
Distribution and Evolution of the Lectin Family in Soybean (Glycine max)



Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification and Distribution of Lectin Genes in Soybean

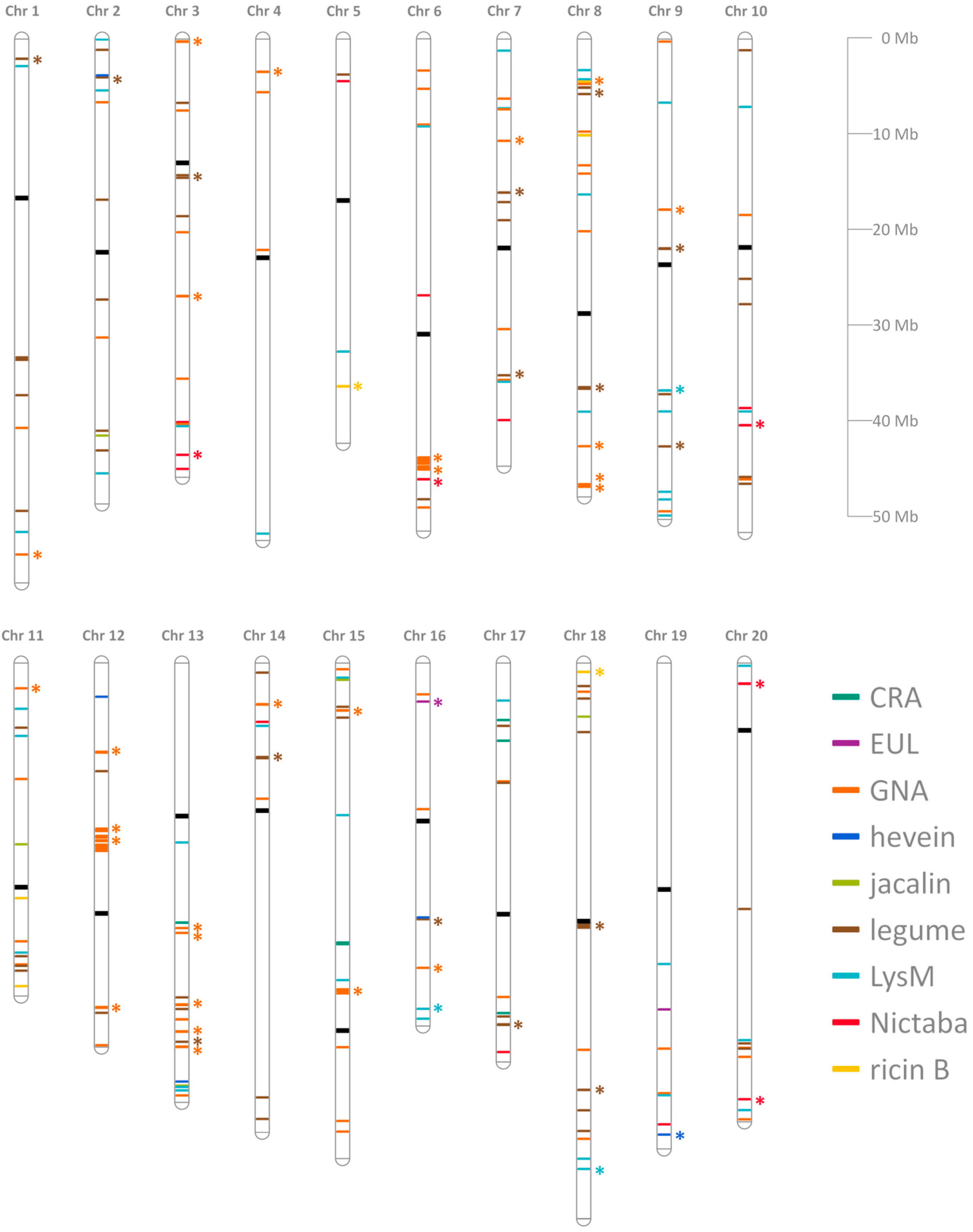{kind=link}
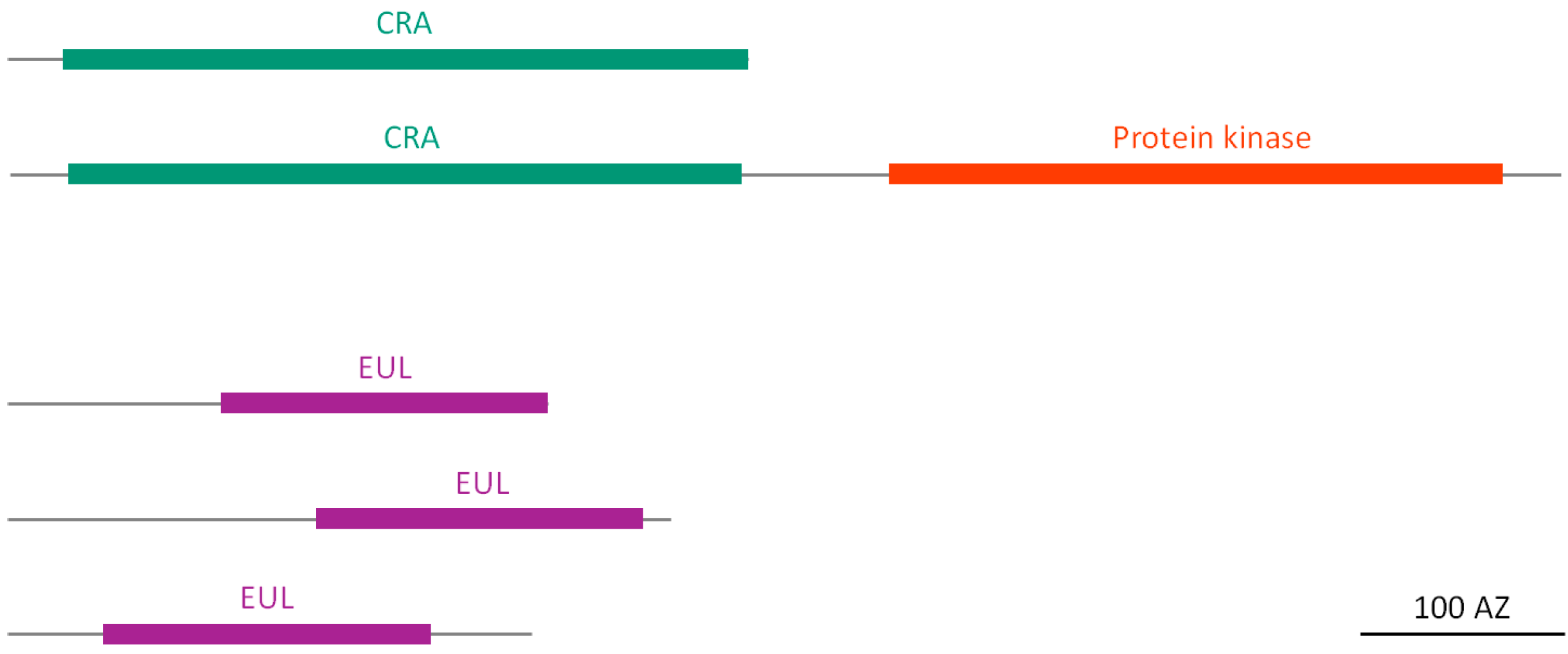{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lectin Domain | Predicted Genes | Percentage | Chromosome Localization |
|---|---|---|---|
| ABA domain | 0 | 0.0 | / |
| Amaranthin domain | 0 | 0.0 | / |
| CRA domain | 6 | 1.7 | 13, 15, 17 |
| Cyanovirin domain | 0 | 0.0 | / |
| EUL domain | 3 | 0.8 | 16, 19 |
| GNA domain | 166 | 46.2 | all except chr 5 |
| Hevein domain | 6 | 1.7 | 2, 12, 13, 16, 19 |
| Jacalin domain | 5 | 1.4 | 2, 11, 13, 15, 18 |
| Legume domain | 94 | 26.2 | all except chr 4 and chr 19 |
| LysM domain | 47 | 13.1 | all except chr 12 |
| Nictaba domain | 22 | 6.1 | 3, 5, 6, 7, 10, 14, 17, 19, 20 |
| Ricin B domain | 10 | 2.8 | 5, 8, 11, 18 |
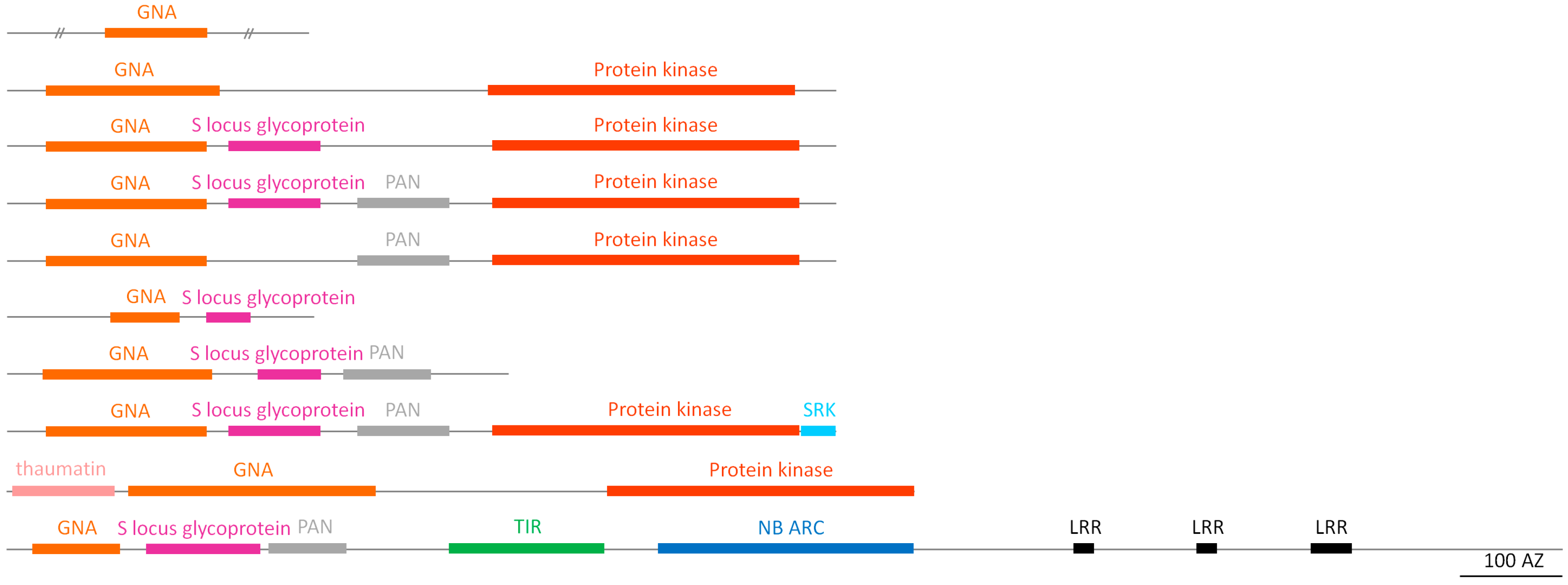
2.2. Domain Organization/Architecture of Putative Soybean Lectins

2.2.1. Homologs of Class V Chitinases (CRA)
| Lectin Family | Predicted Localization |
|---|---|
| CRA family | vacuole, membrane bound |
| Euonymus europaeus lectin family | nucleus, cytoplasm |
| Galanthus nivalis lectin family | vacuole, nucleus, cytoplasm or membrane bound |
| Hevein family | vacuole |
| Jacalin family | nucleus, cytoplasm |
| Legume family | vacuole, nucleus, cytoplasm or membrane bound |
| LysM family | vacuole, nucleus, cytoplasm or membrane bound |
| Nicotiana tabacum lectin family | nucleus, cytoplasm |
| Ricin B family | vacuole, nucleus, cytoplasm |

2.2.2. EUL Homologs
2.2.3. GNA Homologs

2.2.4. Hevein Homologs

2.2.5. Jacalin Homologs
2.2.6. Legume Homologs

2.2.7. LysM Homologs
2.2.8. Nictaba Homologs


2.2.9. Ricin B Homologs
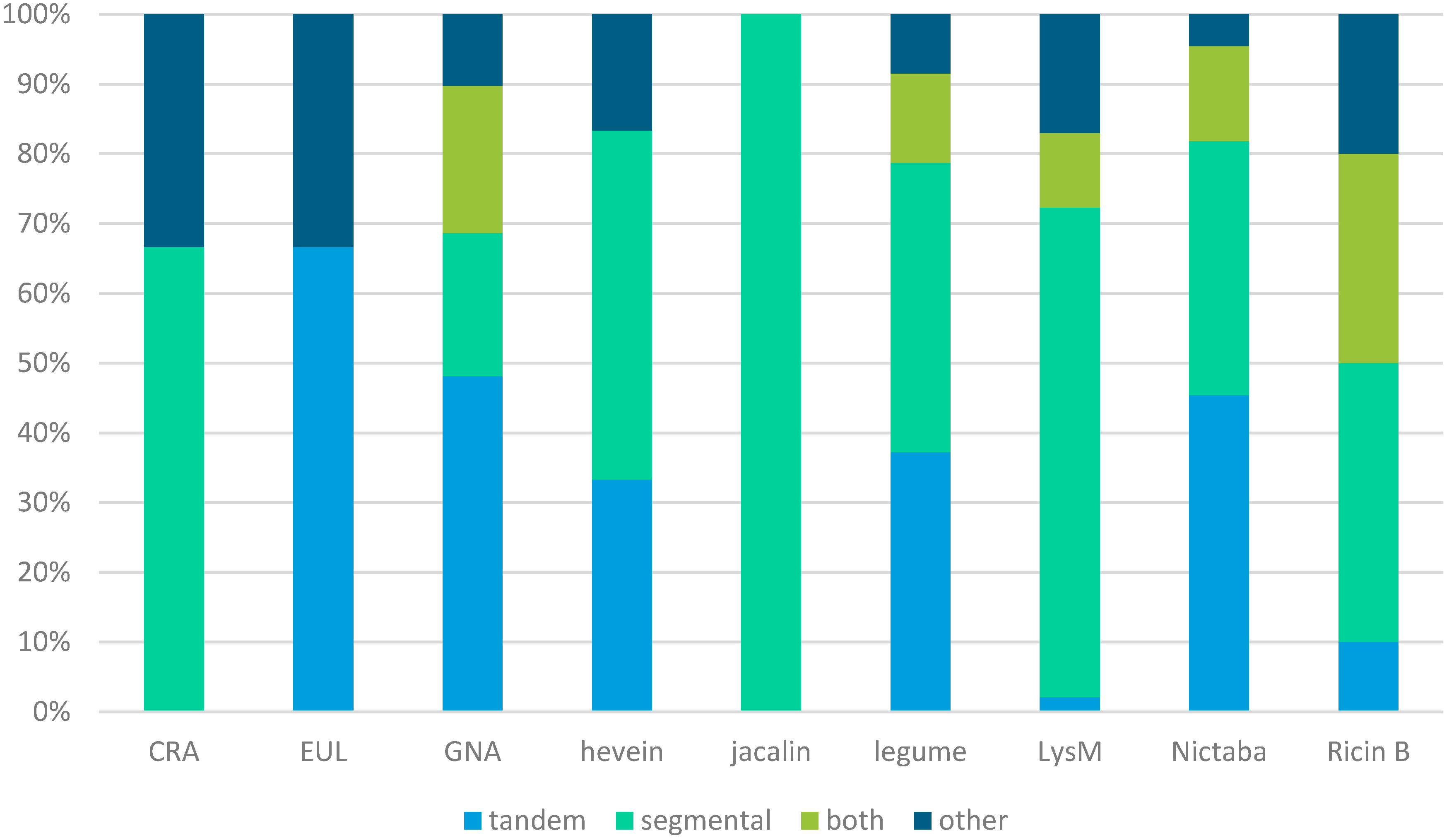
2.3. Tandem and Segmental Duplication Largely Contributed to the Expansion of Lectin Genes in Soybean
| Lectin Family (Number of Genes) | Tandem Duplication | Segmental Duplication | |
|---|---|---|---|
| Duplication Clusters | Genes Involved | Genes Involved | |
| CRA (6) | 0 | 0 | 4 |
| EUL (3) | 1 | 2 | 0 |
| GNA (166) | 26 | 114 | 69 |
| Hevein (6) | 1 | 2 | 3 |
| Jacalin (5) | 0 | 0 | 5 |
| Legume (94) | 16 | 49 | 51 |
| LysM (47) | 2 | 4 | 38 |
| Nictaba (22) | 5 | 13 | 11 |
| Ricin B (10) | 2 | 4 | 7 |

3. Discussion

4. Materials and Methods
4.1. Identification of Lectin Genes in the Soybean Genome
4.2. Construction of Chromosome Map
4.3. Analysis of Lectin Gene Expansion
4.4. Sequence Alignment and Phylogenetic Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Biswas, B.; Scott, P.T.; Gresshoff, P.M. Tree Legumes as Feedstock for Sustainable Biofuel Production: Opportunities and Challenges. J. Plant Physiol. 2011, 168, 1877–1884. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome Sequence of the Palaeopolyploid Soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Soystats. Available online: http://soystats.com/ (accessed on 3 November 2014).
- Graham, P.H.; Vance, C.P. Legumes: Importance and Constraints to Greater Use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Peumans, W.J.; van Damme, E.J.M. Lectins as Plant Defense Proteins. Plant Physiol. 1995, 109, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.M.; Peumans, W.J.; Barre, A.; Rougé, P. Plant Lectins: A Composite of Several Distinct Families of Structurally and Evolutionary Related Proteins with Diverse Biological Roles. CRC Crit. Rev. Plant Sci. 1998, 17, 575–692. [Google Scholar]
- Lannoo, N.; van Damme, E.J.M. Nucleocytoplasmic Plant Lectins. Biochim. Biophys. Acta 2010, 1800, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Lannoo, N.; van Damme, E.J.M. Lectin Domains at the Frontiers of Plant Defense. Front. Plant Sci. 2014, 5, 397:1–397:16. [Google Scholar]
- Van Damme, E.J.M.; Lannoo, N.; Peumans, W.J. Plant Lectins. Adv. Bot. Res. 2008, 48, 107–209. [Google Scholar]
- Van Damme, E.J.M. Lectins. In Lectins: Methods and Protocols; Hirabayashi, J., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2014; Volume 1200, pp. 3–13. [Google Scholar]
- Jiang, S.-Y.; Ma, Z.; Ramachandran, S. Evolutionary History and Stress Regulation of the Lectin Superfamily in Higher Plants. BMC Evol. Biol. 2010, 10, 79:1–79:24. [Google Scholar]
- Perez, S.; Sarkar, A.; Breton, C.; Drouillard, S.; Rivet, A.; Imberty, A. Glyco3D: A Portal for Structural Glycoscience. Available online: http://www.glyco3d.cermav.cnrs.fr (accessed on 4 November 2014).
- Vodkin, L.O.; Rhodes, P.R.; Goldberg, R.B. cA Lectin Gene Insertion Has the Structural Features of a Transposable Element. Cell 1983, 34, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Lis, H.; Sharon, N. The Biochemistry of Plant Lectins (phytohemagglutinins). Annu. Rev. Biochem. 1973, 42, 541–574. [Google Scholar] [CrossRef] [PubMed]
- Lotan, R.; Siegelman, H.W.; Lis, H.; Sharon, N. Subunit Structure of Soybean Agglutinin Subunit Structure of Soybean. J. Biol. Chem. 1974, 249, 1219–1224. [Google Scholar] [PubMed]
- Lis, H.; Sharon, N. Soybean Agglutinin—A Plant Glycoprotein. J. Biol. Chem. 1978, 253, 3468–3476. [Google Scholar] [PubMed]
- Pereira, M.E.A.; Kabat, E.A.; Sharon, N. Immunochemical Studies on the Specificity of Soybean Agglutinin. Carbohydr. Res. 1974, 37, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Nagai, K.; Yamaguchi, H. Direct Demonstration of the Essential Role of the Intramolecular Oligosaccharide Chains in the Folding and Assembly Soybean (Glycine max) Lectin Polypeptides. J. Biochem. 1993, 113, 123–125. [Google Scholar] [PubMed]
- Dessen, A.; Gupta, D.; Sabesan, S.; Brewer, C.F.; Sacchettini, J.C. X-Ray Crystal Structure of the Soybean Agglutinin Cross-Linked with a Biantennary Analog of the Blood Group I Carbohydrate Antigen. Biochemistry 1995, 34, 4933–4942. [Google Scholar] [CrossRef] [PubMed]
- Lis, H.; Fridman, C.; Sharon, N.; Katchalski, E. Multiple Hemagglutinins in Soybean. Arch. Biochem. Biophys. 1966, 8, 301–309. [Google Scholar] [CrossRef]
- Mandal, D.K.; Nieves, E.; Bhattacharyya, L.; Orr, G.A.; Roboz, J.; Yu, Q.; Brewer, C.F. Purification and Characterization of Three Isolectins of Soybean Agglutinin. Evidence for C-Terminal Truncation by Electrospray Ionization Mass Spectrometry. Eur. J. Biochem. 1994, 221, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Spilatro, S.R.; Cochran, G.R.; Walker, R.E.; Cablish, K.L.; Bittner, C. Characterization of a New Lectin of Soybean Vegetative Tissues. Plant Physiol. 1996, 110, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Fouquaert, E.; Peumans, W.J.; Vandekerckhove, T.T.M.; Ongenaert, M.; van Damme, E.J.M. Proteins with an Euonymus Lectin-like Domain Are Ubiquitous in Embryophyta. BMC Plant Biol. 2009, 9, 136:1–136:17. [Google Scholar] [CrossRef] [Green Version]
- Faruque, K.; Begam, R.; Deyholos, M.K. The Amaranthin-like Lectin (LuALL) Genes of Flax: A Unique Gene Family with Members Inducible by Defence Hormones. Plant Mol. Biol. Rep. 2014, 2014, 1–11. [Google Scholar]
- Delporte, A.; van Holle, S.; Lannoo, N.; van Damme, E.J.M. The Tobacco Lectin, Prototype of the Family of Nictaba-Related Proteins. Curr. Protein Pept Sci. 2015, in press. [Google Scholar]
- Voorrips, R. MapChart: Software for the Graphical Presentation of Linkage Maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.M.; Culerrier, R.; Barre, A.; Alvarez, R.; Rougé, P.; Peumans, W.J. A Novel Family of Lectins Evolutionarily Related to Class V Chitinases: An Example of Neofunctionalization in Legumes. Plant Physiol. 2007, 144, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Collinge, D.B.; Kragh, K.M.; Mikkelson, J.D.; Nielsen, K.K.; Rasmussen, U.; Vad, K. Plant Chitinases. Plant J. 1993, 3, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Melchers, L.S.; Apotheker-de Groot, M.; van der Knaap, J.A.; Ponstein, A.S.; Sela-Buurlage, M.B.; Bol, J.F.; Cornelissen, B.J.C.; van den Elzen, P.J.M.; Linthorst, H.J.M. A New Class of Tobacco Chitinases Homologous to Bacterial Exo-chitinases Displays Antifungal Activity. Plant J. 1994, 5, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Ramulu, H.G.; Drula, E.; Coutinho, P.M.; Henrissat, B. The Carbohydrate-Active Enzymes Database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, 490–495. [Google Scholar] [CrossRef]
- Van Damme, E.J.M.; Allen, A.K.; Peumans, W.J. Isolation and Characterization of a Lectin with Exclusive Specificity towards Mannose from Snowdrop (Galanthus nivalis) Bulbs. FEBS Lett. 1987, 215, 140–144. [Google Scholar] [CrossRef]
- Van Parijs, J.; Broekaert, W.F.; Goldstein, I.J.; Peumans, W.J. Hevein: An Antifungal Protein from Rubber-Tree (Hevea brasiliensis) Latex. Planta 1991, 183, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, J.-M.; Fritig, B.; Linthorst, H.J.M.; Meins, F.; Mikkelsen, J.D.; Ryals, J. A Revised Nomenclature for Chitinase Genes. Plant Mol. Biol. Rep. 1996, 14, 102–104. [Google Scholar] [CrossRef]
- Gijzen, M.; Kuflu, K.; Qutob, D.; Chernys, J.T. A Class I Chitinase from Soybean Seed Coat. J. Exp. Bot. 2001, 52, 2283–2289. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.S.; Appukuttan, P.S.; Basu, D. Alpha-D-Galactose-Specific Lectin from Jack Fruit (Artocarpus integra) Seed. J. Biosci. 1982, 4, 257–261. [Google Scholar] [CrossRef]
- Sastry, M.V.K.; Banarjee, P.; Patanjalili, S.R.; Swamy, M.J.; Swarnalatha, G.V.; Surolia, A. Analysis of Saccharide Binding to Artocarpus Integrifolia Lectin Reveals Specific Recognition of T-Antigen. J. Biol. Chem. 1986, 261, 11726–11733. [Google Scholar] [PubMed]
- Naganuma, T.; Hoshino, W.; Shikanai, Y.; Sato, R.; Liu, K.; Sato, S.; Muramoto, K.; Osada, M.; Yoshimi, K.; Ogawa, T. Novel Matrix Proteins of Pteria penguin Pearl Oyster Shell Nacre Homologous to the Jacalin-Related Β-Prism Fold Lectins. PLoS One 2014, 9, e112326. [Google Scholar] [CrossRef] [PubMed]
- Joris, B.; Englebert, S.; Chu, C.P.; Kariyama, R.; Daneo-Moore, L.; Shockman, G.D.; Ghuysen, J.M. Modular Design of the Enterococcus hirae Muramidase-2 and Streptococcus faecalis Autolysin. FEMS Microbiol. Lett. 1992, 70, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Limpens, E.; Franken, C.; Smit, P.; Willemse, J.; Bisseling, T.; Geurts, R. LysM Domain Receptor Kinases Regulating Rhizobial Nod Factor-Induced Infection. Science 2003, 302, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Radutoiu, S.; Madsen, L.H.; Madsen, E.B.; Felle, H.H.; Umehara, Y.; Grønlund, M.; Sato, S.; Nakamura, Y.; Tabata, S.; Sandal, N.; et al. Plant Recognition of Symbiotic Bacteria Requires Two LysM Receptor-like Kinases. Nature 2003, 425, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Indrasumunar, A.; Searle, I.; Lin, M.-H.; Kereszt, A.; Men, A.; Carroll, B.J.; Gresshoff, P.M. Nodulation Factor Receptor Kinase 1α Controls Nodule Organ Number in Soybean (Glycine max L. Merr). Plant J. 2011, 65, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Indrasumunar, A.; Kereszt, A.; Searle, I.; Miyagi, M.; Li, D.; Nguyen, C.D.T.; Men, A.; Carroll, B.J.; Gresshoff, P.M. Inactivation of Duplicated Nod Factor Receptor 5 (NFR5) Genes in Recessive Loss-of-Function Non-Nodulation Mutants of Allotetraploid Soybean (Glycine max L. Merr.). Plant Cell Physiol. 2010, 51, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Peumans, W.J.; Hause, B.; Bras, J.; Kumar, M.; Proost, P.; Barre, A.; Rougé, P.; van Damme, E.J.M. Jasmonate Methyl Ester Induces the Synthesis of a Cytoplasmic/nuclear Chitooligosaccharide-Binding Lectin in Tobacco Leaves. FASEB J. 2002, 16, 905–907. [Google Scholar] [PubMed]
- Barbieri, L.; Valbonesi, P.; Gorini, P.; Pession, A.; Stirpe, F. Polynucleotide: Adenosine Glycosidase Activity of Saporin-L1: Effect on DNA, RNA and Poly (A). Biochem. J. 1996, 319, 507–513. [Google Scholar] [PubMed]
- Roulin, A.; Auer, P.L.; Libault, M.; Schlueter, J.; Farmer, A.; May, G.; Stacey, G.; Doerge, R.W.; Jackson, S.A. The Fate of Duplicated Genes in a Polyploid Plant Genome. Plant J. 2013, 73, 143–153. [Google Scholar] [CrossRef]
- Lee, T.-H.; Tang, H.; Wang, X.; Paterson, A.H. PGDD: A Database of Gene and Genome Duplication in Plants. Nucleic Acids Res. 2013, 41, 1152–1158. [Google Scholar] [CrossRef]
- Liener, I.E.; Pallansch, M.J. Purification of a Toxic Substance from Defatted Soy Bean Flour. J. Biol. Chem. 1952, 197, 29–36. [Google Scholar] [PubMed]
- Ohno, S. Evolution by Gene Duplication; Springer: Berlin, Germany, 1970. [Google Scholar]
- Van de Peer, Y.; Maere, S.; Meyer, A. The Evolutionary Significance of Ancient Genome Duplications. Nat. Rev. Genet. 2009, 10, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Force, A. The Probability of Duplicate Gene Preservation by Subfunctionalization. Genetics 2000, 154, 459–473. [Google Scholar] [PubMed]
- Bai, C.; Sen, P.; Hofmann, K.; Ma, L.; Goebl, M.; Harper, J.M.; Elledge, S.J. SKP1 Connects Cell Cycle Regulators to the Ubiquitin Proteolysis Machinery through a Novel Motif, the F-Box. Cell 1996, 86, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Petroski, M.D.; Deshaies, R.J. Function and Regulation of Cullin-RING Ubiquitin Ligases. Nat. Rev. Mol. Cell Biol. 2005, 6, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Lannoo, N.; Peumans, W.J.; van Damme, E.J.M. Do F-Box Proteins with a C-Terminal Domain Homologous with the Tobacco Lectin Play a Role in Protein Degradation in Plants? Biochem. Soc. Trans. 2008, 36, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Ma, H.; Nei, M.; Kong, H. Evolution of F-Box Genes in Plants: Different Modes of Sequence Divergence and Their Relationships. Proc. Natl. Acad. Sci. USA 2009, 106, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, T.; Yoshida, Y.; Kumanomidou, T.; Hasegawa, Y.; Suzuki, A.; Yamane, T.; Tanaka, K. Structural Basis for the Selection of Glycosylated Substrates by SCF(Fbs1) Ubiquitin Ligase. Proc. Natl. Acad. Sci. USA 2007, 104, 5777–5781. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-C.; Cannon, S.B.; Stacey, G. Evolutionary Genomics of LysM Genes in Land Plants. BMC Evol. Biol. 2009, 9, 183:1–183:13. [Google Scholar]
- Henrissat, B.; Claeyssens, M.; Tomme, P.; Lemesle, L.; Mornon, J.P. Cellulase Families Revealed by Hydrophobic Cluster Analysis. Gene 1989, 81, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Kasprzewska, A. Plant Chitinases—Regulation and Function. Cell. Mol. Biol. Lett. 2003, 8, 809–824. [Google Scholar] [PubMed]
- Svensson, B.; Svendsen, I.; Højrup, P.; Roepstorff, P.; Ludvigsen, S.; Poulsen, F.M. Primary Structure of Barwin: A Barley Seed Protein Closely Related to the C-Terminal Domain of Proteins Encoded by Wound-Induced Plant Genes. Biochemistry 1992, 31, 8767–8770. [Google Scholar] [CrossRef] [PubMed]
- Ludvigsen, S.; Poulsen, F.M. Three-Dimensional Structure in Solution of Barwin, a Protein from Barley Seed. Biochemistry 1992, 31, 8783–8789. [Google Scholar] [CrossRef] [PubMed]
- Stanford, A.; Bevan, M.; Northcote, D. Differential Expression within a Family of Novel Wound-Induced Genes in Potato. Mol. Gen. Genet. 1989, 215, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.; Uknes, S.; Lawton, K.; Winter, A.M.; Chandler, D.; DiMaio, J.; Novitzky, R.; Ward, E.; Ryals, J. Regulation of a Hevein-like Gene in Arabidopsis. Mol. Plant Microbe Interact. 1993, 6, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Ponstein, A.S.; Bres-Vloemans, S.A.; Sela-Buurlage, M.B.; van den Elzen, P.J.; Melchers, L.S.; Cornelissen, B.J. A Novel Pathogen- and Wound-Inducible Tobacco (Nicotiana tabacum) Protein with Antifungal Activity. Plant Physiol. 1994, 104, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.C.; Lin, J.H.; Chua, A.C.N.; Chung, T.Y.; Tsai, I.C.; Tzen, J.T.C.; Chou, W.M. Cloning and Expression of Pathogenesis-Related Protein 4 from Jelly Fig (Ficus awkeotsang makino) Achenes Associated with Ribonuclease, Chitinase and Anti-Fungal Activities. Plant Physiol. Biochem. 2012, 56, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Handa, A.; Hasegawa, P.M.; Bressan, R.A. Proteins Associated with Adaptation of Cultured Tobacco Cells to NaCl. Plant Physiol. 1985, 79, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Woloshuk, C.P.; Meulenhoff, J.S.; Sela-Buurlage, M.; van den Elzen, P.J.M.; Cornelissen, B.J.C. Pathogen-Induced Proteins with Inhibitory Activity toward Phytophthora Infestans. Plant Cell 1991, 3, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-Related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed]
- Hammond-Kosack, K.E.; Jones, J.D.G. Plant Disease Resistance Genes. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 575–607. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Gochicoa, M.T.; Camut, S.; Timmers, A.C.J.; Niebel, A.; Hervé, C.; Boutet, E.; Bono, J.-J.; Imbery, A.; Cullimore, J.V. Characterization of Four Lectin-like Receptor Kinases Expressed in Roots of Medicago truncatula. Structure, Location, Regulation of Expression, and Potential Role in the Symbiosis with Sinorhizobium meliloti. Plant Physiol. 2003, 133, 1893–1910. [Google Scholar] [CrossRef] [PubMed]
- Bouwmeester, K.; Govers, F. Arabidopsis L-Type Lectin Receptor Kinases: Phylogeny, Classification, and Expression Profiles. J. Exp. Bot. 2009, 60, 4383–4396. [Google Scholar] [CrossRef] [PubMed]
- Tanksley, S.D.; Loaiza-Figueroa, F. Gametophytic Self-Incompatibility Is Controlled by a Single Major Locus on Chromosome 1 in Lycopersicon peruvianum. Proc. Natl. Acad. Sci. USA 1985, 82, 5093–5096. [Google Scholar] [CrossRef] [PubMed]
- Tordai, H.; Bányai, L.; Patthy, L. The PAN Module: The N-Terminal Domains of Plasminogen and Hepatocyte Growth Factor Are Homologous with the Apple Domains of the Prekallikrein Family and with a Novel Domain Found in Numerous Nematode Proteins. FEBS Lett. 1999, 461, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Xing, S.; Li, M.; Liu, P. Evolution of S-Domain Receptor-like Kinases in Land Plants and Origination of S-Locus Receptor Kinases in Brassicaceae. BMC Evol. Biol. 2013, 13, 69:1–69:11. [Google Scholar] [CrossRef]
- Sun, X.L.; Yu, Q.Y.; Tang, L.L.; Ji, W.; Bai, X.; Cai, H.; Liu, X.F.; Ding, X.D.; Zhu, Y.M. GsSRK, a G-Type Lectin S-Receptor-like Serine/threonine Protein Kinase, Is a Positive Regulator of Plant Tolerance to Salt Stress. J. Plant Physiol. 2013, 170, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.C.; Wang, Z.; Zhang, X.; Le, M.H.; Sun, J.; Xu, D.; Cheng, J.; Stacey, G. Evolutionary Dynamics of Protein Domain Architecture in Plants. BMC Evol. Biol. 2012, 12, 6:1–6:12. [Google Scholar] [CrossRef]
- Kersting, A.R.; Bornberg-Bauer, E.; Moore, A.D.; Grath, S. Dynamics and Adaptive Benefits of Protein Domain Emergence and Arrangements during Plant Genome Evolution. Genome Biol. Evol. 2012, 4, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.; Farias, C.M.S.A.; del Sol, F.G.; Santos, C.F.; Grangeiro, T.B.; Nagano, C.S.; Cavada, B.S.; Calvete, J.J. The Amino-Acid Sequence of the Glucose/mannose-Specific Lectin Isolated from Parkia platycephala Seeds Reveals Three Tandemly Arranged Jacalin-Related Domains. Eur. J. Biochem. 2001, 268, 4414–4422. [Google Scholar] [CrossRef] [PubMed]
- Agostino, M.; Velkov, T.; Dingjan, T.; Williams, S.J.; Yuriev, E.; Ramsland, P.A. The Carbohydrate-Binding Promiscuity of Euonymus europaeus Lectin Is Predicted to Involve a Single Binding Site. Glycobiology 2015, 25, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Cavada, B.S.; Moreno, F.B.B.; da Rocha, B.A.M.; de Azevedo, W.F.J.; Castellón, R.E.R.; Goersch, G.V.; Nagano, C.S.; de Souza, E.P.; Nascimento, K.S.; Radis-Baptista, G.; et al. cDNA Cloning and 1.75 Å Crystal Structure Determination of PPL2, an Endochitinase and N-Acetylglucosamine-Binding Hemagglutinin from Parkia platycephala Seeds. FEBS J. 2006, 273, 3962–3974. [Google Scholar] [CrossRef] [PubMed]
- Patil, D.N.; Datta, M.; Dev, A.; Dhindwal, S.; Singh, N.; Dasauni, P.; Kundu, S.; Sharma, A.K.; Tomar, S.; Kumar, P. Structural Investigation of a Novel N-Acetyl Glucosamine Binding Chi-Lectin Which Reveals Evolutionary Relationship with Class III Chitinases. PLoS One 2013, 8, e63779. [Google Scholar] [CrossRef] [PubMed]
- Stefanowicz, K.; Lannoo, N.; Proost, P.; van Damme, E.J.M. Arabidopsis F-Box Protein Containing a Nictaba-Related Lectin Domain Interacts with N-Acetyllactosamine Structures. FEBS Open Bio 2012, 2, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Fouquaert, E.; van Damme, E.J.M. Promiscuity of the Euonymus Carbohydrate-Binding Domain. Biomolecules 2012, 2, 415–434. [Google Scholar] [CrossRef] [PubMed]
- Loris, R.; Hamelryck, T.; Bouckaert, J.; Wyns, L. Legume Lectin Structure. Biochim. Biophys. Acta 1998, 1383, 9–36. [Google Scholar] [CrossRef] [PubMed]
- Rougé, P.; Peumans, W.J.; Barre, A.; van Damme, E.J.M. A Structural Basis for the Difference in Specificity between the Two Jacalin-Related Lectins from Mulberry (Morus nigra) Bark. Biochem. Biophys. Res. Commun. 2003, 304, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Fouquaert, E.; Smith, D.F.; Peumans, W.J.; Proost, P.; Balzarini, J.; Savvides, S.N.; van Damme, E.J.M. Related Lectins from Snowdrop and Maize Differ in Their Carbohydrate-Binding Specificity. Biochem. Biophys. Res. Commun. 2009, 380, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Wang, Y.; Zhou, X.; Xie, Y.; Wu, H.; Gao, X. Expression of Soybean Lectin in Transgenic Tobacco Results in Enhanced Resistance to Pathogens and Pests. Plant Sci. 2013, 211, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Shukle, R.H.; Murdock, L.L. Lipoxygenase Trypsin Inhibitor, and Lectin from Soybeans: Effects on Larval Growth of Manduca sexta (Lepidoptera: Sphingidae). Environ. Entomol. 1983, 12, 787–791. [Google Scholar] [CrossRef]
- Singh, K.; Kaur, M.; Rup, P.J.; Singh, J. Exploration for Anti-Insect Properties of Lectin from Seeds of Soybean (Glycine max) Using Bactrocera cucurbitae as a Model. Entomology 2006, 34, 463–473. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: a comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-Scale Protein Function Classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High-Quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539:1–539:6. [Google Scholar]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete Genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Soybean Genetic Map. Available online: http://soybase.org/ (accessed on 14 January 2015).
- Yin, G.; Xu, H.; Xiao, S.; Qin, Y.; Li, Y.; Yan, Y.; Hu, Y. The Large Soybean (Glycine max) WRKY TF Family Expanded by Segmental Duplication Events and Subsequent Divergent Selection among Subgroups. BMC Plant Biol. 2013, 13, 148:1–148:19. [Google Scholar] [CrossRef]
- Sample Availability: Samples are not available from authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Holle, S.; Van Damme, E.J.M. Distribution and Evolution of the Lectin Family in Soybean (Glycine max). Molecules 2015, 20, 2868-2891. https://doi.org/10.3390/molecules20022868
Van Holle S, Van Damme EJM. Distribution and Evolution of the Lectin Family in Soybean (Glycine max). Molecules. 2015; 20(2):2868-2891. https://doi.org/10.3390/molecules20022868
Chicago/Turabian StyleVan Holle, Sofie, and Els J. M. Van Damme. 2015. "Distribution and Evolution of the Lectin Family in Soybean (Glycine max)" Molecules 20, no. 2: 2868-2891. https://doi.org/10.3390/molecules20022868
APA StyleVan Holle, S., & Van Damme, E. J. M. (2015). Distribution and Evolution of the Lectin Family in Soybean (Glycine max). Molecules, 20(2), 2868-2891. https://doi.org/10.3390/molecules20022868