induces cell cycle arrest and cell death of mutant p53-expressing pancreatic cancer cells. Yonsei Med. J. 2006, 47, 688–697. [Google Scholar] [CrossRef]
- Park, S.K.; Sanders, B.G.; Kline, K. Tocotrienols induce apoptosis in breast cancer cell lines via an endoplasmic reticulum stress-dependent increase in extrinsic death receptor signaling. Breast Cancer Res. Treat. 2010, 124, 361–375. [Google Scholar] [CrossRef]
- Kunysz, E.A.; Michel, A.D.; Whiting, R.L.; Woods, K. The human astrocytoma cell line 1321 N1 contains M2-glandular type muscarinic receptors linked to phosphoinositide turnover. Br. J. Pharmacol. 1989, 96, 271–278. [Google Scholar] [CrossRef]
- Meléndez, B.; García-Claver, A.; Ruano, Y.; Campos-Martín, Y.; de Lope, Á.R.; Pérez-Magán, E.; Mur, P.; Torres, S.; Lorente, M.; Velasco, G.; et al. Copy number alterations in glioma cell lines. In Glioma—Exploring Its Biology and Practical Relevance; Ghosh, D.A., Ed.; InTech: Rijeka, Croatia, 2011; pp. 429–448. [Google Scholar]
- Garcia-Claver, A.; Lorente, M.; Mur, P.; Campos-Martin, Y.; Mollejo, M.; Velasco, G.; Melendez, B. Gene expression changes associated with erlotinib response in glioma cell lines. Eur. J. Cancer 2013, 49, 1641–1653. [Google Scholar] [CrossRef]
- Diserens, A.C.; de Tribolet, N.; Martin-Achard, A.; Gaide, A.C.; Schnegg, J.F.; Carrel, S. Characterization of an established human malignant glioma cell line: LN-18. Acta Neuropathol. 1981, 53, 21–28. [Google Scholar] [CrossRef]
- Ishii, N.; Maier, D.; Merlo, A.; Tada, M.; Sawamura, Y.; Diserens, A.C.; van Meir, E.G. Frequent co-alterations of TP53, p16/CDKN2A, p14ARF, PTEN tumor suppressor genes in human glioma cell lines. Brain Pathol. 1999, 9, 469–479. [Google Scholar] [CrossRef]
- Kok, T.M.; Breda, S.G.; Briede, J.J. Genomics-based identification of molecular mechanisms behindthe cancer preventive action of phytochemicals: Potential and challenges. Curr. Pharm. Biotechnol. 2012, 13, 255–264. [Google Scholar] [CrossRef]
- Kannappan, R.; Gupta, S.C.; Kim, J.H.; Aggarwal, B.B. Tocotrienols fight cancer by targeting multiple cell signaling pathways. Gene. Nutr. 2012, 7, 43–52. [Google Scholar] [CrossRef]
- Weng-Yew, W.; Selvaduray, K.R.; Ming, C.H.; Nesaretnam, K. Suppression of tumor growth by palm tocotrienols via the attenuation of angiogenesis. Nutr. Cancer 2009, 61, 367–373. [Google Scholar] [CrossRef]
- Sen, C.K.; Khanna, S.; Roy, S. Tocotrienols in health and disease: The other half of the natural vitamin E family. Mol. Aspects Med. 2007, 28, 692–728. [Google Scholar] [CrossRef]
- Huang, W.Y.; Cai, Y.Z.; Zhang, Y. Natural phenolic compounds from medicinal herbs and dietary plants: Potential use for cancer prevention. Nutr. Cancer 2010, 62, 1–20. [Google Scholar] [CrossRef]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): mechanisms, perspectives and clinical applications. Biochem. Pharm. 2011, 82, 1807–1821. [Google Scholar] [CrossRef]
- Zhang, R.; Banik, N.L.; Ray, S.K. Combination of all-trans retinoic acid and interferon-gamma upregulated p27(kip1) and down regulated CDK2 to cause cell cycle arrest leading to differentiation and apoptosis in human glioblastoma LN18 (PTEN-proficient) and U87MG (PTEN-deficient) cells. Cancer Chemoth. Pharm. 2008, 62, 407–416. [Google Scholar] [CrossRef]
- Gupta, R.A.; Brockman, J.A.; Sarraf, P.; Willson, T.M.; DuBois, R.N. Target genes of peroxisome proliferator-activated receptor gamma in colorectal cancer cells. J. Biol. Chem. 2001, 276, 29681–29687. [Google Scholar]
- Hsieh, T.C.; Wu, J.M. Targeting CWR22Rv1 prostate cancer cell proliferation and gene expression by combinations of the phytochemicals EGCG, genistein and quercetin. Anticancer Res. 2009, 29, 4025–4032. [Google Scholar]
- Tan, S.W.; Ramasamy, R.; Abdullah, M.; Vidyadaran, S. Inhibitory effects of palm α-, γ- and δ-tocotrienol on lipopolysaccharide-induced nitric oxide production in BV2 microglia. Cell. Immunol. 2011, 271, 205–209. [Google Scholar] [CrossRef]
- Suganuma, M.; Okabe, S.; Oniyama, M.; Tada, Y.; Ito, H.; Fujiki, H. Wide distribution of [3H](−)-epigallocatechin gallate, a cancer preventive tea polyphenol, in mouse tissue. Carcinogenesis 1998, 19, 1771–1776. [Google Scholar] [CrossRef]
- Jiang, S.Z.; Wang, N.S.; Mi, S.Q. Plasma pharmacokinetics and tissue distribution of [6]-gingerol in rats. Biopharm. Drug Dispos. 2008, 29, 529–537. [Google Scholar] [CrossRef]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Vadhanam, M.V. Bioavailability of phytochemicals and its enhancement by drug delivery systems. Cancer Lett. 2013. [Google Scholar] [CrossRef]
- Smith, A.; Giunta, B.; Bickford, P.C.; Fountain, M.; Tan, J.; Shytle, R.D. Nanolipidic particles improve the bioavailability and alpha-secretase inducing ability of epigallocatechin-3-gallate (EGCG) for the treatment of Alzheimer’s disease. Int. J. Pharm. 2010, 389, 207–212. [Google Scholar] [CrossRef]
- Chou, T.C.; Talalay, P. Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv. Enzym. Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Zhao, L.; Wientjes, M.G.; Au, J.L. Evaluation of combination chemotherapy: Integration of nonlinear regression, curve shift, isobologram, and combination index analyses. Clin. Cancer Res. 2004, 10, 7994–8004. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rahman, A.A.; Makpol, S.; Jamal, R.; Harun, R.; Mokhtar, N.; Ngah, W.Z.W. Tocotrienol-Rich Fraction, [6]-Gingerol and Epigallocatechin Gallate Inhibit Proliferation and Induce Apoptosis of Glioma Cancer Cells. Molecules 2014, 19, 14528-14541. https://doi.org/10.3390/molecules190914528
Rahman AA, Makpol S, Jamal R, Harun R, Mokhtar N, Ngah WZW. Tocotrienol-Rich Fraction, [6]-Gingerol and Epigallocatechin Gallate Inhibit Proliferation and Induce Apoptosis of Glioma Cancer Cells. Molecules. 2014; 19(9):14528-14541. https://doi.org/10.3390/molecules190914528
Chicago/Turabian StyleRahman, Amirah Abdul, Suzana Makpol, Rahman Jamal, Roslan Harun, Norfilza Mokhtar, and Wan Zurinah Wan Ngah. 2014. "Tocotrienol-Rich Fraction, [6]-Gingerol and Epigallocatechin Gallate Inhibit Proliferation and Induce Apoptosis of Glioma Cancer Cells" Molecules 19, no. 9: 14528-14541. https://doi.org/10.3390/molecules190914528