
Simultaneous Saccharification and Fermentation of Sugar Beet Pulp with Mixed Bacterial Cultures for Lactic Acid and Propylene Glycol Production

 ,
, 

Abstract
:1. Introduction
2. Results and Discussion
2.1. Sugar Beet Pulp Hydrolyzate as a Carbon Source for Lactic Acid Fermentation
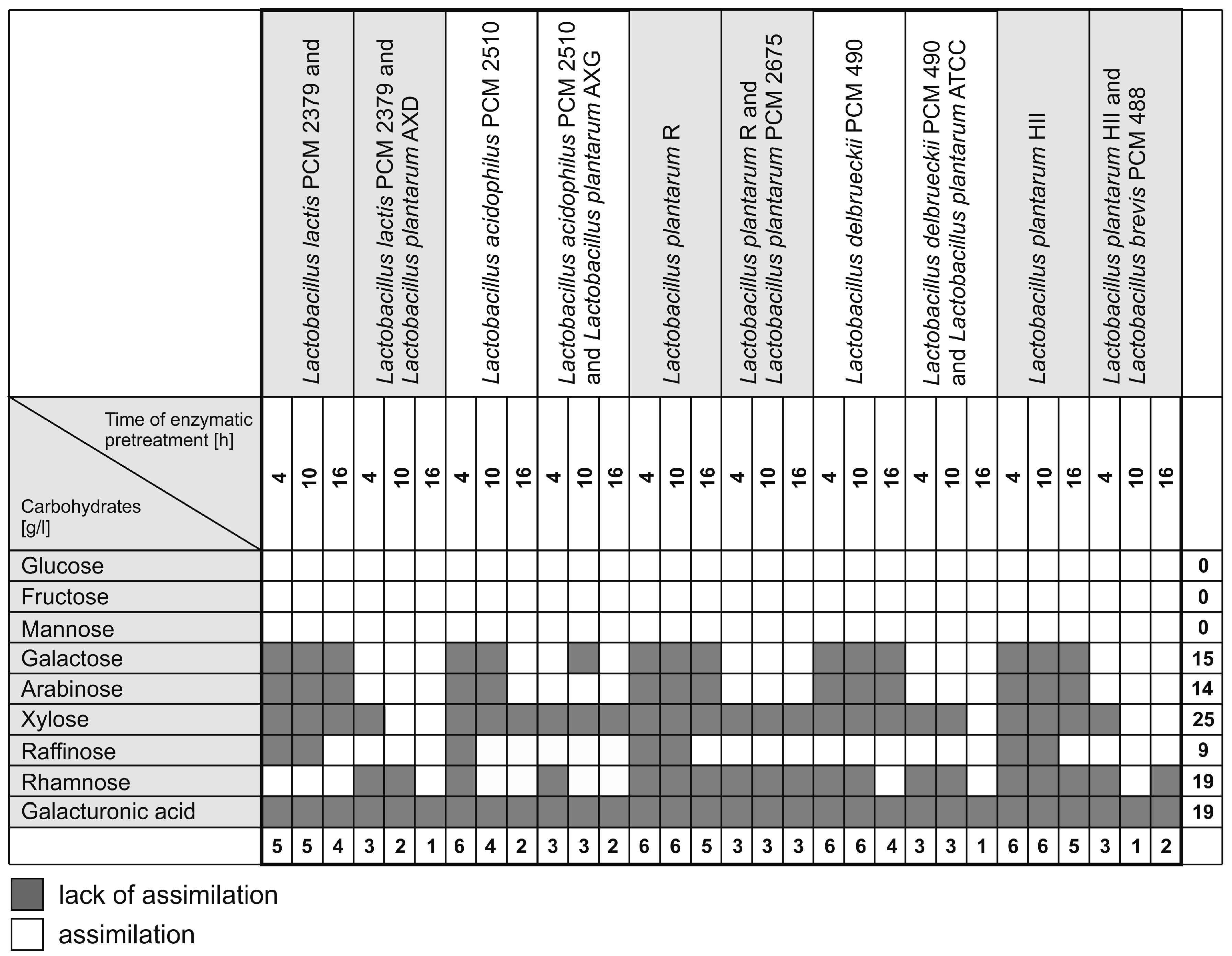
2.2. Carbon Utilization Profiles of Tested LAB Strains
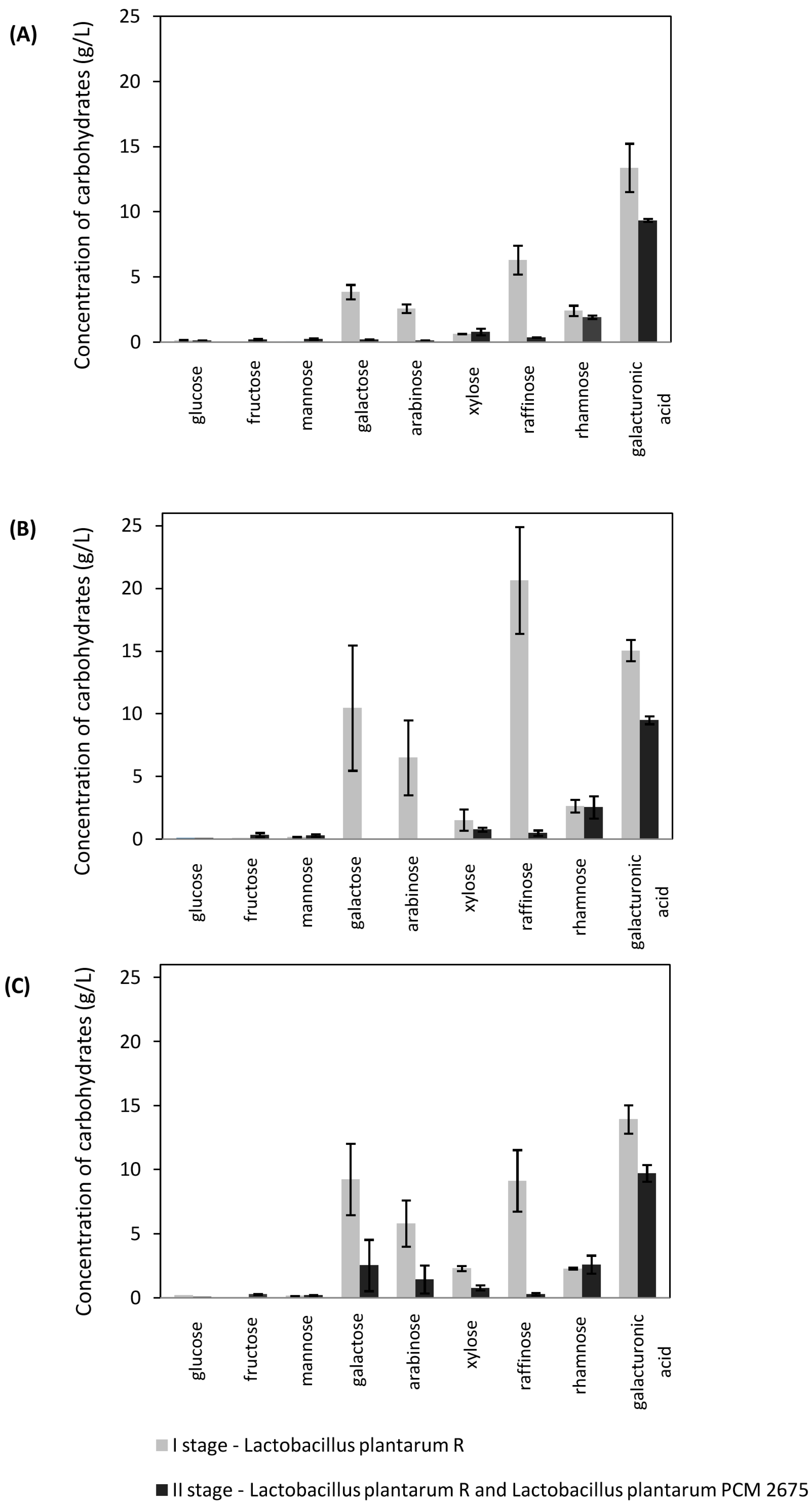
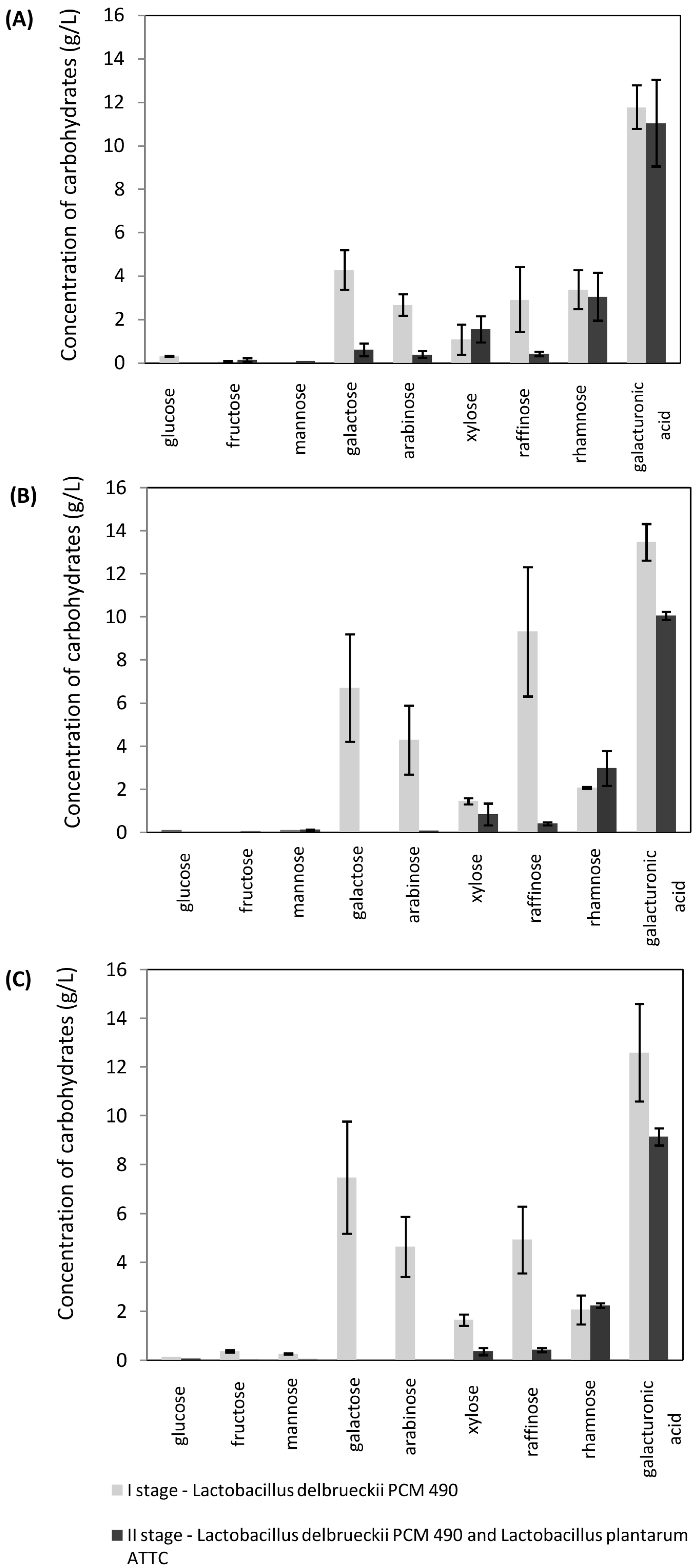
2.3. Lactic Acid Fermentation of Sugar Beet Hydrolyzates Using Mixed Cultures
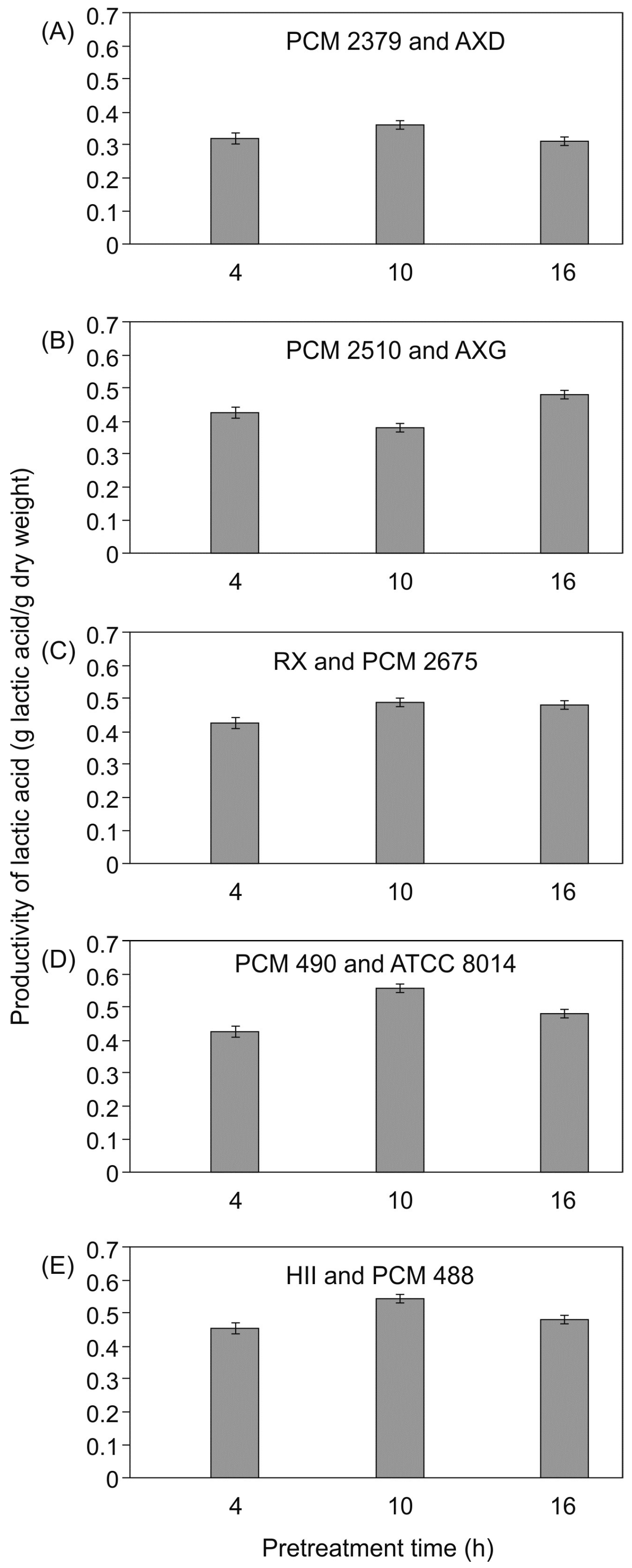
2.4. Lactic Acid Fermentation of Sugar Beet Hydrolyzates in SSF Mode
2.5. Biologically-Synthesized Lactic Acid as a Platform Chemical for Propylene Glycol Synthesis
3. Materials and Methods
3.1. Biological Material
3.1.1. Bacterial Strains
3.1.2. Plant Material
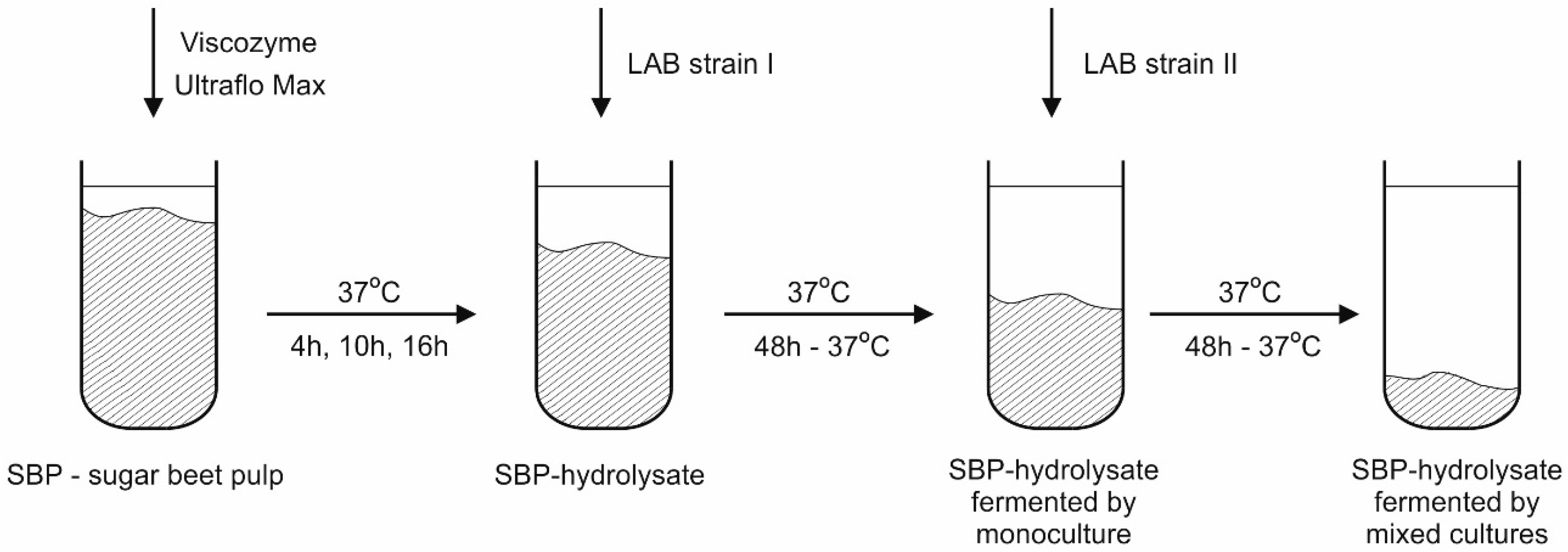
3.2. Hydrolysis and Fermentation
3.2.1. Inoculum Preparation
3.2.2. Sugar Beet Pulp Biomass Preparation and Enzymatic Pre-Treatment
3.2.3. Fermentation
3.2.4. Effectiveness of Hydrolysis
3.3. Propylene Glycol Synthesis
3.4. Culture Media Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- John, R.P.; Nampoothiri, K.M.; Pandey, A. Simultaneous saccharification and l-(+)-lactic acid fermentation of protease-treated wheat bran using mixed culture of lactobacilli. Biotechnol. Lett. 2006, 28, 1823–1826. [Google Scholar] [CrossRef] [PubMed]
- Hofvendahl, K.; Hahn-Hagerdal, B. Factor affecting the fermentative lactic acid production from renewable resources. Enzyme Microb. Technol. 2000, 26, 87–107. [Google Scholar] [CrossRef]
- Nguyen, C.M.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Kima, J.C. d- and l-lactic acid production from fresh sweet potato through simultaneous saccharification and fermentation. Biochem. Eng. J. 2013, 81, 40–46. [Google Scholar] [CrossRef]
- Wang, Y.; Tashiro, Y.; Sonomoto, K. Fermentative production of lactic acid from renewable materials: Recent achievements, prospects, and limits. J. Biosci. Bioeng. 2015, 119, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, X.; Luo, J.; Qi, B.; Wan, Y. An efficient process for lactic acid production from wheat straw by a newly isolated Bacillus coagulans strain IPE22. Bioresour. Technol. 2014, 158, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Qiao, Q.; Chu, D.; Gu, H.; Dao, T.H.; Zhang, J.; Bao, J. Simultaneous saccharification and high titer lactic acid fermentation of corn stover using a newly isolated lactic acid bacterium Pediococcus acidilactici DQ2. Bioresour. Technol. 2013, 135, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Ohgren, K.; Bura, R.; Lesnicki, G.; Saddler, J.; Zacchi, G. A comparison between simultaneous saccharification and fermentation and separate hydrolysis and fermentation using steam-pretreated corn stover. Process Biochem. 2007, 42, 834–839. [Google Scholar] [CrossRef]
- Stenberg, K.; Galbe, M.; Zacchi, G. The influence of lactic acid formation on the simultaneous saccharification and fermentation (SSF) of softwood to etanol. Enzyme Microb. Technol. 2000, 26, 71–79. [Google Scholar] [CrossRef]
- Sasaki, C.; Kushiki, Y.; Asada, C.; Nakamura, Y. Acetone–butanol–ethanol production by separate hydrolysis and fermentation (SHF) and simultaneous saccharification and fermentation (SSF) methods using acorns and wood chips of Quercus acutissima as a carbon source. Ind. Crops Prod. 2014, 62, 286–292. [Google Scholar] [CrossRef]
- Gao, C.; Ma, C.; Xu, P. Biotechnological routes based on lactic acid production from biomass. Biotechnol. Adv. 2011, 29, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Pal, P.; Sikder, J.; Roy, S.; Giorno, L. Process intensification in lactic acid production: A review of membrane based processes. Chem. Eng. Prog. 2009, 48, 1549–1559. [Google Scholar] [CrossRef]
- Berłowska, J.; Binczarski, M.; Dudkiewicz, M.; Kalinowska, H.; Witonska, I.A.; Stanishevsky, A.V. A low-cost method for obtaining high-value bio-based propylene glycol from sugar beet pulp. RSC Adv. 2015, 5, 2299–2304. [Google Scholar] [CrossRef]
- Leijdekkers, A.G.; Bink, J.P.; Geutjes, S.; Schols, H.A.; Gruppen, H. Enzymatic saccharification of sugar beet pulp for the production of galacturonic acid and arabinose; a study on the impact of the formation of recalcitrant oligosaccharides. Bioresour. Technol. 2013, 128, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Olmos, J.C.; Hansen, M.E.Z. Enzymatic depolymerization of sugar beet pulp: Production and characterization of pectin and pectic-oligosaccharides as a potential source for functional Carbohydrates. Chem. Eng. J. 2012, 192, 29–36. [Google Scholar] [CrossRef]
- Micard, V.; Renard, C.M.G.C.; Thibault, J.-F. Enzymatic saccharification of sugar beet pulp. Enzym. Microb. Technol. 1996, 19, 162–170. [Google Scholar] [CrossRef]
- Ward, D.P.; Cárdenas-Fernández, M.; Hewitson, P.; Ignatova, S.; Lye, G.J. Centrifugal partition chromatography in a biorefinery context: Separation of monosaccharides from hydrolysed sugar beet pulp. J. Chromatogr. A 2015, 1411, 84–91. [Google Scholar] [CrossRef] [PubMed]
- HamLey-Bennett, C.; Lye, G.J.; Leak, D. Selective fractionation of Sugar Beet Pulp for release of fermentation and chemical feedstocks; optimisation of thermo-chemical pre-treatment. Bioresour. Technol. 2016, 209, 259–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, F.; Li, Y.; Wan, C. Lactic acid production from corn stover using mixed cultures of Lactobacillus rhamnosus and Lactobacillus brevis. Bioresour. Technol. 2011, 102, 1831–1836. [Google Scholar] [CrossRef] [PubMed]
- Kandeler, O. Carbohydrate metabolism in lactic acid bacteria. Antonie van Leeuwenhoek 1983, 49, 209–224. [Google Scholar] [CrossRef]
- Gobbetti, M.; Lavermicocca, P.; Minervini, F.; De Angelis, M.; Corsetti, A. Arabinose fermentation by Lactobacillus plantarum in sourdough with added pentosans and a-L-arabinofuranosidase: a tool to increase the production of acetic acid. J. Appl. Microbiol. 2000, 88, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Lactic acid production from lignocellulose-derived sugars using lactic acid bacteria: Overview and limits. J. Biotechnol. 2011, 156, 286–301. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Recent advances in lactic acid production by microbial fermentation processes. Biotechnol. Adv. 2013, 31, 877–909. [Google Scholar] [CrossRef] [PubMed]
- Ishola, M.M.; Ylitervo, P.; Taherzadeh, M.J. Co-Utilization of glucose and xylose for enhanced lignocellulosic ethanol production with reverse membrane bioreactors. Membranes 2015, 5, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Nancib, A.; Nancib, N.; Boudrant, J. Production of lactic acid from date juice extract with free cells of single and mixed cultures of Lactobacillus casei and Lactococcus lactis. World J. Microbiol. Biotechnol. 2009, 25, 1423–1429. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Xiao, Y.; Tashiro, Y.; Wang, Y.; Zendo, T.; Sakai, K.; Sonomoto, K. Fed-batch fermentation for enhanced lactic acid production from glucose/xylose mixture without carbon catabolite repression. J. Biosci. Bioeng. 2015, 119, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, M.; Tokunaga, T.; Horiuchi, K.; Hoshino, K.; Sakai, K.; Tanaka, T. Production of L-lactic acid from mixture of xylose and glucose by co-cultivation of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2004, 66, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Deutcher, J. The mechanisms of carbon catabolite repression in bacteria. Curr. Opin. Microbiol. 2008, 11, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Ishola, M.M.; Branbdberg, T.; Taherzadeh, M.J. Simultaneous glucose and xylose utilization for improved ethanol production from lignocellulosic biomass through SSFF with encapsulated yeast. Biomass Bioenerg. 2015, 77, 192–199. [Google Scholar] [CrossRef]
- Siragusa, S.; De Angelis, M.; Calasso, M.; Campanella, D.; Minervini, F.; Di Cagno, R.; Gobbetti, M. Fermentation and proteome profiles of Lactobacillus plantarum strains during growth under food-like conditions. J. Proteomics 2014, 96, 366–380. [Google Scholar] [CrossRef] [PubMed]
- John, R.P.; Nampoothiri, K.M.; Pandey, A. Simultaneous saccharification and fermentation of cassava bagasse for l-(+)-lactic acid production using Lactobacilli. Appl. Biochem. Biotechnol. 2006, 134, 263–272. [Google Scholar] [CrossRef]
- Shen, X.; Xia, L. Lactic acid production from cellulosic waste by immobilized cells of Lactobacillus delbrueckii. World J. Microbiol. Biotechnol. 2006, 22, 1109–1114. [Google Scholar] [CrossRef]
- Binczarski, M.; Berlowska, J.; Stanishevsky, A.; Witonska, I. Biologically synthesized crude calcium lactate as a substrate for propylene glycol production. RSC Adv. 2016, 6, 92420–92427. [Google Scholar] [CrossRef]
- Sample Availability: Samples of crude calcium lactate and propylene glycol are available from the authors.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time of hydrolysis | Carbohydrate Concentration (g/L) | ||
|---|---|---|---|
| 4 h | 10 h | 16 h | |
| Glucose | 18.61 ± 0.70 | 21.79 ± 0.54 | 29.74 ± 1.19 |
| Fructose | 4.52 ± 0.40 | 8.90 ± 0.29 | 12.46 ± 0.60 |
| Mannose | 3.04 ± 0.14 | 5.97 ± 0.17 | 7.04 ± 0.45 |
| Arabinose | 1.54 ± 0.50 | 2.60 ± 0.87 | 3.47 ± 0.82 |
| Galactose | 2.27 ± 0.90 | 3.90 ± 0.39 | 5.18 ± 0.31 |
| Raffinose | 2.48 ± 0.59 | 8.61 ± 0.71 | 16.24 ± 0.60 |
| Rhamnose | 0.88 ± 0.59 | 1.75 ± 0.08 | 2.26 ± 0.30 |
| Xylose | 0.39 ± 0.053 | 0.48 ± 0.038 | 0.47 ± 0.049 |
| Galacturonic acid | 3.66 ± 0.24 | 5.51 ± 0.44 | 7.81 ± 0.19 |
| Strain | Compound Utilization (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Glucocse | Fructose | Mannose | Arabinose | Galactose | Raffinose | Rhamnose | Xylose | Galacturonic acid | |
| Lactobacillus plantarum Hydr II | 100 | 99 | 94 | 0 | 0 | 35 | 7 | 0 | 37 |
| Lactobacillus acidophilus PCM 2510 | 100 | 100 | 95 | 0 | 0 | 0 | 0 | 0 | 27 |
| Lactococcus lactis PCM 2379 | 100 | 100 | 99 | 0 | 0 | 0 | 0 | 0 | 37 |
| Lactobacillus delbrueckii PCM 490 | 100 | 100 | 90 | 0 | 0 | 29 | 33 | 0 | 57 |
| Lactobacillus plantarum R | 100 | 99 | 91 | 0 | 0 | 42 | 7 | 0 | 37 |
| Lactobacillus plantarum AXD | 73 | 100 | 77 | 7 | 0 | 13 | 0 | 0 | 45 |
| Lactobacillus plantarum AXG | 68 | 100 | 78 | 26 | 0 | 0 | 0 | 0 | 35 |
| Lactobacillus brevis PCM 488 | 67 | 55 | 82 | 10 | 0 | 29 | 4 | 0 | 35 |
| Lactobacillus plantarum ATCC 8014 | 73 | 100 | 84 | 9 | 0 | 16 | 0 | 0 | 42 |
| Lactobacillus plantarum PCM 2675 | 76 | 100 | 81 | 5 | 7 | 11 | 0 | 0 | 59 |
| LAB Strains | Group I of LAB Strains | Group II of LAB Strains |
|---|---|---|
| I pair | Lactococcuss lactis PCM 2379 | Lactobacillus plantarum AXD |
| II pair | Lactobacillus acidophilus PCM 2510 | Lactobacillus plantarum AXG |
| III pair | Lactobacillus plantarum HII | Lactobacillus brevis PCM 488 |
| IV pair | Lactobacillus delbrueckii PCM 490 | Lactobacillus plantarum ATCC 8014 |
| V pair | Lactobacillus plantarum R | Lactobacillus plantarum PCM 2675 |
| Strains | Step | Hydrolyzate (g) | Lactic Acid (g/L) |
|---|---|---|---|
| Lactococcus lactis PCM 2379 | I | 44.79 ± 0.81 | 47.88 ± 1.50 |
| Lactococcus lactis PCM 2379 and Lactobacillus plantarum AXD | II | 46.85 ± 0.04 | 49.66 ± 0.70 |
| Lactobacillus acidophilus PCM 2510 | I | 53.74 ± 1.08 | 42.60 ± 2.37 |
| Lactobacillus acidophilus PCM 2510 and Lactobacillus plantarum AXG | II | 55.66 ± 0.10 | 52.61 ± 1.65 |
| Lactobacillus delbrueckii PCM 490 | I | 49.90 ± 0.11 | 57.78 ± 0.26 |
| Lactobacillus delbrueckii PCM 490 and Lactobacillus plantarum ATCC | II | 57.06 ± 0.97 | 58.39 ± 1.40 |
| Lactobacillus plantarum R | I | 46.92 ± 0.40 | 51.34 ± 0.26 |
| Lactobacillus plantarum R and Lactobacillus plantarum PCM 2675 | II | 57.37 ± 0.62 | 53.75 ± 0.14 |
| Lactobacillus plantarum HII | I | 48.31 ± 0.10 | 55.79 ± 0.54 |
| Lactobacillus plantarum HII and Lactobacillus brevis PCM 488 | II | 58.36 ± 0.48 | 59.57 ± 0.83 |
| Substratum | Initial Concentration of Lactate Ions (g/L) | Conversion of Lactate Ions (%) | Selectivity to Propylene Glycol (%) |
|---|---|---|---|
| Water solution of lactic acid | 45.04 | 96.41 | 63.12 |
| 60.32 | 52.71 | 96.82 | |
| Biologically synthesized lactic acid solution * | 41.80 | 53.63 | 67.42 |
| 43.96 | 88.76 | 68.44 | |
| 44.32 | 51.11 | 83.43 | |
| 45.85 | 49.52 | 94.12 | |
| 47.11 | 38.24 | 96.89 | |
| 50.08 | 43.83 | 94.89 | |
| 57.64 | 40.64 | 97.37 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berlowska, J.; Cieciura, W.; Borowski, S.; Dudkiewicz, M.; Binczarski, M.; Witonska, I.; Otlewska, A.; Kregiel, D. Simultaneous Saccharification and Fermentation of Sugar Beet Pulp with Mixed Bacterial Cultures for Lactic Acid and Propylene Glycol Production. Molecules 2016, 21, 1380. https://doi.org/10.3390/molecules21101380
Berlowska J, Cieciura W, Borowski S, Dudkiewicz M, Binczarski M, Witonska I, Otlewska A, Kregiel D. Simultaneous Saccharification and Fermentation of Sugar Beet Pulp with Mixed Bacterial Cultures for Lactic Acid and Propylene Glycol Production. Molecules. 2016; 21(10):1380. https://doi.org/10.3390/molecules21101380
Chicago/Turabian StyleBerlowska, Joanna, Weronika Cieciura, Sebastian Borowski, Marta Dudkiewicz, Michal Binczarski, Izabela Witonska, Anna Otlewska, and Dorota Kregiel. 2016. "Simultaneous Saccharification and Fermentation of Sugar Beet Pulp with Mixed Bacterial Cultures for Lactic Acid and Propylene Glycol Production" Molecules 21, no. 10: 1380. https://doi.org/10.3390/molecules21101380