
Effect of Donepezil, Tacrine, Galantamine and Rivastigmine on Acetylcholinesterase Inhibition in Dugesia tigrina

 ,
,
Abstract
:1. Introduction
2. Results and Discussion

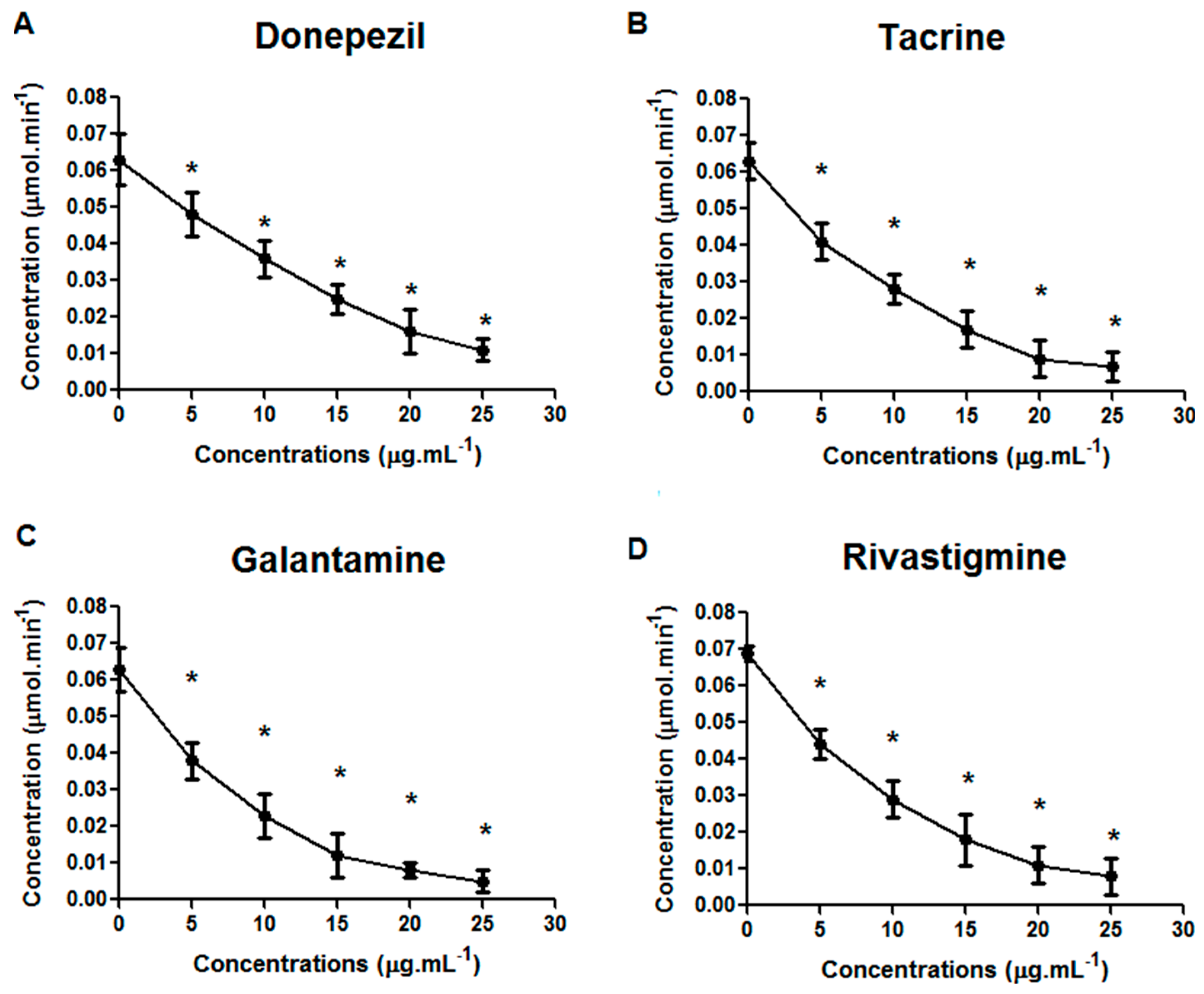{kind=link}
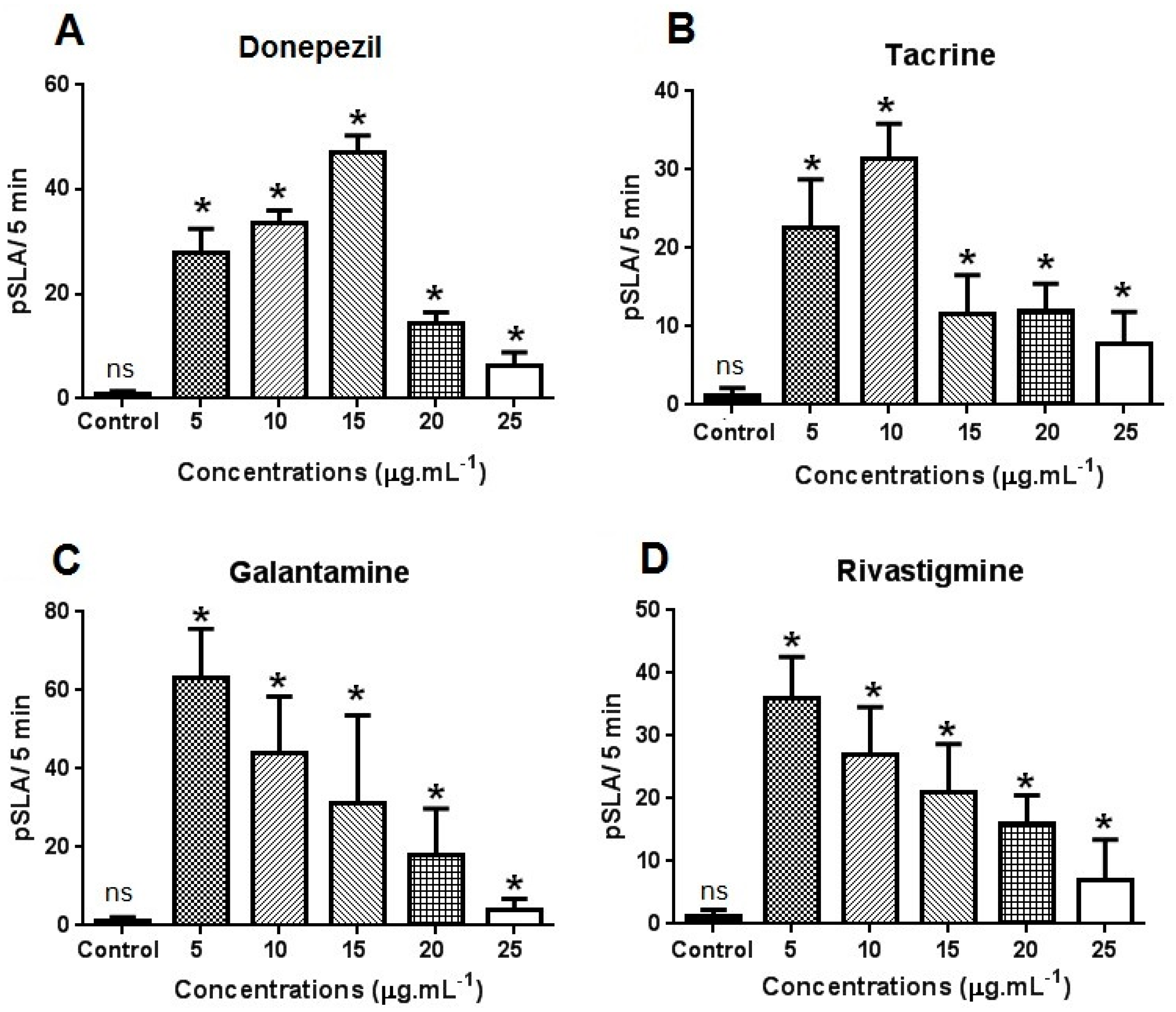{kind=link}
{kind=link}
| Mortality | ||||||||
|---|---|---|---|---|---|---|---|---|
| Drugs | Control | 5 μg·mL−1 | 10 μg·mL−1 | 15 μg·mL−1 | 20 μg·mL−1 | 25 μg·mL−1 | LC50 | Confidence Interval |
| Donepezil | 0 | 1 ns | 3 ns | 4 * | 5 * | 8 * | >25 | 20–45 |
| Tacrine | 0 | 2 ns | 2 ns | 3 ns | 6 * | 10 * | ≤21.2 | 17.5–31.7 |
| Rivastigmine | 0 | 1 ns | 3 ns | 5 * | 7 * | 12 * | ≤17.4 | 15–21 |
| Galantamine | 0 | 4 * | 6 * | 12 * | 15 * | 15 * | ≤8.3 | 5.5–12.5 |



3. Experimental Section
3.1. Collection and Acclimation of D. tigrina
3.2. Evaluation of Mortality
3.3. Preparation of the Enzymatic Extract
3.4. Determination of Acetylcholinesterase
3.5. Measures of Seizure-Like Activity
3.6. Motility Measurement (pLMV)
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Roberson, M.R.; Harrell, L.E. Cholinergic activity and amyloid precursor protein metabolism. Brain Res. Rev. 1997, 25, 50–69. [Google Scholar] [CrossRef]
- McGeer, E.G.; McGeer, P.L. Clinically tested drugs for Alzheimer’s disease. Expert Opin. Investig. Drugs 2003, 12, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Ademiluyi, A.O.; Akinyemi, A.J. Inhibition of acetylcholinesterase activities and some pro-oxidant induced lipid peroxidation in rat brain by two varieties of ginger (Zingiber officinale). Exp. Toxicol. Pathol. 2012, 64, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Mejri, N.; Said, N.M.; Guizani, S.; Essouissi, I.; Saidi, M. Preliminary studies of acetylcholinesterase activity in the rat brain using N-phenylferrocenecarboxamide labelled by the technetium-99. Nucl. Med. Biol. 2013, 40, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Ozarowski, M.; Mikolajczak, P.L.; Bogacz, A.; Gryszczynska, A.; Kujawska, M.; Jodynis-Liebert, J.; Piasecka, A.; Napieczynska, H.; Szulc, M.; Kujawski, R.; et al. Rosmarinus officinalis L. leaf extract improves memory impairment and affects acetylcholinesterase and butyrylcholinesterase activities in rat brain. Fitoterapia 2013, 91, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kaur, M.; Kukreja, H.; Chugh, R.; Silakari, O.; Singh, D. Acetylcholinesterase inhibitors as Alzheimer therapy: From nerve toxins to neuroprotection. Eur. J. Med. Chem. 2013, 70, 165–188. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.N.; Han, H.; Hu, X.D.; Feng, G.F.; Qian, Y.H. The effects of perindopril on cognitive impairment induced by d-galactose and aluminum trichloride via inhibition of acetylcholinesterase activity and oxidative stress. Pharmacol. Biochem. Behav. 2013, 114–115, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Pagán, O.R.; Deats, S.; Baker, D.; Montgomery, E.; Wilk, G.; Tenaglia, M.; Semon, J. Planarians require an intact brain to behaviorally react to cocaine, but not to react to nicotine. Neuroscience 2013, 246, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Cebria, F. Regenerating the central nervous system: How easy for planarians! Dev. Genes Evol. 2007, 217, 733–748. [Google Scholar] [CrossRef] [PubMed]
- Cebria, F.; Nakazawa, M.; Mineta, K.; Ikeo, K.; Gojobori, T.; Agata, K. Dissecting planarian central nervous system regeneration by the expression of neural-specific genes. Dev. Growth Differ. 2002, 44, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Aoki, R.; Wake, H.; Sasaki, H.; Agata, K. Recording and spectrum analysis of the planarian electroencephalogram. Neuroscience 2009, 159, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Farrell, M.S.; Gilmore, K.; Raffa, R.B.; Walker, E.A. Behavioral characterization of serotonergic activation in the flatworm Planaria. Behav. Pharmacol. 2008, 19, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Kitamura, Y.; Umesono, Y.; Takeuchi, K.; Takata, K.; Taniguchi, T.; Agata, K. Identification of glutamic acid decarboxylase gene and distribution of GABAergic nervous system in the planarian Dugesia japonica. Neuroscience 2008, 153, 1103–1114. [Google Scholar] [CrossRef] [PubMed]
- Gentile, L.; Cebria, F.; Bartscherer, K. The planarian flatworm: An in vivo model for stem cell biology and nervous system regeneration. Dis. Model. Mech. 2011, 4, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, L.; Desaer, C. Carbamazepine inhibits distinct chemoconvulsant-induced seizure-like activity in Dugesia tigrina. Pharmacol. Biochem. Behav. 2011, 99, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Forsthoefel, D.J.; James, N.P.; Escobar, D.J.; Stary, J.M.; Vieira, A.P.; Waters, F.A.; Newmark, P.A. An RNAi screen reveals intestinal regulators of branching morphogenesis, differentiation, and stem cell proliferation in planarians. Dev. Cell 2012, 23, 691–704. [Google Scholar] [CrossRef] [PubMed]
- Isolani, M.E.; Pietra, D.; Balestrini, L.; Borghini, A.; Deri, P.; Imbriani, M.; Bianucci, A.M.; Batistoni, R. The in vivo effect of chelidonine on the stem cell system of planarians. Eur. J. Pharmacol. 2012, 686, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Caricati-Neto, A.; D’Angelo L, C.; Reuter, H.; Hyppolito Jurkiewicz, N.; Garcia, A.G.; Jurkiewicz, A. Enhancement of purinergic neurotransmission by galantamine and other acetylcholinesterase inhibitors in the rat vas deferens. Eur. J. Pharmacol. 2004, 503, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Geerts, H.; Guillaumat, P.O.; Grantham, C.; Bode, W.; Anciaux, K.; Sachak, S. Brain levels and acetylcholinesterase inhibition with galantamine and donepezil in rats, mice, and rabbits. Brain Res. 2005, 1033, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Porcel, J.; Montalban, X. Anticholinesterasics in the treatment of cognitive impairment in multiple sclerosis. J. Neurol. Sci. 2006, 245, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Yanagida, T.; Takeuchi, H.; Kitamura, Y.; Takata, K.; Minamino, H.; Shibaike, T.; Tsushima, J.; Kishimoto, K.; Yasui, H.; Taniguchi, T.; et al. Synergistic effect of galantamine on nicotine-induced neuroprotection in hemiparkinsonian rat model. Neurosci. Res. 2008, 62, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Jonckers, E.; van der Linden, A.; Verhoye, M. Functional magnetic resonance imaging in rodents: An unique tool to study in vivo pharmacologic neuromodulation. Curr. Opin. Pharmacol. 2013, 13, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Orhan, I.E.; Senol, F.S.; Ercetin, T.; Kahraman, A.; Celep, F.; Akaydin, G.; Sener, B.; Dogan, M. Assessment of anticholinesterase and antioxidant properties of selected sage (Salvia) species with their total phenol and flavonoid contents. Ind. Crop. Prod. 2013, 41, 21–30. [Google Scholar] [CrossRef]
- Ribeiro, P.; El-Shehabi, F.; Patocka, N. Classical transmitters and their receptors in flatworms. Parasitology 2005, 131, S19–S40. [Google Scholar] [CrossRef] [PubMed]
- Bajgarl, J.; Fusek, J.; Skopec, F. Changes of cholinesterases in the blood and some tissues following administration of tacrin and its two derivatives to rats. Neurochem. Int. 1994, 24, 555–558. [Google Scholar] [CrossRef]
- Myhrer, T.; Enger, S.; Aas, P. Antiparkinson drugs used as prophylactics for nerve agents: Studies of cognitive side effects in rats. Pharmacol. Biochem. Behav. 2008, 89, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Scali, C.; Casamenti, F.; Bellucci, A.; Costagli, C.; Schmidt, B.; Pepeu, G. Effect of subchronic administration of metrifonate, rivastigmine and donepezil on brain acetylcholine in aged F344 rats. J. Neural Transm. 2002, 109, 1067–1080. [Google Scholar]
- Braga, M.F.; Harvey, A.L.; Rowan, E.G. Effects of tacrine, velnacrine (HP029), suronacrine (HP128), and 3,4-diaminopyridine on skeletal neuromuscular transmission in vitro. Br. J. Pharmacol. 1991, 102, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Creeley, C.E.; Wozniak, D.F.; Nardi, A.; Farber, N.B.; Olney, J.W. Donepezil markedly potentiates memantine neurotoxicity in the adult rat brain. Neurobiol. Aging 2008, 29, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.-H.; Yang, Z.-D.; Xue, W.-W.; Sheng, J.; Shi, Y.; Yao, X.-J. Isolation, characterization and acetylcholinesterase inhibitory activity of alkaloids from roots of Stemona sessilifolia. Fitoterapia 2013, 89, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Santos-Bueso, E.; Serrador-Garcia, M.; Garcia-Sanchez, J. [Donepezil as treatment of Charles Bonnet syndrome]. Arch. Soc. Esp. Oftalmol. 2013, 88, 369–370. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, T.; Kaden, B.; Fischer, J.P.; Bickel, U.; Barz, H.; Gusztony, G.; Cervos-Navarro, J.; Kewitz, H. Inhibition of acetylcholinesterase activity in human brain tissue and erythrocytes by galanthamine, physostigrnine and tacrine. Eur. J. Clin. Chem. Clin. Biochem. 1991, 29, 487–492. [Google Scholar] [PubMed]
- Lallement, G.; Denoyer, M.; Collet, A.; Pernot-Marino, I.; Baubichon, D.; Monmaur, P.; Blanchet, G. Changes in hippocampal acetylcholine and glutamate extracellular levels during soman-induced seizures: Influence of septal cholinoceptive cells. Neurosci. Lett. 1992, 139, 104–107. [Google Scholar] [CrossRef]
- McDonough, J.H., Jr.; Shih, T.M. Neuropharmacological mechanisms of nerve agent-induced seizure and neuropathology. Neurosci. Biobehav. Rev. 1997, 21, 559–579. [Google Scholar] [CrossRef]
- Carpentier, P.; Lambrinidis, M.; Blanchet, G. Early dendritic changes in hippocampal pyramidal neurones (field CA1) of rats subjected to acute soman intoxication: A light microscopic study. Brain Res. 1991, 541, 293–299. [Google Scholar] [CrossRef]
- Janowsky, D.S.; Davis, J.M.; Overstreet, D.H. Anticholinesterase (DFP) toxicity antagonism by chronic donepezil: A potential nerve agent treatment. Pharmacol. Biochem. Behav. 2005, 81, 917–922. [Google Scholar] [CrossRef] [PubMed]
- Lagadic-Gossmann, D.; Rissel, M.; Le Bot, M.A.; Guillouzo, A. Toxic effects of tacrine on primary hepatocytes and liver epithelial cells in culture. Cell Biol. Toxicol. 1998, 14, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Torres-Da Matta, J.; Kanaan, S.; Nery da Mata, A. Changes in glycolytic metabolism during the regeneration of planarians Dugesia tigrina (Girard). Comp. Biochem. Physiol. B Comp. Biochem. 1989, 93, 391–396. [Google Scholar] [CrossRef]
- Buttarelli, F.R.; Pontieri, F.E.; Margotta, V.; Palladini, G. Acetylcholine/dopamine interaction in planaria. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2000, 125, 225–231. [Google Scholar] [CrossRef]
- Wu, H.P.; Persinger, M.A. Increased mobility and stem-cell proliferation rate in Dugesia tigrina induced by 880nm light emitting diode. J. Photochem. Photobiol. 2011, 102, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Rawls, S.M.; Thomas, T.; Adeola, M.; Patil, T.; Raymondi, N.; Poles, A.; Loo, M.; Raffa, R.B. Topiramate antagonizes NMDA- and AMPA-induced seizure-like activity in planarians. Pharmacol. Biochem. Behav. 2009, 93, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Raffa, R.B.; Cavallo, F.; Capasso, A. Flumazenil-sensitive dose-related physical dependence in planarians produced by two benzodiazepine and one non-benzodiazepine benzodiazepine-receptor agonists. Eur. J. Pharmacol. 2007, 564, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.C.; Alvares, E.P.; Gama, P.; Silveira, M. Serotonin in the nervous system of the head region of the land planarian Bipalium kewense. Tissue Cell 2003, 35, 479–486. [Google Scholar] [CrossRef]
- Raffa, R.B.; Holland, L.J.; Schulingkamp, R.J. Quantitative assessment of dopamine D2 antagonist activity using invertebrate (Planaria) locomotion as a functional endpoint. J. Pharmacol. Toxicol. Methods 2001, 45, 223–226. [Google Scholar] [CrossRef]
- Rawls, S.M.; Patil, T.; Yuvasheva, E.; Raffa, R.B. First evidence that drugs of abuse produce behavioral sensitization and cross-sensitization in planarians. Behav. Pharmacol. 2010, 21, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Guecheva, T.N.; Erdtmann, B.; Benfato, M.S.; Henriques, J.A.P. Stress protein response and catalase activity in freshwater planarian Dugesia (Girardia) schubarti exposed to copper. Ecotoxicol. Environ. Saf. 2003, 56, 351–357. [Google Scholar] [CrossRef]
- Rowlands, A.L.; Pagán, O.R. Parthenolide prevents the expression of cocaine-induced withdrawal behavior in planarians. Eur. J. Pharmacol. 2008, 583, 170–172. [Google Scholar] [CrossRef] [PubMed]
- Pagán, O.R.; Rowlands, A.L.; Azam, M.; Urban, K.R.; Bidja, A.H.; Roy, D.M.; Feeney, R.B.; Afshari, L.K. Reversal of cocaine-induced planarian behavior by parthenolide and related sesquiterpene lactones. Pharmacol. Biochem. Behav. 2008, 89, 160–170. [Google Scholar]
- Geethaa, S.; Thavamany, P.J.; Chiew, S.P.; Thong, O.M. Interference from ordinarily used solvents in the outcomes of Artemia salina lethality test. J. Adv. Pharm. Technol. Res. 2013, 4, 179–182. [Google Scholar] [PubMed]
- Stevens, A.S.; Pirotte, N.; Plusquin, M.; Willems, M.; Neyens, T.; Artois, T.; Smeets, S. Toxicity profiles and solvent–toxicant interference in the planarian Schmidtea mediterranea after dimethylsulfoxide (DMSO) exposure. J. Appl. Toxicol. 2015, 35, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.; Wang, Y.H.; Wu, Y.H. Developmental exposures to ethanol or dimethylsulfoxide at low concentrations alter locomotor activity in larval zebrafish: Implications for behavioral toxicity bioassays. Aquat. Toxicol. 2011, 102, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, I.R.; Martins, R.M.; Sá, E.; Melo, M.L.; Soares, A.M. Acute and chronic toxicity of dimethylsulfoxide to Daphnia magna. Bull. Environ. Contam. Toxicol. 2003, 70, 1264–1268. [Google Scholar] [CrossRef] [PubMed]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- Zhang, X.; Zhang, B.; Yi, H.; Zhao, B. Mortality and antioxidant responses in the planarian (Dugesia japonica) after exposure to copper. Toxicol. Ind. Health 2014, 30, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Isomae, K.; Morimoto, S.; Hasegawa, H.; Morita, K.; Kamei, J. Effects of T-82, a novel acetylcholinesterase inhibitor, on impaired learning and memory in passive avoidance task in rats. Eur. J. Pharmacol. 2003, 465, 97–103. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds isolated comercial donepezil, tacrin, galantamin and rivastigmin are available from the authors.
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bezerra da Silva, C.; Pott, A.; Elifio-Esposito, S.; Dalarmi, L.; Fialho do Nascimento, K.; Moura Burci, L.; De Oliveira, M.; De Fátima Gaspari Dias, J.; Maria Warumby Zanin, S.; Gomes Miguel, O.; et al. Effect of Donepezil, Tacrine, Galantamine and Rivastigmine on Acetylcholinesterase Inhibition in Dugesia tigrina. Molecules 2016, 21, 53. https://doi.org/10.3390/molecules21010053
Bezerra da Silva C, Pott A, Elifio-Esposito S, Dalarmi L, Fialho do Nascimento K, Moura Burci L, De Oliveira M, De Fátima Gaspari Dias J, Maria Warumby Zanin S, Gomes Miguel O, et al. Effect of Donepezil, Tacrine, Galantamine and Rivastigmine on Acetylcholinesterase Inhibition in Dugesia tigrina. Molecules. 2016; 21(1):53. https://doi.org/10.3390/molecules21010053
Chicago/Turabian StyleBezerra da Silva, Cristiane, Arnildo Pott, Selene Elifio-Esposito, Luciane Dalarmi, Kátia Fialho do Nascimento, Ligia Moura Burci, Maislian De Oliveira, Josiane De Fátima Gaspari Dias, Sandra Maria Warumby Zanin, Obdulio Gomes Miguel, and et al. 2016. "Effect of Donepezil, Tacrine, Galantamine and Rivastigmine on Acetylcholinesterase Inhibition in Dugesia tigrina" Molecules 21, no. 1: 53. https://doi.org/10.3390/molecules21010053