
The Influence of Sesquiterpenes from Myrica rubra on the Antiproliferative and Pro-Oxidative Effects of Doxorubicin and Its Accumulation in Cancer Cells

 , ,
, ,
Abstract
:
1. Introduction

2. Results and Discussion
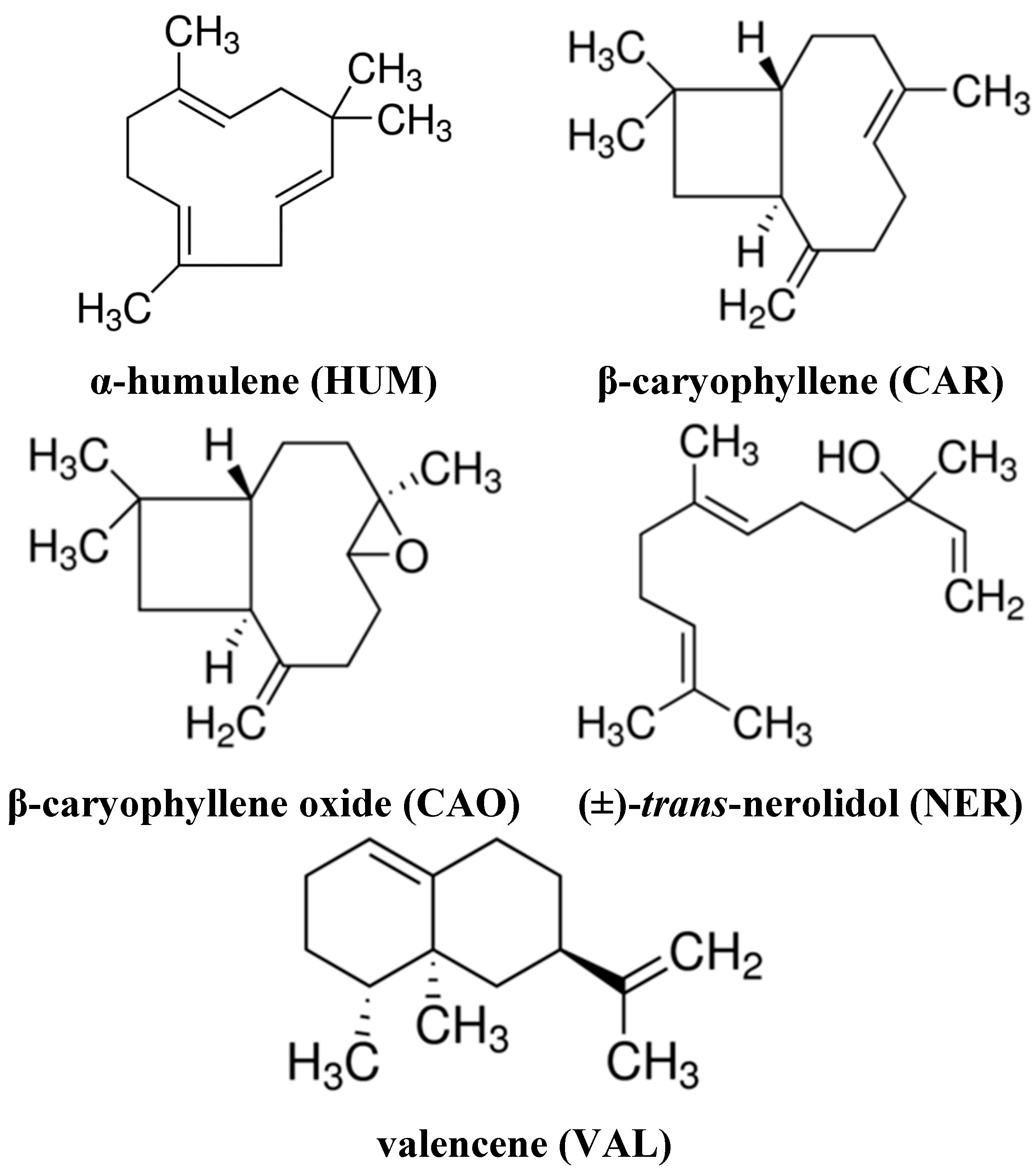
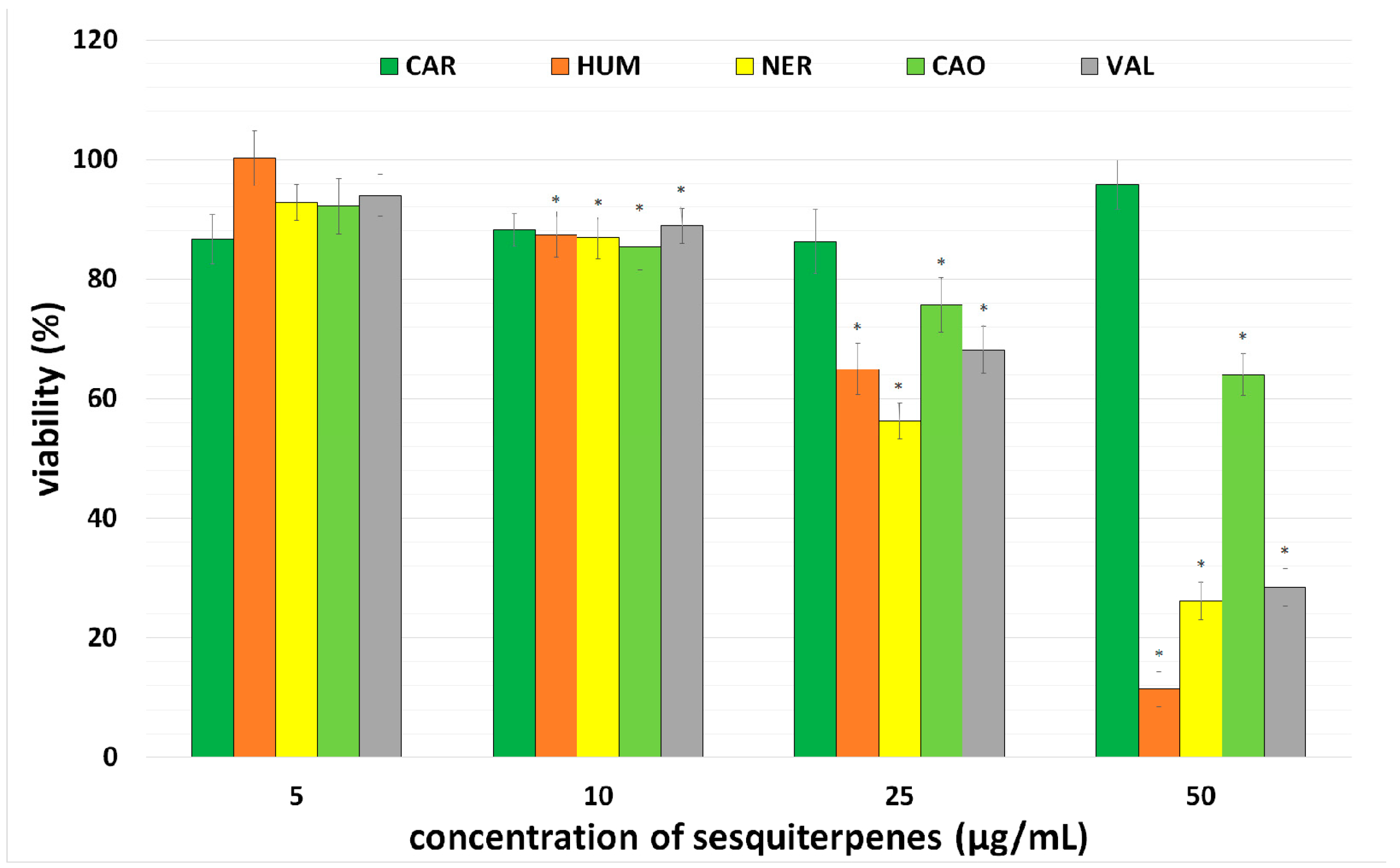
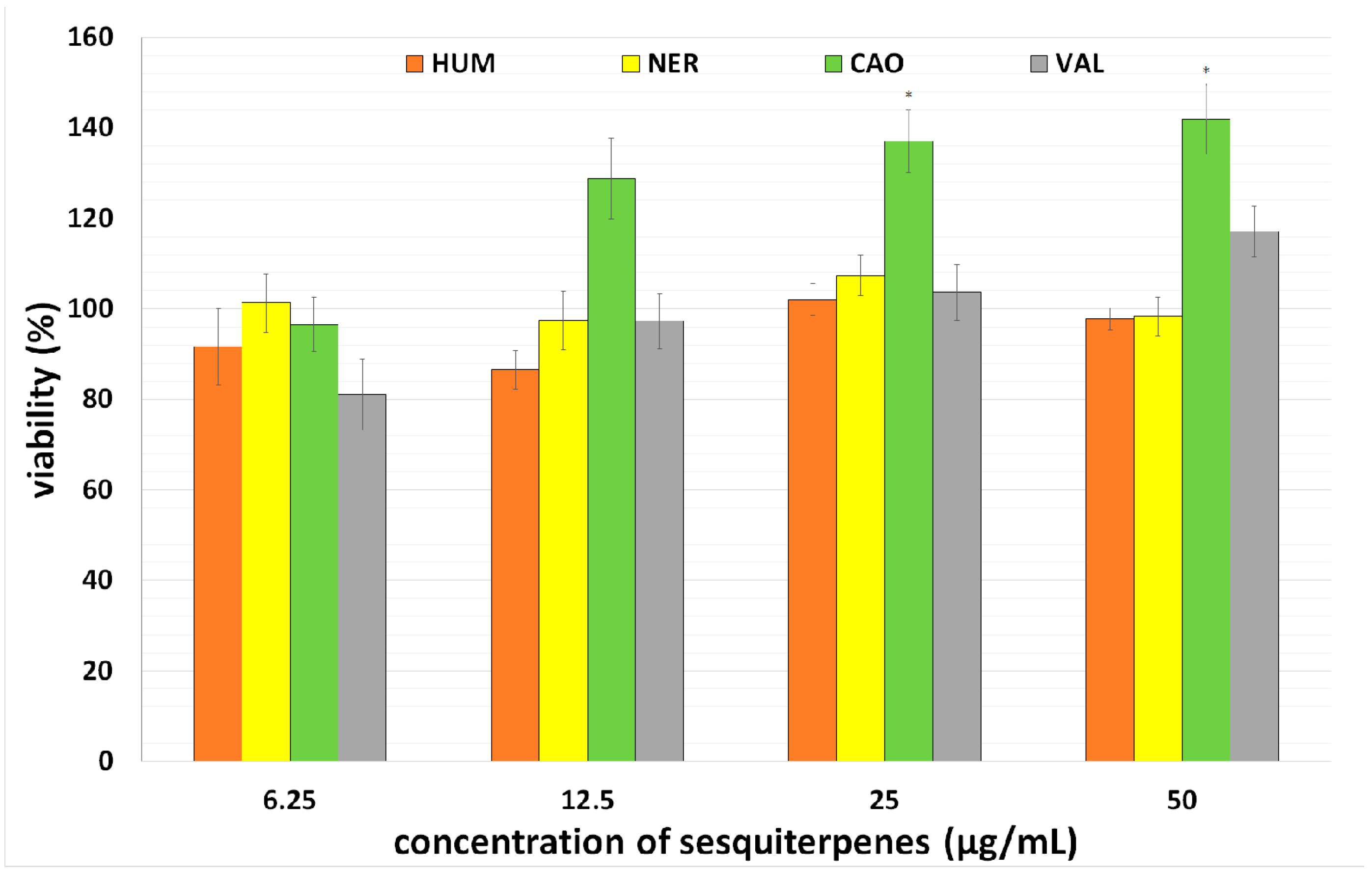
2.1. Effect of Individual Sesquiterpenes on Cell Proliferation/Viability


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (µg/mL) | |
|---|---|
| CAR | - |
| HUM | 24.4 ± 2.4 |
| VAL | 38.1 ± 2.4 |
| CAO | 57.7 ± 3.9 |
| NER | 28.7 ± 2.5 |

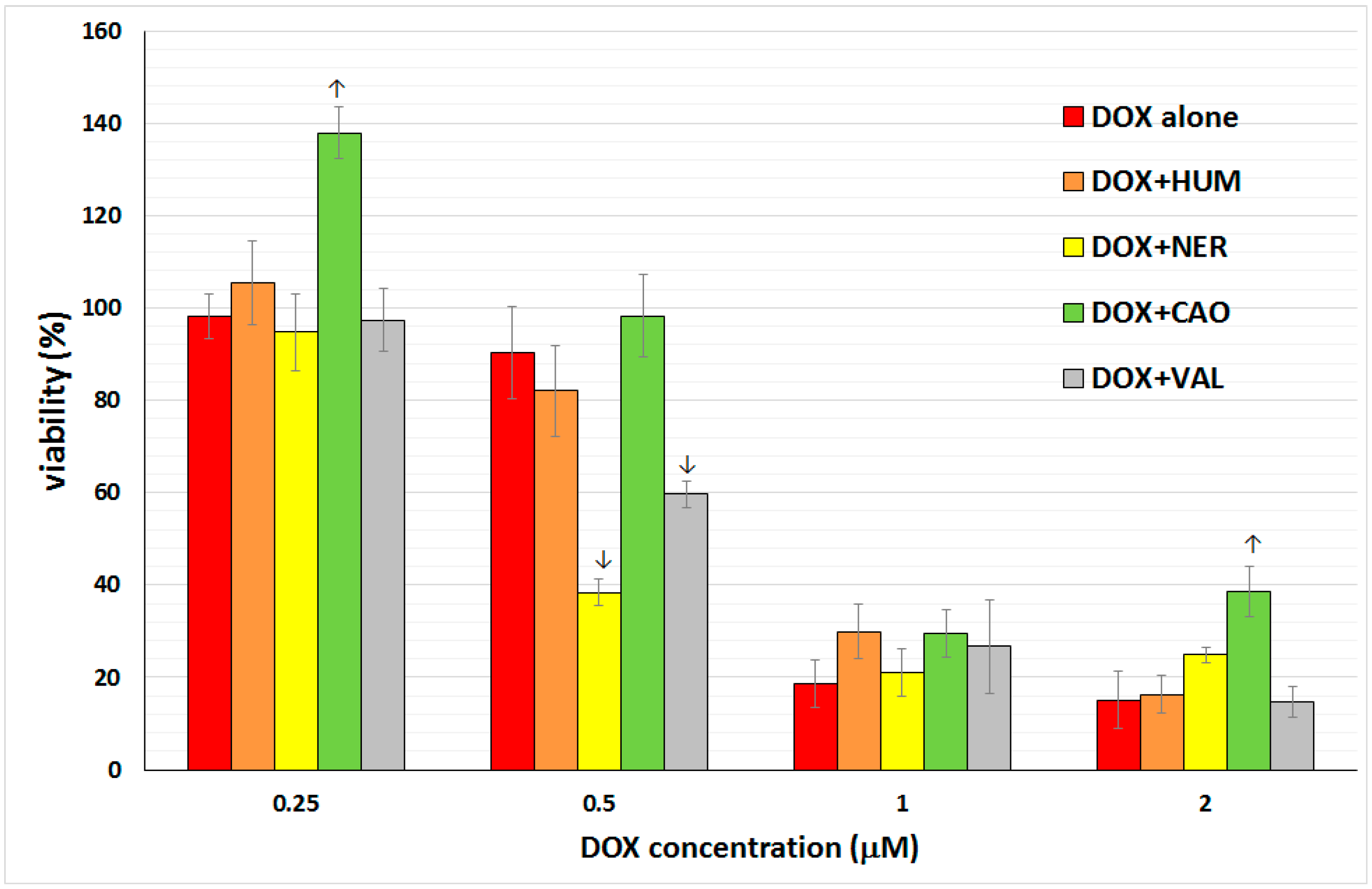
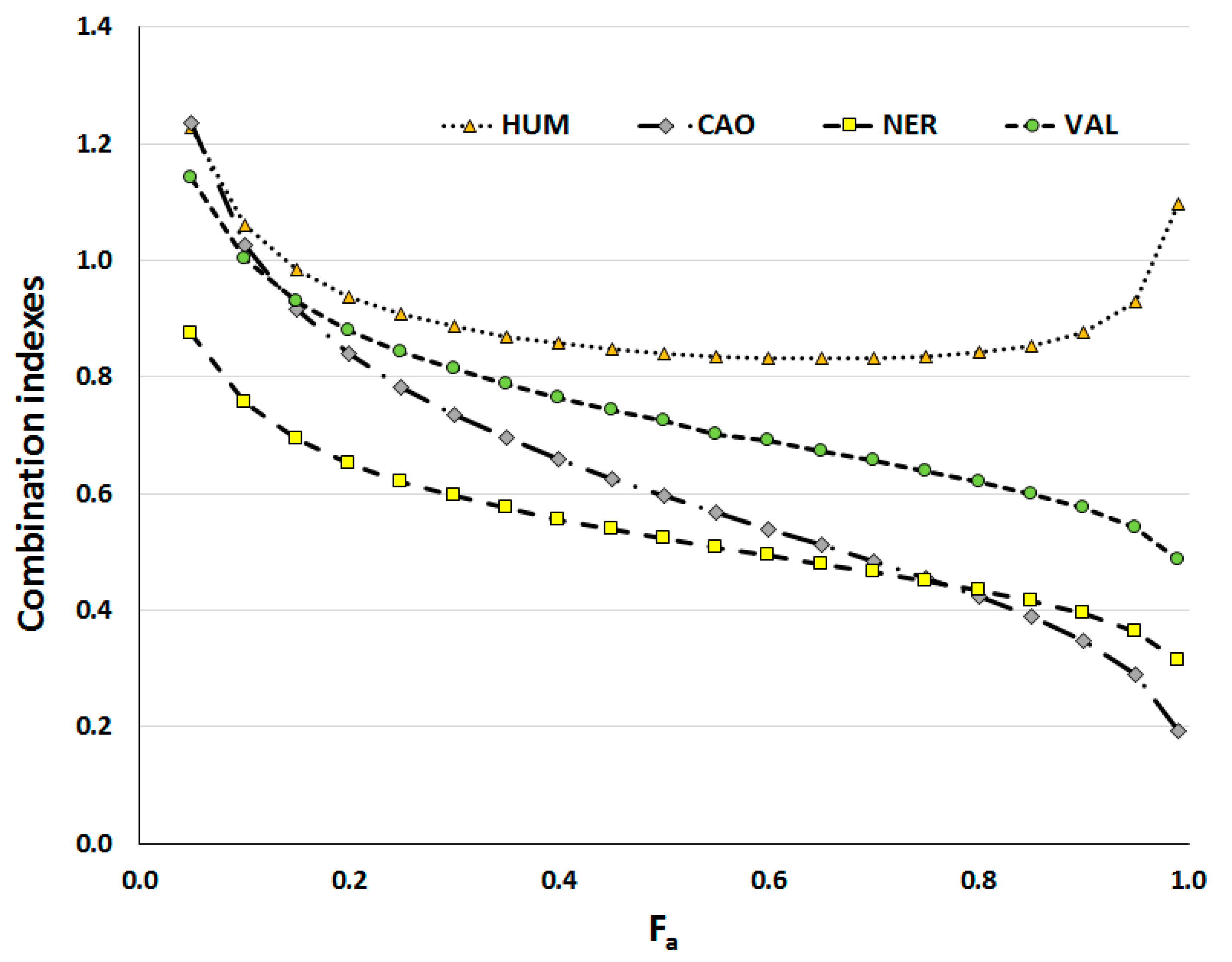
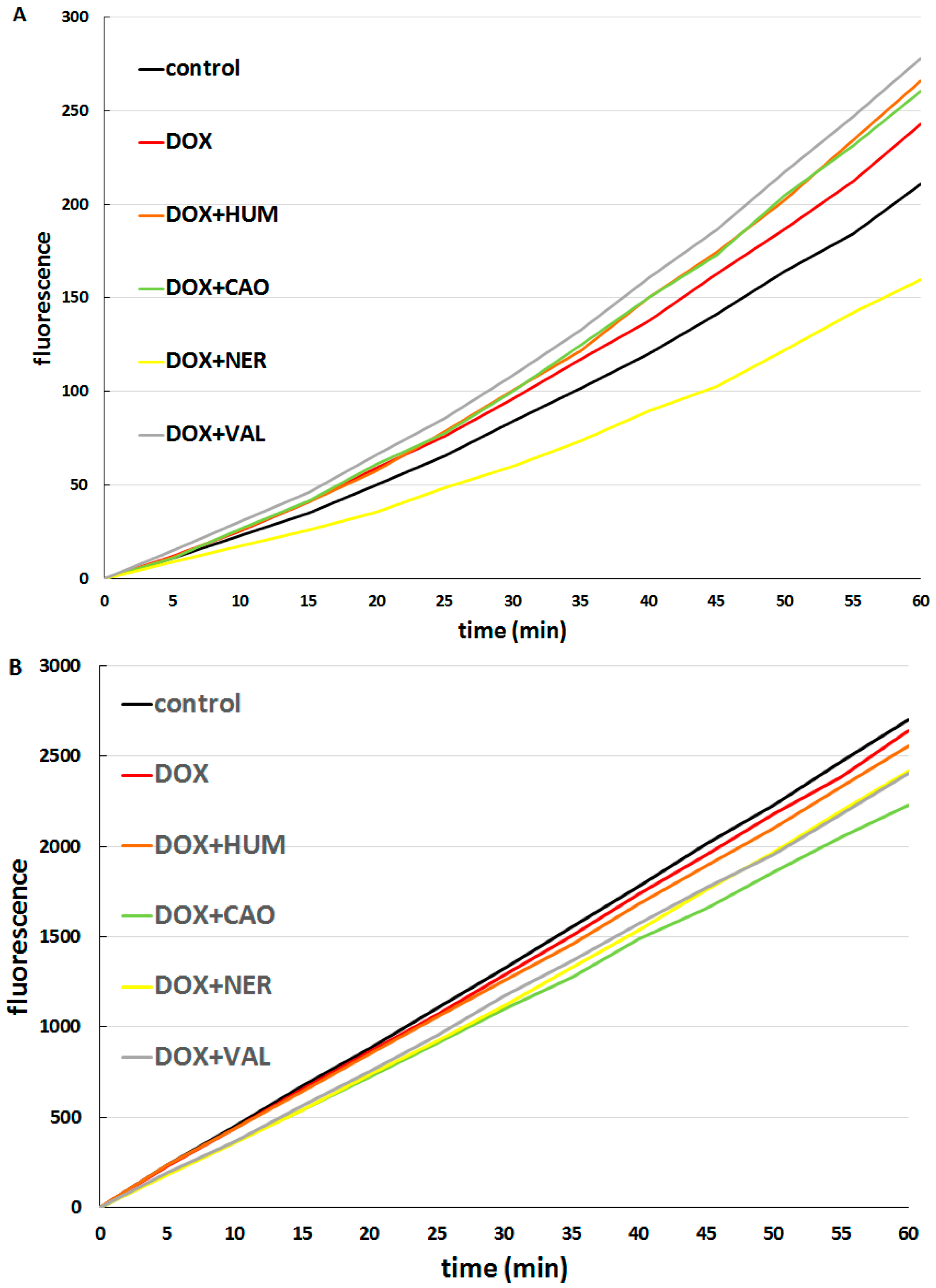
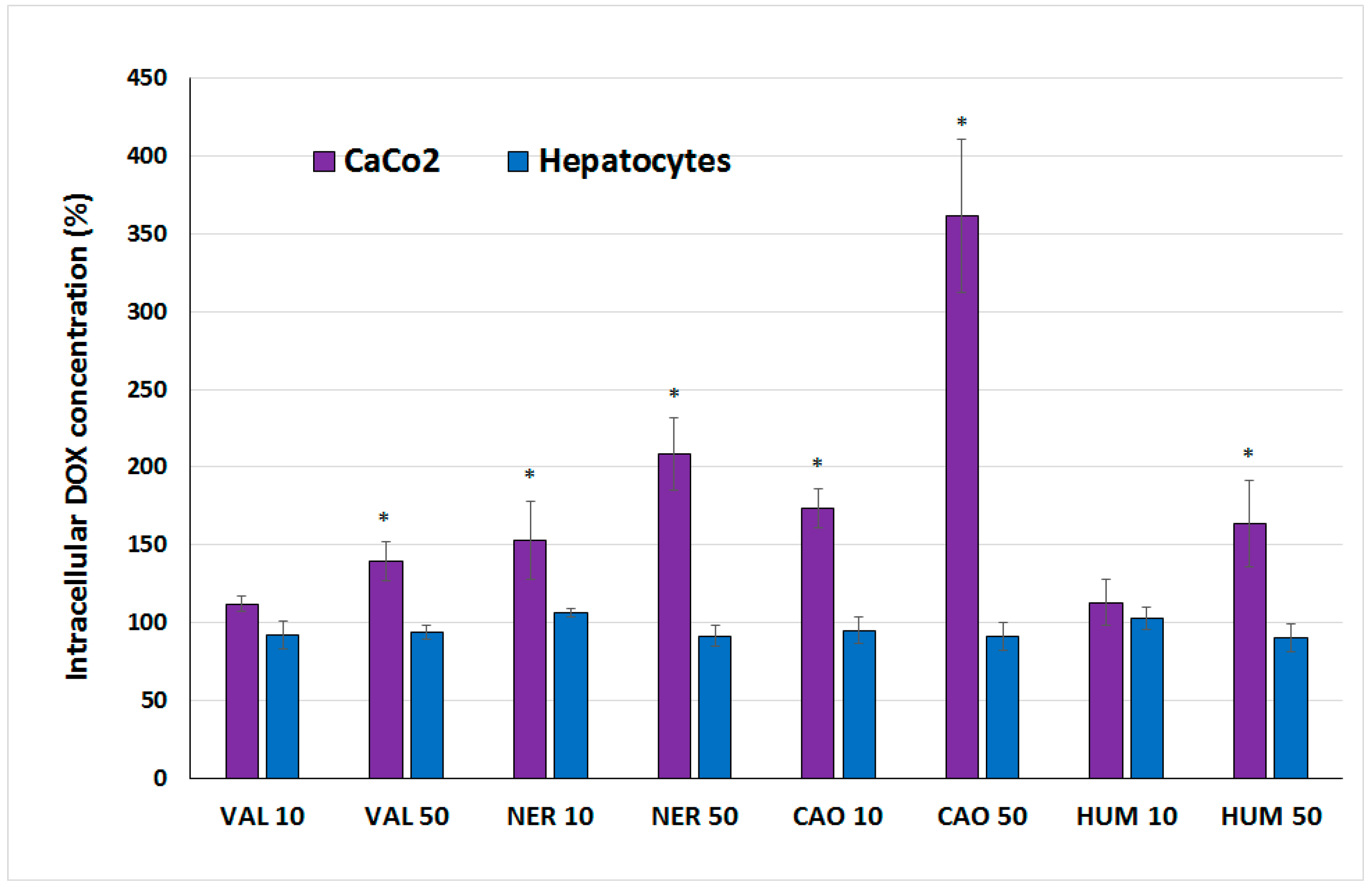
2.2. Effect of Sesquiterpenes in Combinations with Doxorubicin





3. Experimental Section
3.1. Chemicals and Reagents
3.2. Cancer Cell Culture
3.3. Preparation of Primary Culture of Rat Hepatocytes
3.4. Tests of Cell Viability
3.4.1. The Neutral Red Uptake (NRU) Test
3.4.2. The MTT Assay
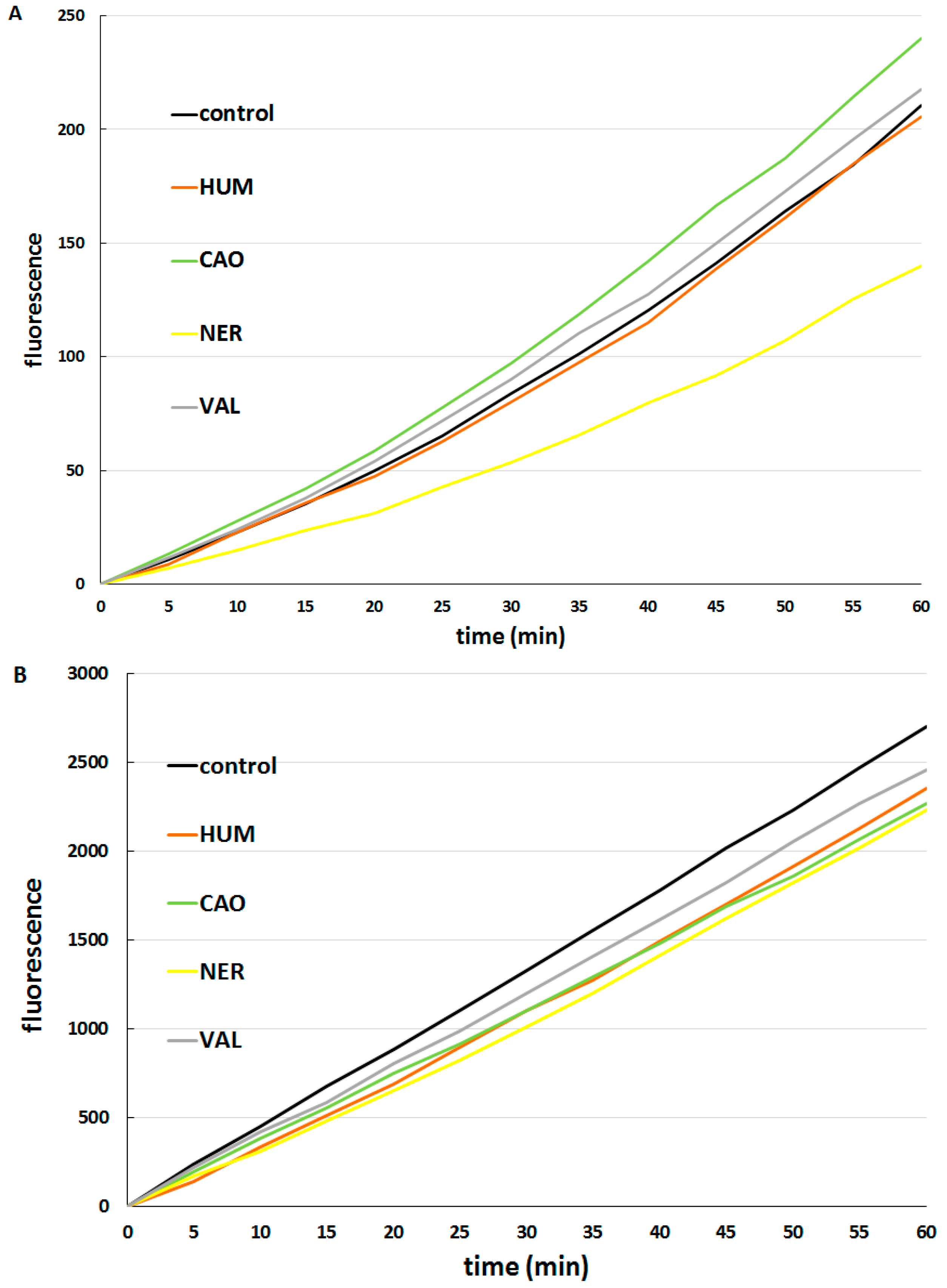
3.5. Determination of Intracellular Reactive Oxygen Species (ROS) Formation
3.6. Test of DOX Accumulation in Cells
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Merfort, I. Perspectives on Sesquiterpene Lactones in Inflammation and Cancer. Curr. Drug Targets 2011, 12, 1560–1573. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Lehtonen, M.; Suuronen, T.; Kaarniranta, K.; Huuskonen, J. Terpenoids: Natural inhibitors of NF-kappa B signaling with anti-inflammatory and anticancer potential. Cell. Mol. Life Sci. 2008, 65, 2979–2999. [Google Scholar] [CrossRef] [PubMed]
- Bartikova, H.; Hanusova, V.; Skalova, L.; Ambroz, M.; Bousova, I. Antioxidant, Pro-Oxidant and Other Biological Activities of Sesquiterpenes. Curr. Top. Med. Chem. 2014, 14, 2478–2494. [Google Scholar] [CrossRef] [PubMed]
- Pajak, B.; Orzechowski, A.; Gajkowska, B. Molecular basis of parthenolide-dependent proapoptotic activity in cancer cells. Folia Histochem. Cytobiol. 2008, 46, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, Y.; Gupta, V.K.; Jaitak, V. Anticancer activity of essential oils: A review. J. Sci. Food Agric. 2013, 93, 3643–3653. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Lu, J.J.; Huang, M.Q.; Bao, J.L.; Chen, X.P.; Wang, Y.T. Terpenoids: Natural products for cancer therapy. Expert Opin. Investig. Drugs 2012, 21, 1801–1818. [Google Scholar] [CrossRef] [PubMed]
- Legault, J.; Pichette, A. Potentiating effect of β-caryophyllene on anticancer activity of α-humulene, isocaryophyllene and paclitaxel. J. Pharm. Pharmacol. 2007, 59, 1643–1647. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Cho, S.K.; Kim, K.D.; Nam, D.; Chung, W.S.; Jang, H.J.; Lee, S.G.; Shim, B.S.; Sethi, G.; Ahn, K.S. β-Caryophyllene oxide potentiates TNF α-induced apoptosis and inhibits invasion through down-modulation of NF-kappa B-regulated gene products. Apoptosis 2014, 19, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Abdul, A.B.; Abdelwahab, S.I.; Jalinas, J.B.; Al-Zubairi, A.S.; Taha, M.M.E. Combination of Zerumbone and Cisplatin to Treat Cervical Intraepithelial Neoplasia in Female BALB/c Mice. Int. J. Gynecol. Cancer 2009, 19, 1004–1010. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.S.; Dang, Y.Y.; Chen, X.P.; Lu, J.J.; Wang, Y.T. Furanodiene Presents Synergistic Anti-proliferative Activity With Paclitaxel via Altering Cell Cycle and Integrin Signaling in 95-D Lung Cancer Cells. Phytother. Res. 2014, 28, 296–299. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.F.; Li, Y.B.; Wang, S.P.; Tan, W.; Chen, X.P.; Chen, M.W.; Wang, Y.T. Furanodiene enhances tamoxifen-induced growth inhibitory activity of ERa-positive breast cancer cells in a PPAR gamma independent manner. J. Cell. Biochem. 2012, 113, 2643–2651. [Google Scholar] [CrossRef] [PubMed]
- Adorjan, B.; Buchbauer, G. Biological properties of essential oils: An updated review. Flavour Fragr. J. 2010, 25, 407–426. [Google Scholar] [CrossRef]
- Bansal, A.; Moriarity, D.M.; Takaku, S.; Setzer, W.N. Chemical composition and cytotoxic activity of the leaf essential oil of Ocotea tonduzii from Monteverde, Costa Rica. Nat. Prod. Commun. 2007, 2, 781–784. [Google Scholar]
- Erdogan, A.; Ozkan, A. Effects of Thymus revolutus Celak essential oil and its two major components on Hep G2 cells membrane. Biologia 2013, 68, 105–111. [Google Scholar] [CrossRef]
- Raut, J.S.; Karuppayil, S.M. A status review on the medicinal properties of essential oils. Ind. Crops Prod. 2014, 62, 250–264. [Google Scholar] [CrossRef]
- Langhansova, L.; Hanusova, V.; Rezek, J.; Stohanslova, B.; Ambroz, M.; Kralova, V.; Vanek, T.; Lou, J.D.; Yun, Z.L.; Yang, J.; et al. Essential oil from Myrica rubra leaves inhibits cancer cell proliferation and induces apoptosis in several human intestinal lines. Ind. Crops Prod. 2014, 59, 20–26. [Google Scholar] [CrossRef]
- Kang, W.; Li, Y.; Xu, Y.; Jiang, W.; Tao, Y. Characterization of Aroma Compounds in Chinese Bayberry (Myrica rubra Sieb. et Zucc.) by Gas Chromatography Mass Spectrometry (GC-MS) and Olfactometry (GC-O). J. Food Sci. 2012, 77, C1030–C1035. [Google Scholar] [CrossRef] [PubMed]
- Hanusova, V.; Bousova, I.; Skalova, L. Possibilities to increase the effectiveness of doxorubicin in cancer cells killing. Drug Metab. Rev. 2011, 43, 540–557. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.D.; Huang, H.Z.; Xu, C.J.; Li, X.; Chen, K.S. Biological Activities of Extracts from Chinese Bayberry (Myrica rubra Sieb. et Zucc.): A Review. Plant Foods Hum. Nutr. 2013, 68, 97–106. [Google Scholar] [CrossRef] [PubMed]
- El Hadri, A.; del Rio, M.A.G.; Sanz, J.; Coloma, A.G.; Idaomar, M.; Ozonas, B.R.; Gonzalez, J.B.; Reus, M.I.S. Cytotoxic activity of α-humulene and trans-caryophyllene from Salvia officinalis in animal and human tumor cells. Anal. Real Acad. Nac. Farm. 2010, 76, 343–356. [Google Scholar]
- Legault, J.; Dahl, W.; Debiton, E.; Pichette, A.; Madelmont, J.C. Antitumor activity of balsam fir oil: Production of reactive oxygen species induced by α-humulene as possible mechanism of action. Planta Med. 2003, 69, 402–407. [Google Scholar] [PubMed]
- Sylvestre, M.; Pichette, A.; Lavoie, S.; Longtin, A.; Legault, J. Composition and cytotoxic activity of the leaf essential oil of Comptonia peregrina (L.) Coulter. Phytother. Res. 2007, 21, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Chen, Q.; Liu, Y.; Zhou, X.; Wang, X. Isolation and Biological Activities of Decanal, Linalool, Valencene, and Octanal from Sweet Orange Oil. J. Food Sci. 2012, 77, C1156–C1161. [Google Scholar] [CrossRef] [PubMed]
- Jun, N.J.; Mosaddik, A.; Moon, J.Y.; Jang, K.C.; Lee, D.S.; Ahn, K.S.; Cho, S.K. Cytotoxic Activity of β-Caryophyllene Oxide Isolated from Jeju Guava (Psidium cattleianum Sabine) Leaf. Rec. Natl. Prod. 2011, 5, 242–246. [Google Scholar]
- Park, K.R.; Nam, D.; Yun, H.M.; Lee, S.G.; Jang, H.J.; Sethi, G.; Cho, S.K.; Ahn, K.S. β-Caryophyllene oxide inhibits growth and induces apoptosis through the suppression of PI3K/AKT/mTOR/S6K1 pathways and ROS-mediated MAPKs activation. Cancer Lett. 2011, 312, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Di Sotto, A.; Maffei, F.; Hrelia, P.; Castelli, F.; Sarpietro, M.G.; Mazzanti, G. Genotoxicity assessment of β-caryophyllene oxide. Regul. Toxicol. Pharmacol. 2013, 66, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.J.; Kim, H.J.; Chun, H.S. Quantitative structure-activity relationship (QSAR) for neuroprotective activity of terpenoids. Life Sci. 2007, 80, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Krishnaiah, Y.S.R.; Al-Saidan, S.M.; Jayaram, B. Effect of nerodilol, carvone and anethole on the in vitro transdermal delivery of selegiline hydrochloride. Pharmazie 2006, 61, 46–53. [Google Scholar] [PubMed]
- Saeedi, M.; Morteza-Semnani, K. Penetration-Enhancing Effect of the Essential Oil and Methanolic Extract of Eryngium bungei on Percutaneous Absorption of Piroxicam through Rat Skin. J. Essent. Oil Bear. Plants 2009, 12, 728–741. [Google Scholar] [CrossRef]
- Williams, A.C.; Barry, B.W. Penetration enhancers. Adv. Drug Deliv. Rev. 2012, 64, 128–137. [Google Scholar] [CrossRef]
- Chou, T.C. Drug Combination Studies and Their Synergy Quantification Using the Chou-Talalay Method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.S.; Torchilin, V.P. Hormetic/Cytotoxic Effects of Nigella sativa Seed Alcoholic and Aqueous Extracts on MCF-7 Breast Cancer Cells Alone or in Combination with Doxorubicin. Cell Biochem. Biophys. 2013, 66, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Riganti, C.; Doublier, S.; Costamagna, C.; Aldieri, E.; Pescarmona, G.; Ghigo, D.; Bosia, A. Activation of nuclear factor-kappa B pathway by Simvastatin and RhoA silencing increases doxorubicin cytotoxicity in human colon cancer HT29 cells. Mol. Pharmacol. 2008, 74, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, M.; Szulawska-Mroczek, A.; Hartman, M.L.; Nejc, D.; Czyz, M. Parthenolide Complements the Cell Death-inducing Activity of Doxorubicin in Melanoma Cells. Anticancer Res. 2013, 33, 3205–3212. [Google Scholar] [PubMed]
- Martin-Cordero, C.; Leon-Gonzalez, A.J.; Calderon-Montano, J.M.; Burgos-Moron, E.; Lopez-Lazaro, M. Pro-Oxidant Natural Products as Anticancer Agents. Curr. Drug Targets 2012, 13, 1006–1028. [Google Scholar] [CrossRef] [PubMed]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fang, P.; Mai, J.; Choi, E.T.; Wang, H.; Yang, X.F. Targeting mitochondrial reactive oxygen species as novel therapy for inflammatory diseases and cancers. J. Hematol. Oncol. 2013, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, S.; D’Abrosca, B.; Golino, A.; Mastellone, C.; Piccolella, S.; Fiorentino, A.; Monaco, P. Antioxidant evaluation of polyhydroxylated nerolidols from redroot pigweed (Amaranthus retroflexus) leaves. Lwt-Food Sci. Technol. 2008, 41, 1665–1671. [Google Scholar] [CrossRef]
- Vinholes, J.; Goncalves, P.; Martel, F.; Coimbra, M.A.; Rocha, S.M. Assessment of the antioxidant and antiproliferative effects of sesquiterpenic compounds in in vitro CaCo-2 cell models. Food Chem. 2014, 156, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Joseph, J.A. Quantifying cellular oxidative stress by dichlorofluorescein assay using microplate reader. Free Radic. Biol. Med. 1999, 27, 612–616. [Google Scholar] [CrossRef]
- Skarka, A.; Skarydova, L.; Stambergova, H.; Wsol, V. Anthracyclines and their metabolism in human liver microsomes and the participation of the new microsomal carbonyl reductase. Chem. Biol. Interact. 2011, 191, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ambrož, M.; Boušová, I.; Skarka, A.; Hanušová, V.; Králová, V.; Matoušková, P.; Szotáková, B.; Skálová, L. The Influence of Sesquiterpenes from Myrica rubra on the Antiproliferative and Pro-Oxidative Effects of Doxorubicin and Its Accumulation in Cancer Cells. Molecules 2015, 20, 15343-15358. https://doi.org/10.3390/molecules200815343
Ambrož M, Boušová I, Skarka A, Hanušová V, Králová V, Matoušková P, Szotáková B, Skálová L. The Influence of Sesquiterpenes from Myrica rubra on the Antiproliferative and Pro-Oxidative Effects of Doxorubicin and Its Accumulation in Cancer Cells. Molecules. 2015; 20(8):15343-15358. https://doi.org/10.3390/molecules200815343
Chicago/Turabian StyleAmbrož, Martin, Iva Boušová, Adam Skarka, Veronika Hanušová, Věra Králová, Petra Matoušková, Barbora Szotáková, and Lenka Skálová. 2015. "The Influence of Sesquiterpenes from Myrica rubra on the Antiproliferative and Pro-Oxidative Effects of Doxorubicin and Its Accumulation in Cancer Cells" Molecules 20, no. 8: 15343-15358. https://doi.org/10.3390/molecules200815343
APA StyleAmbrož, M., Boušová, I., Skarka, A., Hanušová, V., Králová, V., Matoušková, P., Szotáková, B., & Skálová, L. (2015). The Influence of Sesquiterpenes from Myrica rubra on the Antiproliferative and Pro-Oxidative Effects of Doxorubicin and Its Accumulation in Cancer Cells. Molecules, 20(8), 15343-15358. https://doi.org/10.3390/molecules200815343