
Comparative Study of Two Table Grape Varieties with Contrasting Texture during Cold Storage

Abstract
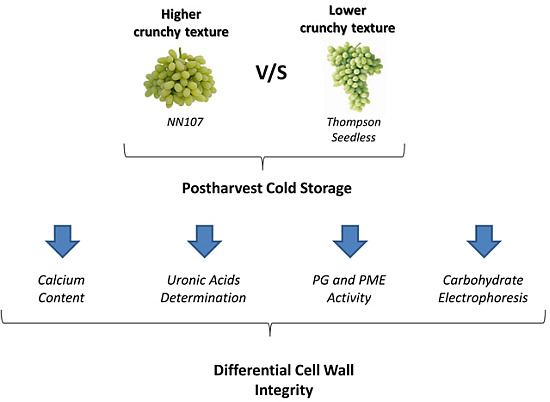
:
1. Introduction
2. Results and Discussion
2.1. Phenotypic Analysis of Grape Berries

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thompson Seedless | NN107 | |||
|---|---|---|---|---|
| Storage Time (days) | Curve Area (N·mm) | Rate Relative to Harvest (%) | Curve Area (N·mm) | Rate Relative to Harvest (%) |
| 0 (harvest) | 84 ± 10a | 100 | 182 ± 38a * | 100 |
| 30 | 84 ± 13a | 100 (0) | 166 ± 22ab * | 91.2 (8.8) |
| 30 + SL | 80 ± 12ab | 95.2 (4.8) | 158 ± 17bc * | 86.8 (4.4) |
| 60 | 75 ± 10b | 89.3 (5.9) | 150 ± 19bc * | 82.4 (4.4) |
| 60 + SL | 65 ± 11c | 77.4 (11.9) | 145 ± 25c * | 79.7 (2.7) |
| Thompson Seedless | ||||||
|---|---|---|---|---|---|---|
| Berry diameter | Soluble solids content | Titratable acidity | ||||
| Storage Time (days) | Mean Value (mm) | Rate Relative to Harvest (%) | Mean Value (%w/w) | Rate Relative to Harvest (%) | Mean Value (g·L−1) | Rate Relative to Harvest (%) |
| 0 (harvest) | 19.0 ± 1.2ab | 100 | 19.2 ± 1.6c | 100 | 0.4 ± 0.0a | 100 |
| 30 | 19.2 ± 1.3a | 101.1 (−1.1) | 20.3 ± 1.0ab | 105.7 (−5.7) | 0.3 ± 0.0b | 75 (25) |
| 30 + SL | 18.2 ± 1.1b | 95.8 (5.3) | 21.0 ± 0.8a | 109.4 (−3.7) | 0.3 ± 0.0b | 75 (0) |
| 60 | 19.2 ± 1.0a | 101.1 (−5.3) | 20.1 ± 1.0b | 104.7 (4.7) | 0.3 ± 0.0b | 75 (0) |
| 60 + SL | 18.6 ± 1.0ab | 97.9 (3.2) | 20.8 ± 1.1ab | 108.3 (−3.6) | 0.4 ± 0.0a | 100 (−25) |
| NN107 | ||||||
|---|---|---|---|---|---|---|
| Berry Diameter | Soluble Solids Content | Titratable Acidity | ||||
| Storage Time (days) | Mean Value (mm) | Rate Relative to Harvest (%) | Mean Value (%w/w) | Rate Relative to Harvest (%) | Mean Value (g·L−1) | Rate Relative to Harvest (%) |
| 0 (harvest) | 20.7 ± 1.5a | 100 | 18.2 ± 0.9b | 100 | 0.4 ± 0.0a | 100 |
| 30 | 20.3 ± 1.1ab | 98.1 (1.9) | 18.1 ± 0.6b | 99.5 (0.5) | 0.4 ± 0.0a | 100 (0) |
| 30 + SL | 19.8 ± 0.8b | 95.7 (2.4) | 18.6 ± 0.8b | 102.2 (−2.7) | 0.4 ± 0.0a | 100 (0) |
| 60 | 20.0 ± 0.9ab | 96.6 (−0.9) | 18.7 ± 0.8b | 102.7 (−0.5) | 0.4 ± 0.0a | 100 (0) |
| 60 + SL | 19.7 ± 1.3b | 95.2 (1.4) | 19.6 ± 1.1a | 107.7 (−5.0) | 0.4 ± 0.0a | 100 (0) |
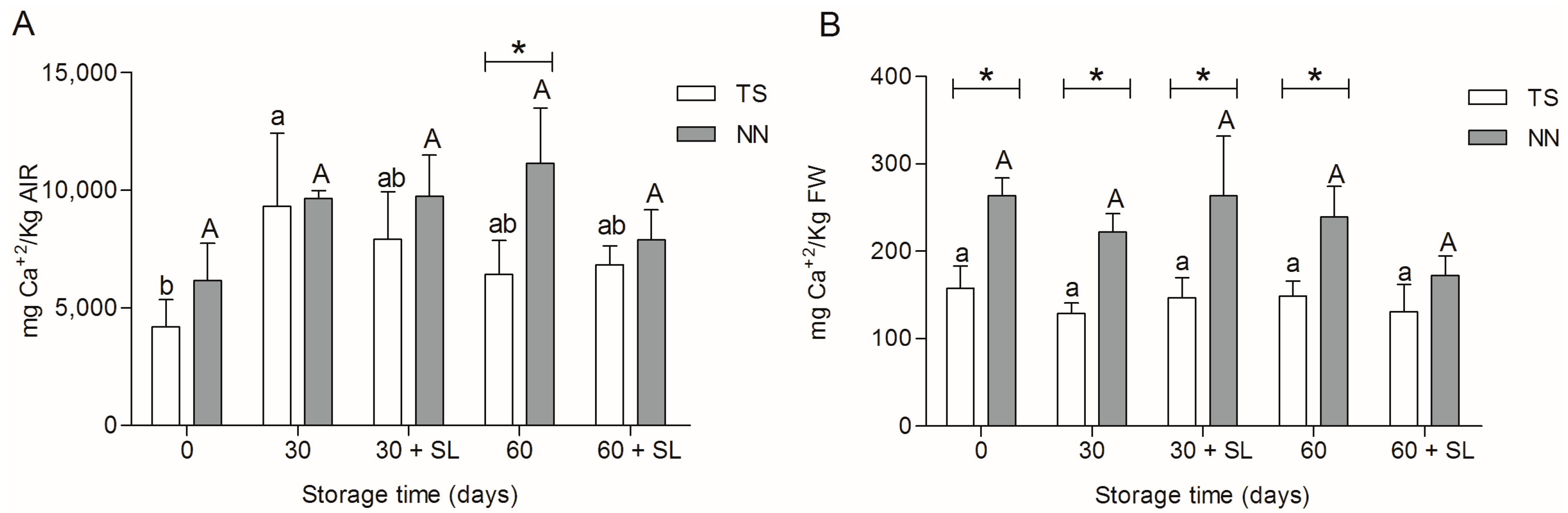
2.2. Calcium Content

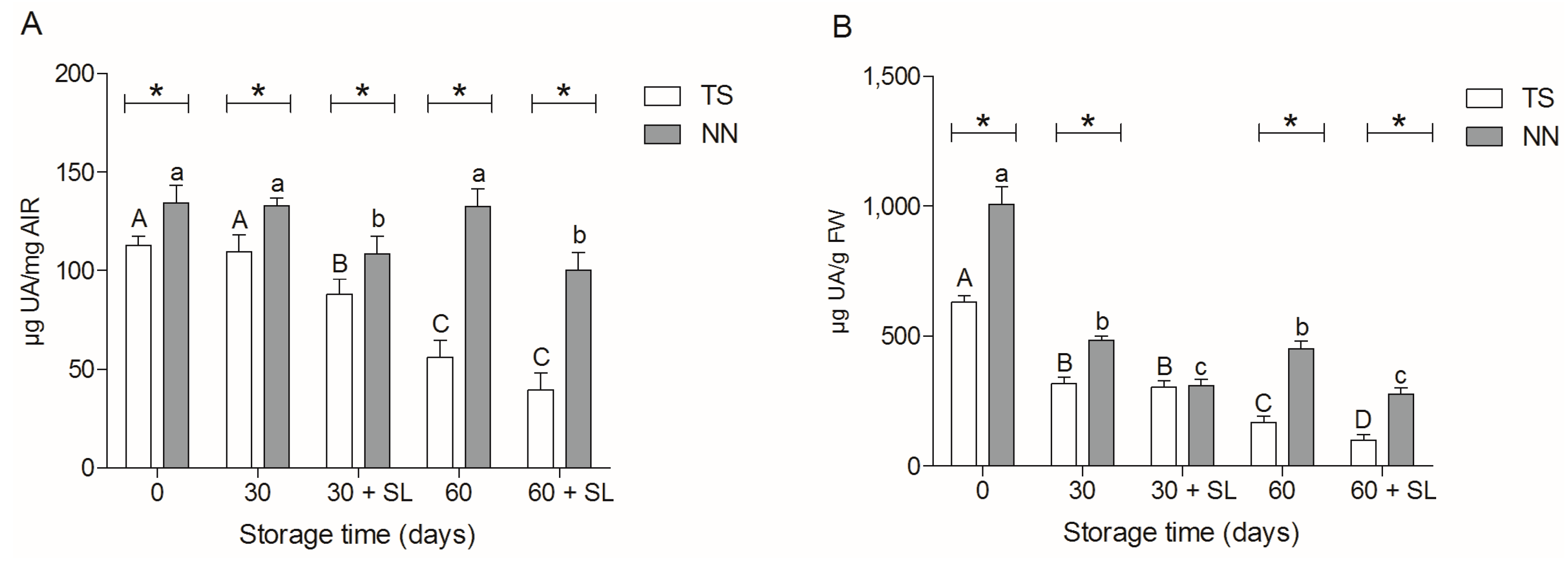
2.3. Uronic Acid Determination

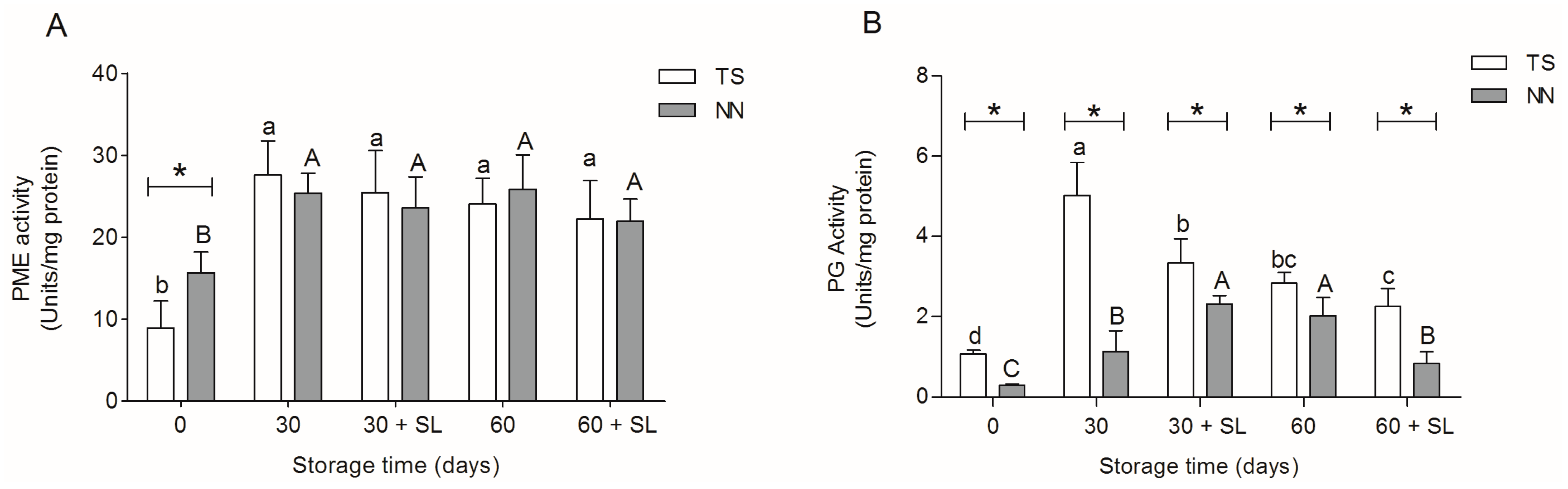
2.4. Activity of Cell Wall Modifying Enzymes

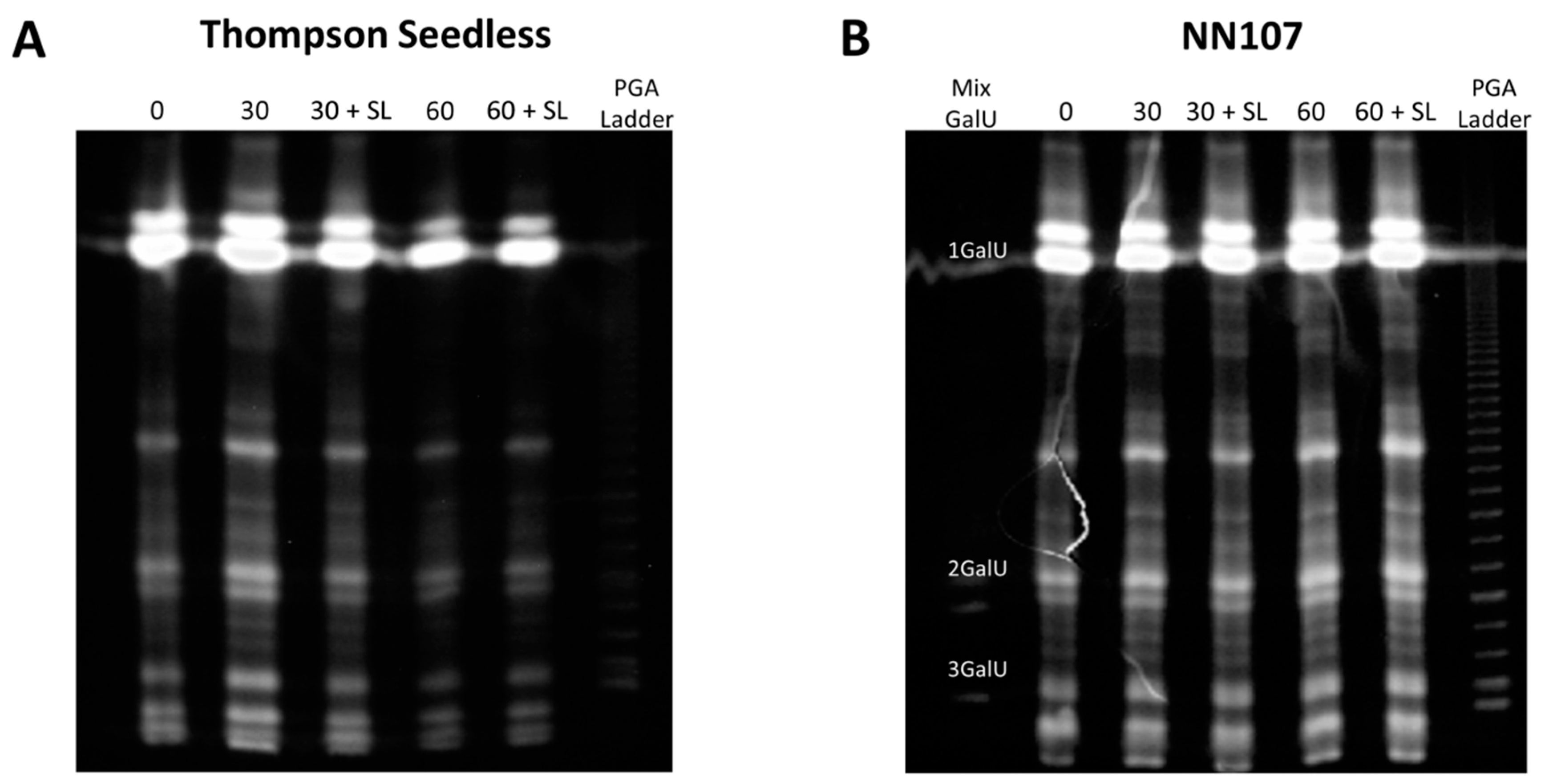
2.5. Carbohydrate Electrophoresis

3. Experimental Section
3.1. Plant Material and Maturity Parameters
3.2. Texture Analysis
3.3. Alcohol-Insoluble Residue Preparation
3.4. Uronic Acid Analysis
3.5. Polysaccharide Analysis Using Carbohydrate Gel Electrophoresis
3.6. Calcium Quantification
3.7. Protein Extraction
3.8. Pectin Methylesterase Activity
3.9. Polygalacturonase Activity
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Valverde, J.M.; Guillen, F.; Martinez-Romero, D.; Castillo, S.; Serrano, M.; Valero, D. Improvement of table grapes quality and safety by the combination of modified atmosphere packaging (MAP) and eugenol, menthol, or thymol. J. Agr. Food Chem. 2005, 53, 7458–7464. [Google Scholar] [CrossRef]
- Sato, A.; Yamane, H.; Hirakawa, N.; Otobe, K.; Yamada, M. Varietal differences in the texture of grape berries measured by penetration tests. Vitis 1997, 36, 7–10. [Google Scholar]
- Ruiz, S.; Moyano, A.; Navia, P. Acumulación de compuestos nitrogenados en relación al problema de baya blanda en uva de mesa. Agricultura Técnica 2004, 64, 426–430. [Google Scholar] [CrossRef]
- Nunan, K.J.; Sims, I.M.; Bacic, A.; Robinson, S.P.; Fincher, G.B. Changes in cell wall composition during ripening of grape berries. Plant Physiol. 1998, 118, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wu, Y.; Li, Y. Changes in firmness, cell wall composition and cell wall hydrolases of grapes stored in high oxygen atmospheres. Food Res. Int. 2005, 38, 769–776. [Google Scholar] [CrossRef]
- Brummell, D.A. Cell wall disassembly in ripening fruit. Funct. Plant Biol. 2006, 33, 103–119. [Google Scholar] [CrossRef]
- Goulao, L.; Oliveira, C. Cell wall modifications during fruit ripening: When a fruit is not the fruit. Trends Food Sci. Technol. 2008, 19, 4–25. [Google Scholar] [CrossRef]
- Bosch, M.; Hepler, P.K. Pectin methylesterases and pectin dynamics in pollen tubes. Plant Cell 2005, 17, 3219–3226. [Google Scholar] [CrossRef] [PubMed]
- Goulao, L.F. Pectin de-esterification and fruit softening: Revisiting a classical hypothesis. Stewart Postharvest Rev. 2010, 6, 1–12. [Google Scholar] [CrossRef]
- Rolle, L.; Giacosa, S.; Gerbi, V.; Bertolino, M.; Novello, V. Varietal comparison of the chemical, physical, and mechanical properties of five colored table grapes. Int. J. Food Prop. 2013, 16, 598–612. [Google Scholar] [CrossRef]
- Sims, C.; Halbrooks, M. Quality comparison of “Orlando Seedless” with “Thompson Seedless” grapes. Proc. Florida Sta. Hort. Soc. 1986, 99, 193–194. [Google Scholar]
- Río Segade, S.; Giacosa, S.; Torchio, F.; de Palma, L.; Novello, V.; Gerbi, V.; Rolle, L. Impact of different advanced ripening stages on berry texture properties of “Red Globe”and “Crimson Seedless” table grape cultivars (Vitis vinifera L.). Sci. Hortic. Amsterdam 2013, 160, 313–319. [Google Scholar]
- Grant, G.T.; Morris, E.R.; Rees, D.A.; Smith, P.J.; Thom, D. Biological interactions between polysaccharides and divalent cations: The egg-box model. FEBS Lett. 1973, 32, 195–198. [Google Scholar] [CrossRef]
- Bush, D.S. Calcium regulation in plant cells and its role in signaling. Annu. Rev. Plant Biol. 1995, 46, 95–122. [Google Scholar] [CrossRef]
- Gilroy, S.; Bethke, P.C.; Jones, R.L. Calcium homeostasis in plants. J. Cell Sci. 1993, 106, 453–461. [Google Scholar] [PubMed]
- Micheli, F. Pectin methylesterases: Cell wall enzymes with important roles in plant physiology. Trends Plant Sci. 2001, 6, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Zoccatelli, G.; Zenoni, S.; Savoi, S.; Dal Santo, S.; Tononi, P.; Zandonà, V.; Dal Cin, A.; Guantieri, V.; Pezzotti, M.; Tornielli, G. Skin pectin metabolism during the postharvest dehydration of berries from three distinct grapevine cultivars. Aust. J. Grape Wine R. 2013, 19, 171–179. [Google Scholar] [CrossRef]
- Botondi, R.; Lodola, L.; Mencarelli, F. Postharvest ethylene treatment affects berry dehydration, polyphenol and anthocyanin content by increasing the activity of cell wall enzymes in Aleatico wine grape. Eur. Food Res. Technol. 2011, 232, 679–685. [Google Scholar] [CrossRef]
- Balic, I.; Ejsmentewicz, T.; Sanhueza, D.; Silva, C.; Peredo, T.; Olmedo, P.; Barros, M.; Verdonk, J.C.; Paredes, R.; Meneses, C. Biochemical and physiological study of the firmness of table grape berries. Postharvest Biol. Technol. 2014, 93, 15–23. [Google Scholar] [CrossRef]
- Wei, J.; Ma, F.; Shi, S.; Qi, X.; Zhu, X.; Yuan, J. Changes and postharvest regulation of activity and gene expression of enzymes related to cell wall degradation in ripening apple fruit. Postharvest Biol. Technol. 2010, 56, 147–154. [Google Scholar] [CrossRef]
- Zhou, R.; Li, Y.; Yan, L.; Xie, J. Effect of edible coatings on enzymes, cell-membrane integrity, and cell-wall constituents in relation to brittleness and firmness of Huanghua pears (Pyrus pyrifolia Nakai, cv. Huanghua) during storage. Food Chem. 2011, 124, 569–575. [Google Scholar] [CrossRef]
- Cabanne, C.; Donèche, B. Changes in polygalacturonase activity and calcium content during ripening of grape berries. Am. J. Enol. Vitic. 2001, 52, 331–335. [Google Scholar]
- Buescher, R.; Hudson, J.; Adams, J. Inhibition of polygalacturonase softening of cucumber pickles by calcium chloride. J. Food Sci. 1979, 44, 1786–1787. [Google Scholar] [CrossRef]
- Varela, P.; Salvador, A.; Fiszman, S. Changes in apple tissue with storage time: Rheological, textural and microstructural analyses. J. Food Eng. 2007, 78, 622–629. [Google Scholar] [CrossRef]
- Varela, P.; Salvador, A.; Hernando, I.; Pérez‐Munuera, I.; Quiles, A.; Ángeles Lluch, M.A.; Fiszman, S.M. Eating quality of “Flor de Invierno”pears: Chemical and structural aspects. Int. J. Food Sci. Tech. 2007, 42, 1052–1058. [Google Scholar] [CrossRef]
- Salvador, A.; Sanz, T.; Fiszman, S. Changes in colour and texture and their relationship with eating quality during storage of two different dessert bananas. Postharvest Biol. Technol. 2007, 43, 319–325. [Google Scholar] [CrossRef]
- Azeredo, H.; Brito, E.S.; Moreira, G.E.; Farias, V.L.; Bruno, L.M. Effect of drying and storage time on the physico‐chemical properties of mango leathers. Int. J. Food Sci. Technol. 2006, 41, 635–638. [Google Scholar] [CrossRef]
- Lefever, G.; Vieuille, M.; Delage, N.; Monteclerc, J.; Bompeix, G. Characterization of cell wall enzyme activities, pectin composition, and technological criteria of strawberry cultivars (Fragaria× ananassa Duch). J. Food Sci. 2004, 69, 221–226. [Google Scholar]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Goubet, F.; Morriswood, B.; Dupree, P. Analysis of methylated and unmethylated polygalacturonic acid structure by polysaccharide analysis using carbohydrate gel electrophoresis. Anal. Biochem. 2003, 321, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Deytieux-Belleau, C.; Vallet, A.; Doneche, B.; Geny, L. Pectin methylesterase and polygalacturonase in the developing grape skin. Plant Physiol. Biochem. 2008, 46, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, A.E.; Austin, P.J. Continuous spectrophotometric assay for plant pectin methyl esterase. J. Agric. Food Chem. 1986, 34, 440–444. [Google Scholar] [CrossRef]
- Lohani, S.; Trivedi, P.K.; Nath, P. Changes in activities of cell wall hydrolases during ethylene-induced ripening in banana: Effect of 1-MCP, ABA and IAA. Postharvest Biol. Technol. 2004, 31, 119–126. [Google Scholar] [CrossRef]
- The R Project for Statistical Computing. Available online: http://www.R-project.org (accessed on 18 February 2015).
- Sample Availability: No available.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ejsmentewicz, T.; Balic, I.; Sanhueza, D.; Barria, R.; Meneses, C.; Orellana, A.; Prieto, H.; Defilippi, B.G.; Campos-Vargas, R. Comparative Study of Two Table Grape Varieties with Contrasting Texture during Cold Storage. Molecules 2015, 20, 3667-3680. https://doi.org/10.3390/molecules20033667
Ejsmentewicz T, Balic I, Sanhueza D, Barria R, Meneses C, Orellana A, Prieto H, Defilippi BG, Campos-Vargas R. Comparative Study of Two Table Grape Varieties with Contrasting Texture during Cold Storage. Molecules. 2015; 20(3):3667-3680. https://doi.org/10.3390/molecules20033667
Chicago/Turabian StyleEjsmentewicz, Troy, Iván Balic, Dayan Sanhueza, Romina Barria, Claudio Meneses, Ariel Orellana, Humberto Prieto, Bruno G. Defilippi, and Reinaldo Campos-Vargas. 2015. "Comparative Study of Two Table Grape Varieties with Contrasting Texture during Cold Storage" Molecules 20, no. 3: 3667-3680. https://doi.org/10.3390/molecules20033667