Neuroprotective Effects of Daphnetin against NMDA Receptor-Mediated Excitotoxicity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
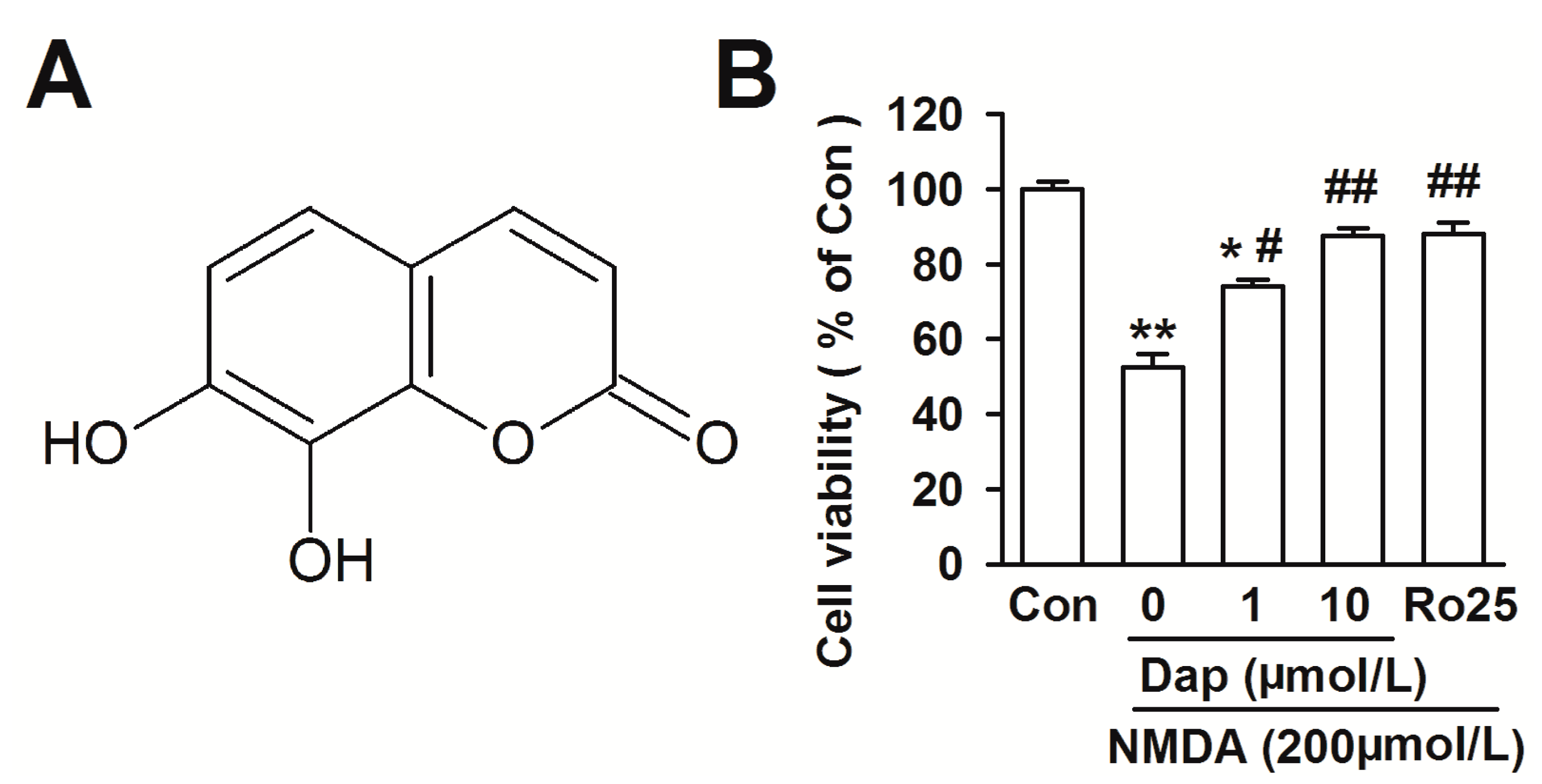
2.1. Effects of Dap on Cell Viability of Cortical Neurons
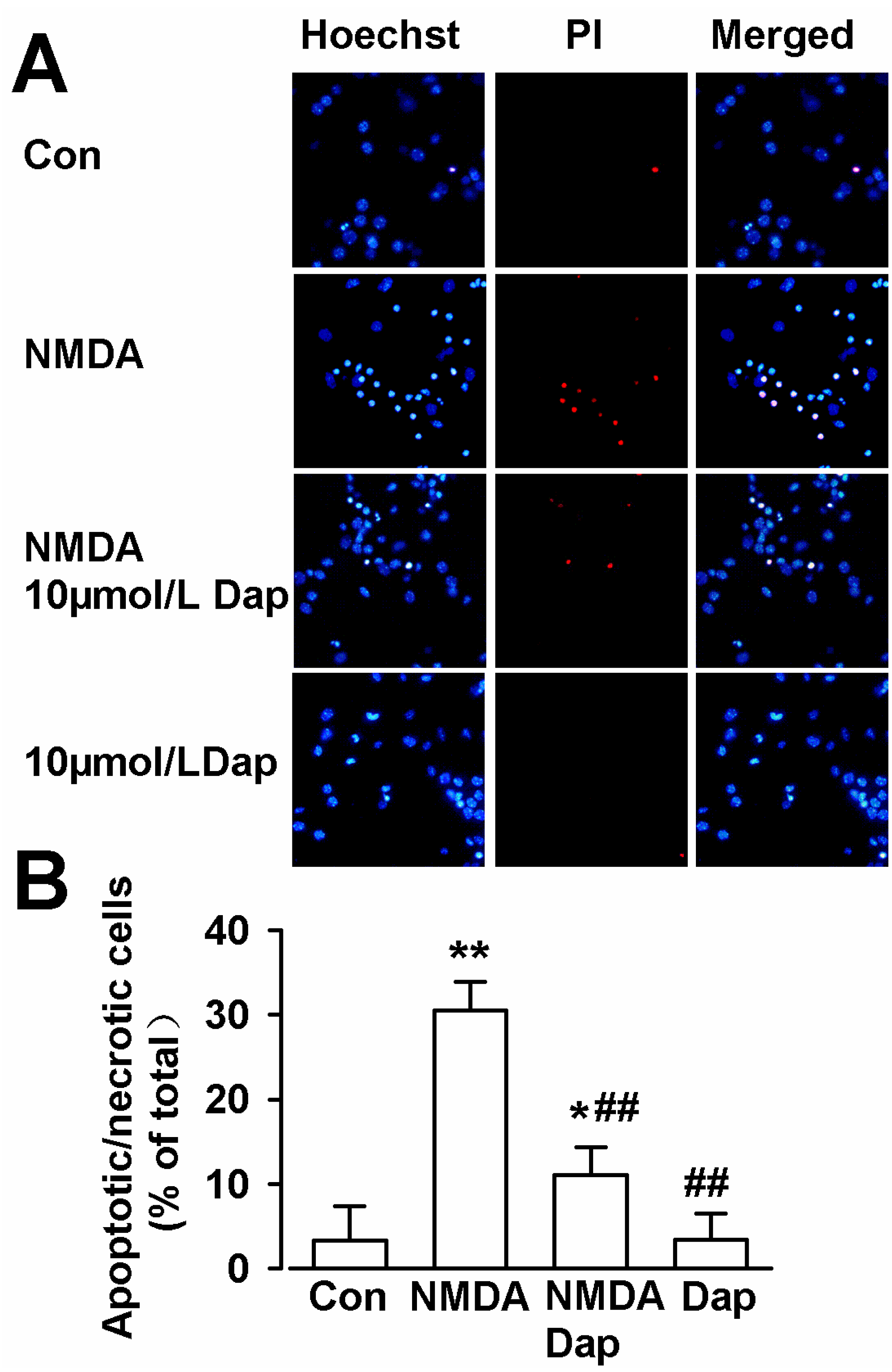
2.2. Anti-Apoptotic Activities of Dap
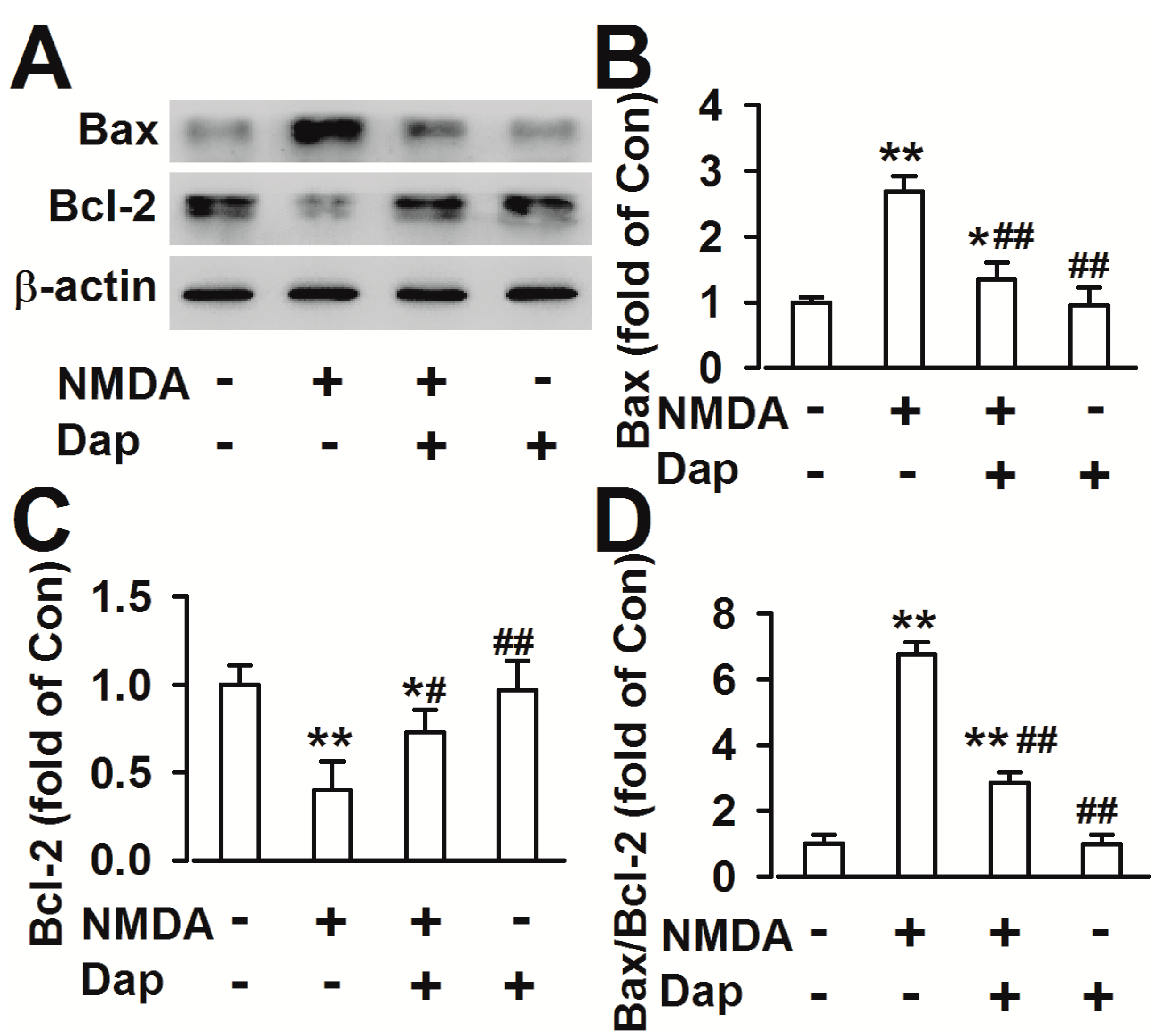
2.3. Effects of Dap on the Expression of Apoptotic Proteins


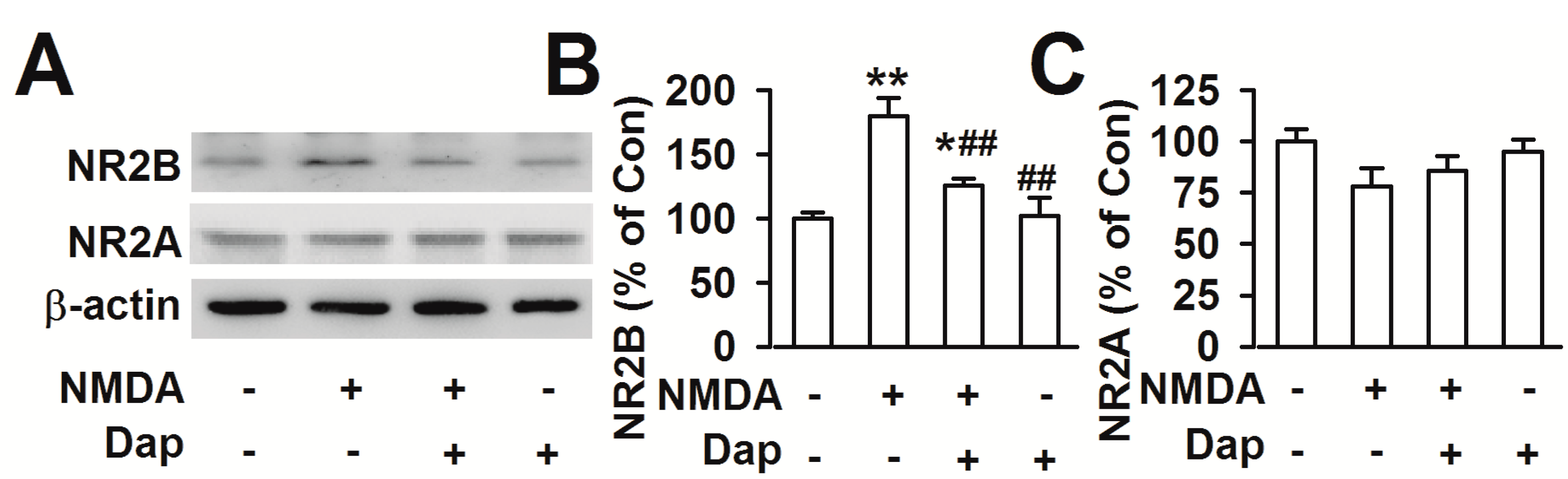
2.4. Effects of Dap on Expression of NR2A- and NR2B-Containing NMDARs

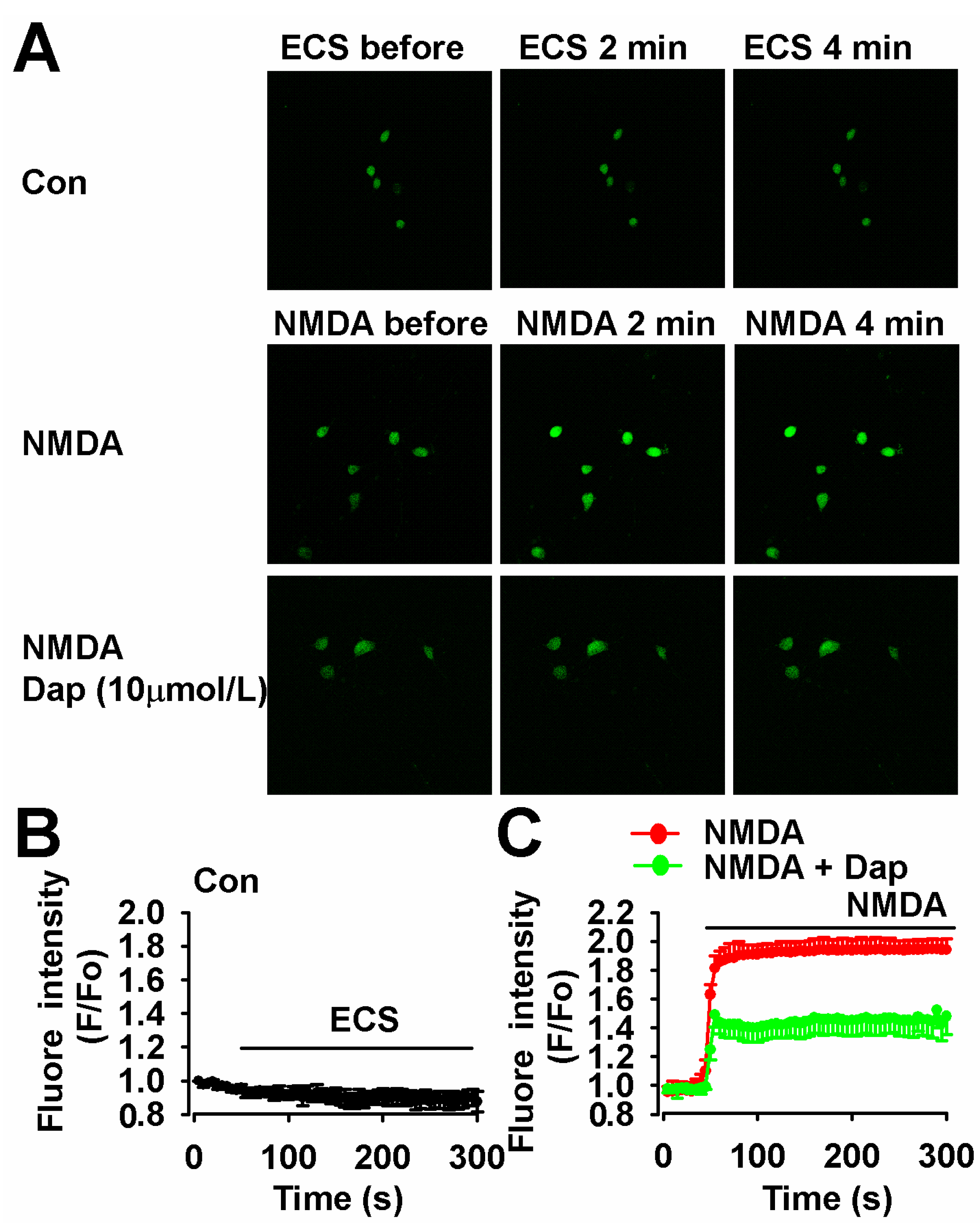
2.5. Inhibition of NMDA-Induced Ca2+ Overload by Dap
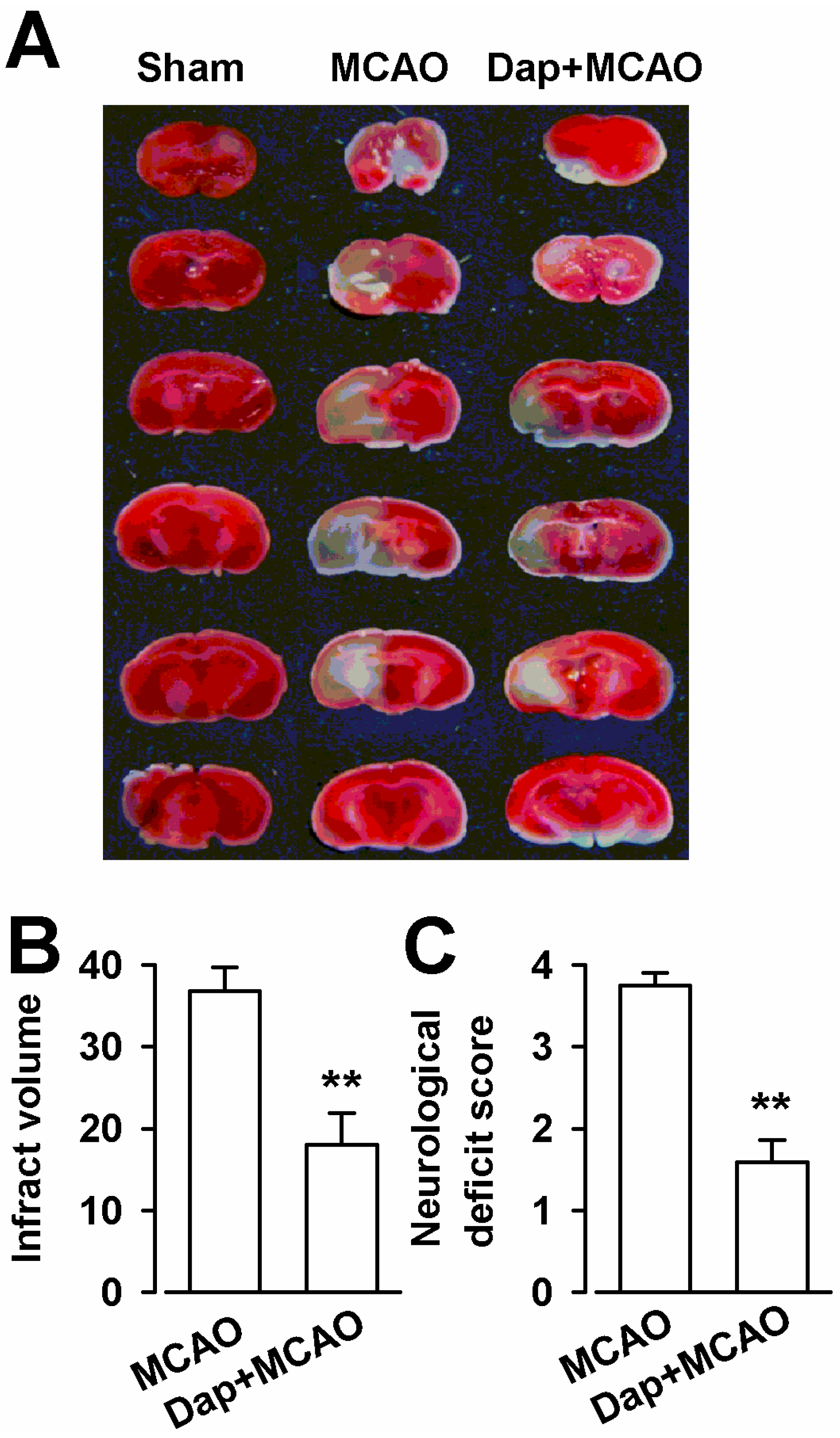
2.6. Neuroprotection of Dap against Cerebral Ischemia-Reperfusion Insult


3. Experimental
3.1. General Information
3.2. Primary Mouse Cortical Neuronal Culture
3.3. Cell Viability Analysis
3.4. Hoechst 33258 and PI Double Staining
3.5. Western Blot Analysis
3.6. Calcium Imaging
3.7. Middle Cerebral Artery Occlusion (MCAO)
3.8. Neurobehavioral Evaluation
3.9. Infarct Volume Assessment
3.10. Data Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yao, R.; Fu, Y.; Li, S.; Tu, L.; Zeng, X.; Kuang, N. Regulatory effect of daphnetin, a coumarin extracted from Daphne odora, on the balance of Treg and Th17 in collagen-induced arthritis. Eur. J. Pharmacol. 2011, 670, 286–294. [Google Scholar] [CrossRef]
- Liang, S.C.; Ge, G.B.; Liu, H.X.; Zhang, Y.Y.; Wang, L.M.; Zhang, J.W.; Yin, L.; Li, W.; Fang, Z.Z.; Wu, J.J.; et al. Identification and characterization of human UDP-glucuronosyltransferases responsible for the in vitro glucuronidation of daphnetin. Drug Metab. Dispos. 2010, 38, 973–980. [Google Scholar] [CrossRef]
- Yan, L.; Zhou, X.; Zhang, Z.; Luo, H.M. Neurotrophic effects of 7,8-dihydroxycoumarin in primary cultured rat cortical neurons. Neurosci. Bull. 2012, 28, 493–498. [Google Scholar] [CrossRef]
- Gonda, X. Basic pharmacology of NMDA receptors. Curr. Pharm. Des. 2012, 18, 1558–1567. [Google Scholar] [CrossRef]
- Hu, N.W.; Ondrejcak, T.; Rowan, M.J. Glutamate receptors in preclinical research on Alzheimer’s disease: Update on recent advances. Pharmacol. Biochem. Behav. 2012, 100, 855–862. [Google Scholar] [CrossRef]
- Paoletti, P. Molecular basis of NMDA receptor functional diversity. Eur. J. Neurosci. 2011, 33, 1351–1365. [Google Scholar] [CrossRef]
- Wang, Y.T.; Salter, M.W. Regulation of NMDA receptors by tyrosine kinases and phosphatases. Nature 1994, 369, 233–235. [Google Scholar] [CrossRef]
- Yang, E.B.; Zhao, Y.N.; Zhang, K.; Mack, P. Daphnetin, one of coumarin derivatives, is a protein kinase inhibitor. Biochem. Biophys. Res. Commun. 1999, 260, 682–685. [Google Scholar] [CrossRef]
- Du, G.; Tu, H.; Li, X.; Pei, A.; Chen, J.; Miao, Z.; Li, J.; Wang, C.; Xie, H.; Xu, X.; et al. Daphnetin, a natural coumarin derivative, provides the neuroprotection against glutamate-induced toxicity in HT22 cells and ischemic brain injury. Neurochem. Res. 2014, 39, 269–275. [Google Scholar] [CrossRef]
- Liu, S.B.; Zhang, N.; Guo, Y.Y.; Zhao, R.; Shi, T.Y.; Feng, S.F.; Wang, S.Q.; Yang, Q.; Li, X.Q.; Wu, Y.M.; et al. G-protein-coupled receptor 30 mediates rapid neuroprotective effects of estrogen via depression of NR2B-containing NMDA receptors. J. Neurosci. 2012, 32, 4887–4900. [Google Scholar] [CrossRef]
- Liu, Y.; Wong, T.P.; Aarts, M.; Rooyakkers, A.; Liu, L.; Lai, T.W.; Wu, D.C.; Lu, J.; Tymianski, M.; Craig, A.M.; et al. NMDA receptor subunits have differential roles in mediating excitotoxic neuronal death both in vitro and in vivo. J. Neurosci. 2007, 27, 2846–2857. [Google Scholar] [CrossRef]
- Stanika, R.I.; Pivovarova, N.B.; Brantner, C.A.; Watts, C.A.; Winters, C.A.; Andrews, S.B. Coupling diverse routes of calcium entry to mitochondrial dysfunction and glutamate excitotoxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 9854–9859. [Google Scholar] [CrossRef]
- Vander Jagt, T.A.; Connor, J.A.; Weiss, J.H.; Shuttleworth, C.W. Intracellular Zn2+ increases contribute to the progression of excitotoxic Ca2+ increases in apical dendrites of CA1 pyramidal neurons. Neuroscience 2009, 159, 104–114. [Google Scholar] [CrossRef]
- Liao, M.J.; Lin, L.F.; Zhou, X.; Zhou, X.W.; Xu, X.; Cheng, X.; Gao, Q.; Luo, H.M. Daphnetin prevents chronic unpredictable stress-induced cognitive deficits. Fundam. Clin. Pharmacol. 2013, 27, 510–516. [Google Scholar] [CrossRef]
- Jayanarayanan, S.; Smijin, S.; Peeyush, K.T.; Anju, T.R.; Paulose, C.S. NMDA and AMPA receptor mediated excitotoxicity in cerebral cortex of streptozotocin induced diabetic rat: Ameliorating effects of curcumin. Chem. Biol. Interact. 2013, 201, 39–48. [Google Scholar] [CrossRef]
- Spalloni, A.; Nutini, M.; Longone, P. Role of the N-methyl-d-aspartate receptors complex in amyotrophic lateral sclerosis. Biochim. Biophys. Acta 2012, 1832, 312–322. [Google Scholar] [CrossRef]
- Wang, C.; Pei, A.; Chen, J.; Yu, H.; Sun, M.L.; Liu, C.F.; Xu, X. A natural coumarin derivative esculetin offers neuroprotection on cerebral ischemia/reperfusion injury in mice. J. Neurochem. 2012, 121, 1007–1013. [Google Scholar] [CrossRef]
- Sun, M.; Hu, J.; Song, X.; Wu, D.; Kong, L.; Sun, Y.; Wang, D.; Wang, Y.; Chen, N.; Liu, G. Coumarin derivatives protect against ischemic brain injury in rats. Eur. J. Med. Chem. 2013, 67, 39–53. [Google Scholar] [CrossRef]
- Wang, H.; Wu, L.J.; Kim, S.S.; Lee, F.J.; Gong, B.; Toyoda, H.; Ren, M.; Shang, Y.Z.; Xu, H.; Liu, F.; et al. FMRP acts as a key messenger for dopamine modulation in the forebrain. Neuron 2008, 59, 634–647. [Google Scholar] [CrossRef]
- Brewer, G.J.; Torricelli, J.R.; Evege, E.K.; Price, P.J. Optimized survival of hippocampal neurons in B27-supplemented Neurobasal, a new serum-free medium combination. J. Neurosci. Res. 1993, 35, 567–576. [Google Scholar] [CrossRef]
- Li, Z.L.; Liu, J.C.; Liu, S.B.; Li, X.Q.; Yi, D.H.; Zhao, M.G. Improvement of vascular function by acute and chronic treatment with the GPR30 agonist G1 in experimental diabetes mellitus. PLoS One 2012, 7, e38787. [Google Scholar] [CrossRef]
- Zhang, X.N.; Li, J.M.; Yang, Q.; Feng, B.; Liu, S.B.; Xu, Z.H.; Guo, Y.Y.; Zhao, M.G. Anti-apoptotic effects of hyperoside via inhibition of NR2B-containing NMDA receptors. Pharmacol. Rep. 2010, 62, 949–955. [Google Scholar] [CrossRef]
- Wu, Y.; Peng, H.; Cui, M.; Whitney, N.P.; Huang, Y.; Zheng, J.C. CXCL12 increases human neural progenitor cell proliferation through Akt-1/FOXO3a signaling pathway. J. Neurochem. 2009, 109, 1157–1167. [Google Scholar] [CrossRef]
- Yang, L.; Li, X.B.; Yang, Q.; Zhang, K.; Zhang, N.; Guo, Y.Y.; Feng, B.; Zhao, M.G.; Wu, Y.M. The neuroprotective effect of praeruptorin C against NMDA-induced apoptosis through down-regulating of GluN2B-containing NMDA receptors. Toxicol. In Vitro 2013, 27, 908–914. [Google Scholar] [CrossRef]
- Tian, Z.; Yu, W.; Liu, H.B.; Zhang, N.; Li, X.B.; Zhao, M.G.; Liu, S.B. Neuroprotective effects of curculigoside against NMDA-induced neuronal excitoxicity in vitro. Food Chem. Toxicol. 2012, 50, 4010–4015. [Google Scholar] [CrossRef]
- Shi, T.Y.; Feng, S.F.; Xing, J.H.; Wu, Y.M.; Li, X.Q.; Zhang, N.; Tian, Z.; Liu, S.B.; Zhao, M.G. Neuroprotective effects of Salidroside and its analogue tyrosol galactoside against focal cerebral ischemia in vivo and H2O2-induced neurotoxicity in vitro. Neurotox. Res. 2012, 21, 358–367. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the daphnetin are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yang, L.; Yang, Q.; Zhang, K.; Li, Y.-J.; Wu, Y.-M.; Liu, S.-B.; Zheng, L.-H.; Zhao, M.-G. Neuroprotective Effects of Daphnetin against NMDA Receptor-Mediated Excitotoxicity. Molecules 2014, 19, 14542-14555. https://doi.org/10.3390/molecules190914542
Yang L, Yang Q, Zhang K, Li Y-J, Wu Y-M, Liu S-B, Zheng L-H, Zhao M-G. Neuroprotective Effects of Daphnetin against NMDA Receptor-Mediated Excitotoxicity. Molecules. 2014; 19(9):14542-14555. https://doi.org/10.3390/molecules190914542
Chicago/Turabian StyleYang, Le, Qi Yang, Kun Zhang, Yu-Jiao Li, Yu-Mei Wu, Shui-Bing Liu, Lian-He Zheng, and Ming-Gao Zhao. 2014. "Neuroprotective Effects of Daphnetin against NMDA Receptor-Mediated Excitotoxicity" Molecules 19, no. 9: 14542-14555. https://doi.org/10.3390/molecules190914542