
Identification of the Main Intermediate Precursor of l-Ergothioneine Biosynthesis in Human Biological Specimens

 ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
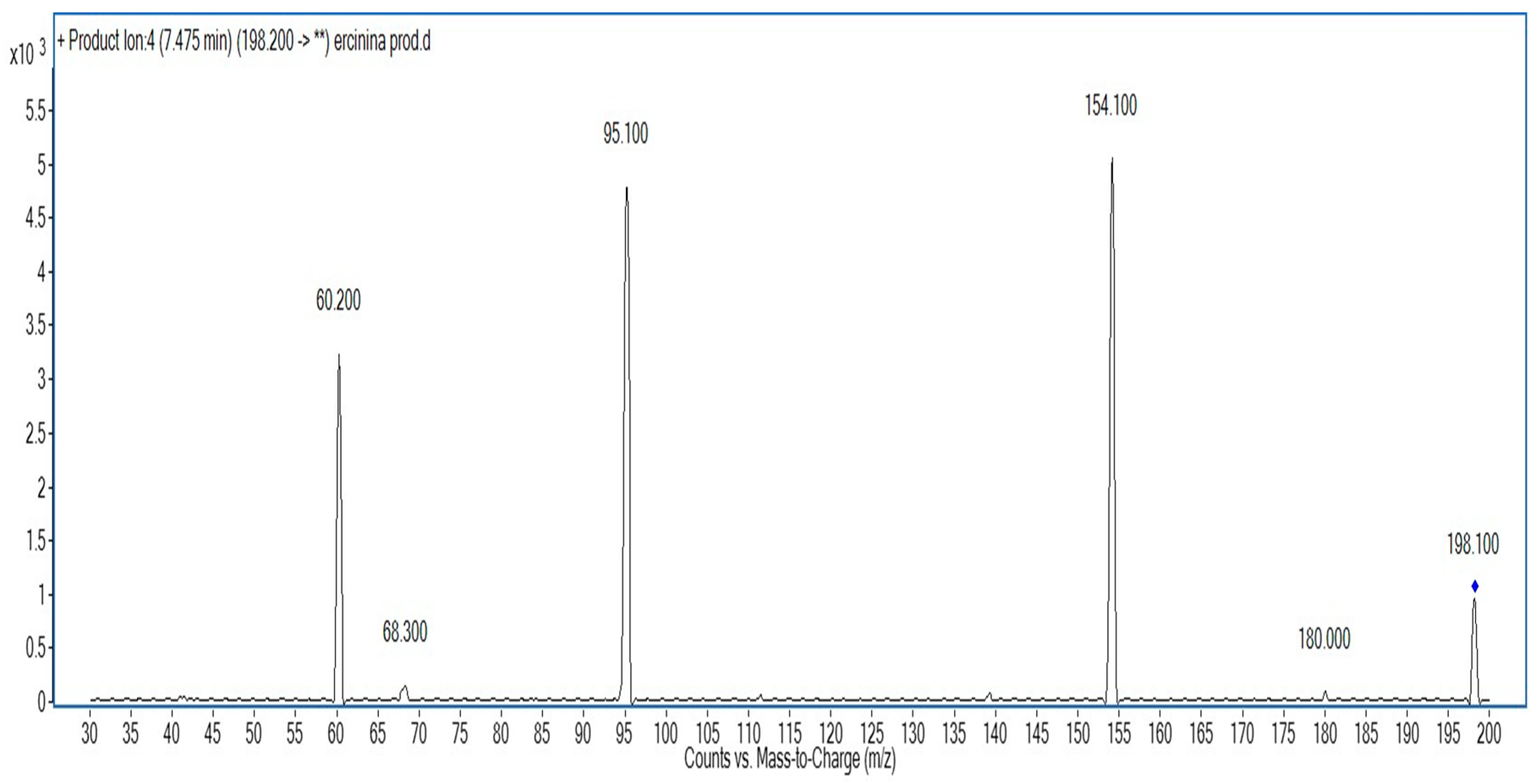
2. Results and Discussion
3. Material and Methods
3.1. Chemicals
3.2. Collection and Processing of Biological Samples
3.3. Hercynine Synthesis
3.4. Semiquantitative Analysis
3.5. CE Equipment and Electrophoretic Conditions
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Paul, B.D.; Snyder, S.H. The unusual amino acid l-ergothioneine is a physiologic cytoprotectant. Cell Death Differ. 2010, 17, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Ey, J.; Schömig, E.; Taubert, D. Dietary sources and antioxidant effects of ergothioneine. J. Agric. Food Chem. 2007, 55, 6466–6474. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, C.; Bauer, T.; Surek, B.; Schömig, E.; Gründemann, D. Cyanobacteria produce high levels of ergothioneine. Food Chem. 2011, 129, 1766–1769. [Google Scholar] [CrossRef]
- Gründemann, D. The ergothioneine transporter controls and indicates ergothioneine activity—A review. Prev. Med. 2012, 54, S71–S74. [Google Scholar] [CrossRef] [PubMed]
- Tokuhiro, S.; Yamada, R.; Chang, C.; Suzuki, A.; Kochi, Y.; Sawada, T.; Suzuki, M.; Nagasaki, M.; Ohtsuki, M.; Ono, M.; et al. An intronic SNP in a RUNX1 binding site of SLC22A4, encoding an organic cation transporter, is associated with rheumatoid arthritis. Nat. Genet. 2003, 35, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Peltekova, V.D.; Wintle, R.F.; Rubin, L.A.; Amos, C.I.; Huang, Q.; Gu, X.; Newman, B.; Oene, M.V.; Cescon, D.; Greenberg, G.; et al. Functional variants of OCTN cation transporter genes are associated with Crohn disease. Nat. Genet. 2004, 36, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Turner, E.; Brewster, J.A.; Simpson, N.A.; Walker, J.J.; Fisher, J. Imidazole-based erythrocyte markers of oxidative stress in preeclampsia-an NMR investigation. Reprod. Sci. 2009, 16, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Sotgia, S.; Zinellu, A.; Arru, D.; Nieddu, S.; Strina, A.; Ariu, D.; Pintus, G.; Carru, C.; Bogliolo, L.; Ledda, S. Amniotic fluid l-ergothioneine concentrations in pregnant sheep after natural mating and transfer of vitrified/thawed in vitro produced embryos. Res. Vet. Sci. 2015, 102, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Shires, T.K.; Brummel, M.C.; Pulido, J.S.; Stegink, L.D. Ergothioneine distribution in bovine and porcine ocular tissues. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1997, 117, 117–120. [Google Scholar] [CrossRef]
- Arduini, A.; Mancinelli, G.; Radatti, G.L.; Hochstein, P.; Cadenas, E. Possible mechanism of inhibition of nitrite-induced oxidation of oxyhemoglobin by l-ergothioneine and uric acid. Arch. Biochem. Biophys. 1992, 294, 398–402. [Google Scholar] [CrossRef]
- Misiti, F.; Castagnola, M.; Zuppi, C.; Giardina, B.; Messana, I. Role of ergothioneine on S-nitrosoglutathione catabolism. Biochem. J. 2001, 356, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Cheah, I.K.; Halliwell, B. Ergothioneine; antioxidant potential, physiological function and role in disease. Biochim. Biophys. Acta 2012, 1822, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Gründemann, D.; Harlfinger, S.; Golz, S.; Geerts, A.; Lazar, A.; Berkels, R.; Jung, N.; Rubbert, A.; Schömig, E. Discovery of the ergothioneine transporter. Proc. Natl. Acad. Sci. USA 2005, 102, 5256–5261. [Google Scholar] [CrossRef] [PubMed]
- Melville, D.B. Ergothioneine. Vitam. Horm. 1959, 17, 155–204. [Google Scholar]
- Servillo, L.; Castaldo, D.; Casale, R.; D’Onofrio, N.; Giovane, A.; Cautela, D.; Balestrieri, M.L. An uncommon redox behavior sheds light on the cellular antioxidant properties of ergothioneine. Free Radic. Biol. Med. 2015, 79, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Heath, H.; Toennies, G. The preparation and properties of ergothioneine disulfide. Biochem. J. 1958, 68, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, V.N.; Ishikawa, Y.; Melville, D.B. Conversion of histidine to hercynine by Neurospora crassa. J. Bacteriol. 1970, 101, 881–884. [Google Scholar] [PubMed]
- Servillo, L.; Giovane, A.; Casale, R.; D’Onofrio, N.; Ferrari, G.; Cautela, D.; Castaldo, D.; Balestrieri, M.L. Occurrence and Analysis of Betaines in Fruits in Betaine. In Betaine: Chemistry, Analysis, Function and Effects; Preedy, V.R., Ed.; Royal Society of Chemistry: Cambridge, UK, 2015; pp. 178–199. [Google Scholar]
- Grigat, S.; Harlfinger, S.; Pal, S.; Striebinger, R.; Golz, S.; Geerts, A.; Lazar, A.; Schömig, E.; Gründemann, D. Probing the substrate specificity of the ergothioneine transporter with methimazole, hercynine, and organic cations. Biochem. Pharmacol. 2007, 74, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Heath, H. The metabolism of 35S-labelled 2-thiolhistidine and ergothioneine in the rat. Biochem. J. 1953, 54, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Kumosani, T.A. l-Ergothioneine level in red blood cells of healthy human males in the Western province of Saudi Arabia. Exp. Mol. Med. 2001, 33, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Chaudière, J.; Ferrari-Iliou, R. Intracellular antioxidants: From chemical to biochemical mechanisms. Food Chem. Toxicol. 1999, 37, 949–962. [Google Scholar] [CrossRef]
- Arduini, A.; Eddy, L.; Hochstein, P. The reduction of ferryl myoglobin by ergothioneine: A novel function for ergothioneine. Arch. Biochem. Biophys. 1990, 281, 41–43. [Google Scholar] [CrossRef]
- Robertson, D.S. Biochemical and biomedical aspects of metabolic imidazoles. Clin. Biochem. 2011, 44, 1363–1369. [Google Scholar] [CrossRef] [PubMed]
- Markova, N.G.; Karaman-Jurukovska, N.; Dong, K.K.; Damaghi, N.; Smiles, K.A.; Yarosh, D.B. Skin cells and tissue are capable of using l-ergothioneine as an integral component of their antioxidant defense system. Free Radic. Biol. Med. 2009, 46, 1168–1176. [Google Scholar] [CrossRef] [PubMed]
- Konishi, Y.; Tamaki, N.; Tunemori, F.; Masumitu, N.; Okumura, H.; Hama, T. Studies on l-Ergothioneine II, Origin of l-Ergothioneine in animal organs. Vitam 1972, 46, 127–130. [Google Scholar]
- Barger, G.; Ewin, A.J. The constitution of ergothioneine: A betaine related to histidine. J. Chem. Soc. Trans. 1911, 99, 2336–2341. [Google Scholar] [CrossRef]
- Cheah, I.K.; Tang, R.M.; Yew, T.S.; Lim, K.H.; Halliwell, B. Administration of Pure Ergothioneine to Healthy Human Subjects: Uptake, Metabolism, and Effects on Biomarkers of Oxidative Damage and Inflammation. Antioxid. Redox Signal. 2016. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Biological samples are not available from the authors.



{kind=link}
{kind=link}
| Samples | ERT (µmol/L ± SD) | ERY (µmol/L ± SD) |
|---|---|---|
| Whole Blood (N = 3) | 66 ± 2.2 | 1.3 ± 0.1 |
| Plasma (N = 3) | 1.1 ± 0.3 | 0.2 ± 0.1 |
| Urine (N = 3) | 0.2 ± 0.2 | 0.5 ± 0.3 |
| Saliva (N = 3) | 0.6 ± 0.2 | 2.2 ± 0.5 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sotgia, S.; Mangoni, A.A.; Forteschi, M.; Murphy, R.B.; Elliot, D.; Sotgiu, E.; Pintus, G.; Carru, C.; Zinellu, A. Identification of the Main Intermediate Precursor of l-Ergothioneine Biosynthesis in Human Biological Specimens. Molecules 2016, 21, 1298. https://doi.org/10.3390/molecules21101298
Sotgia S, Mangoni AA, Forteschi M, Murphy RB, Elliot D, Sotgiu E, Pintus G, Carru C, Zinellu A. Identification of the Main Intermediate Precursor of l-Ergothioneine Biosynthesis in Human Biological Specimens. Molecules. 2016; 21(10):1298. https://doi.org/10.3390/molecules21101298
Chicago/Turabian StyleSotgia, Salvatore, Arduino A. Mangoni, Mauro Forteschi, Rhys B. Murphy, David Elliot, Elisabetta Sotgiu, Gianfranco Pintus, Ciriaco Carru, and Angelo Zinellu. 2016. "Identification of the Main Intermediate Precursor of l-Ergothioneine Biosynthesis in Human Biological Specimens" Molecules 21, no. 10: 1298. https://doi.org/10.3390/molecules21101298
APA StyleSotgia, S., Mangoni, A. A., Forteschi, M., Murphy, R. B., Elliot, D., Sotgiu, E., Pintus, G., Carru, C., & Zinellu, A. (2016). Identification of the Main Intermediate Precursor of l-Ergothioneine Biosynthesis in Human Biological Specimens. Molecules, 21(10), 1298. https://doi.org/10.3390/molecules21101298