
Improvement of Transmembrane Transport Mechanism Study of Imperatorin on P-Glycoprotein-Mediated Drug Transport


Abstract
:1. Introduction
2. Results
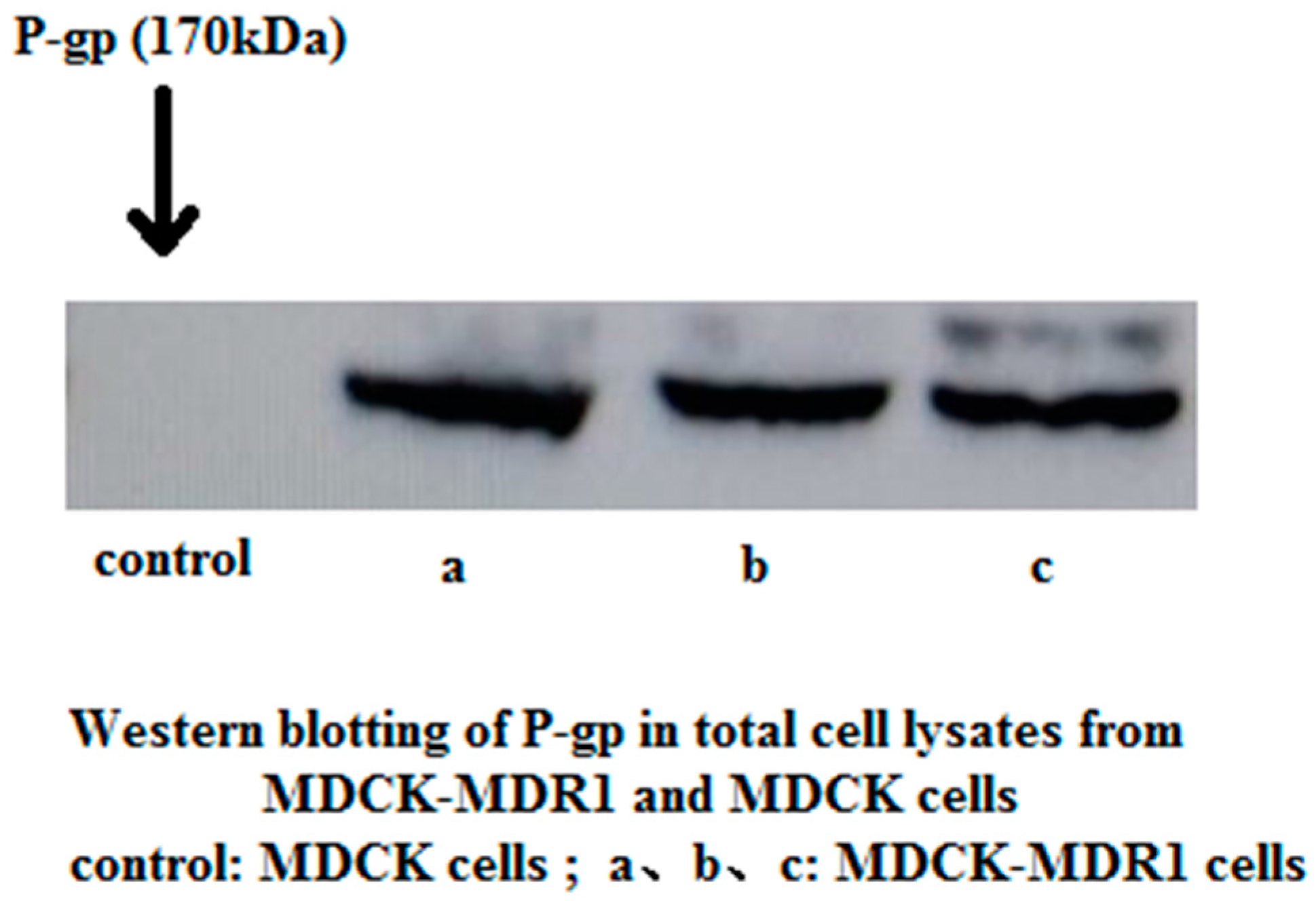
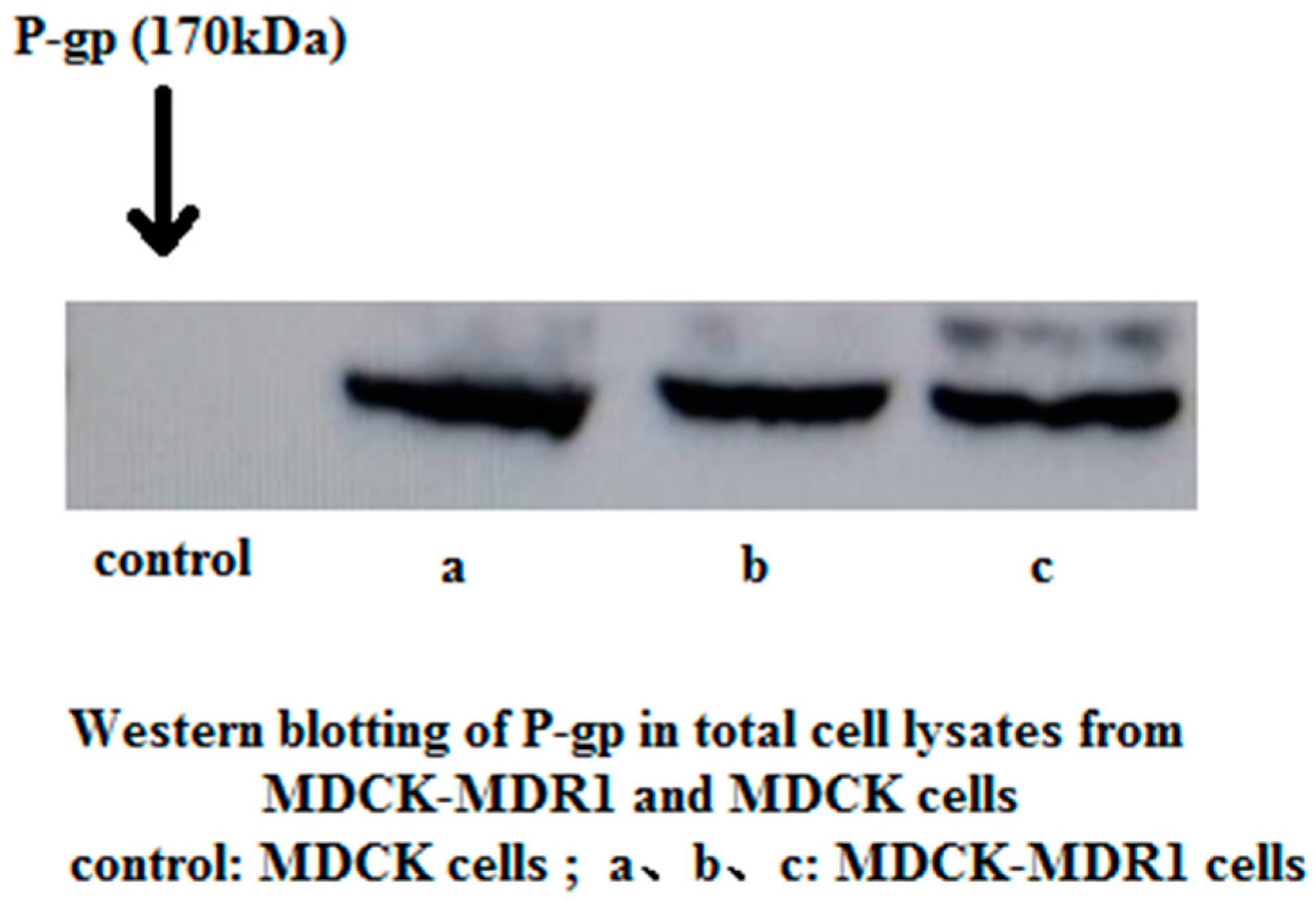
2.1. Western Blotting to Detect P-gp Expression in MDCK and MDCK-MDR1 Cells
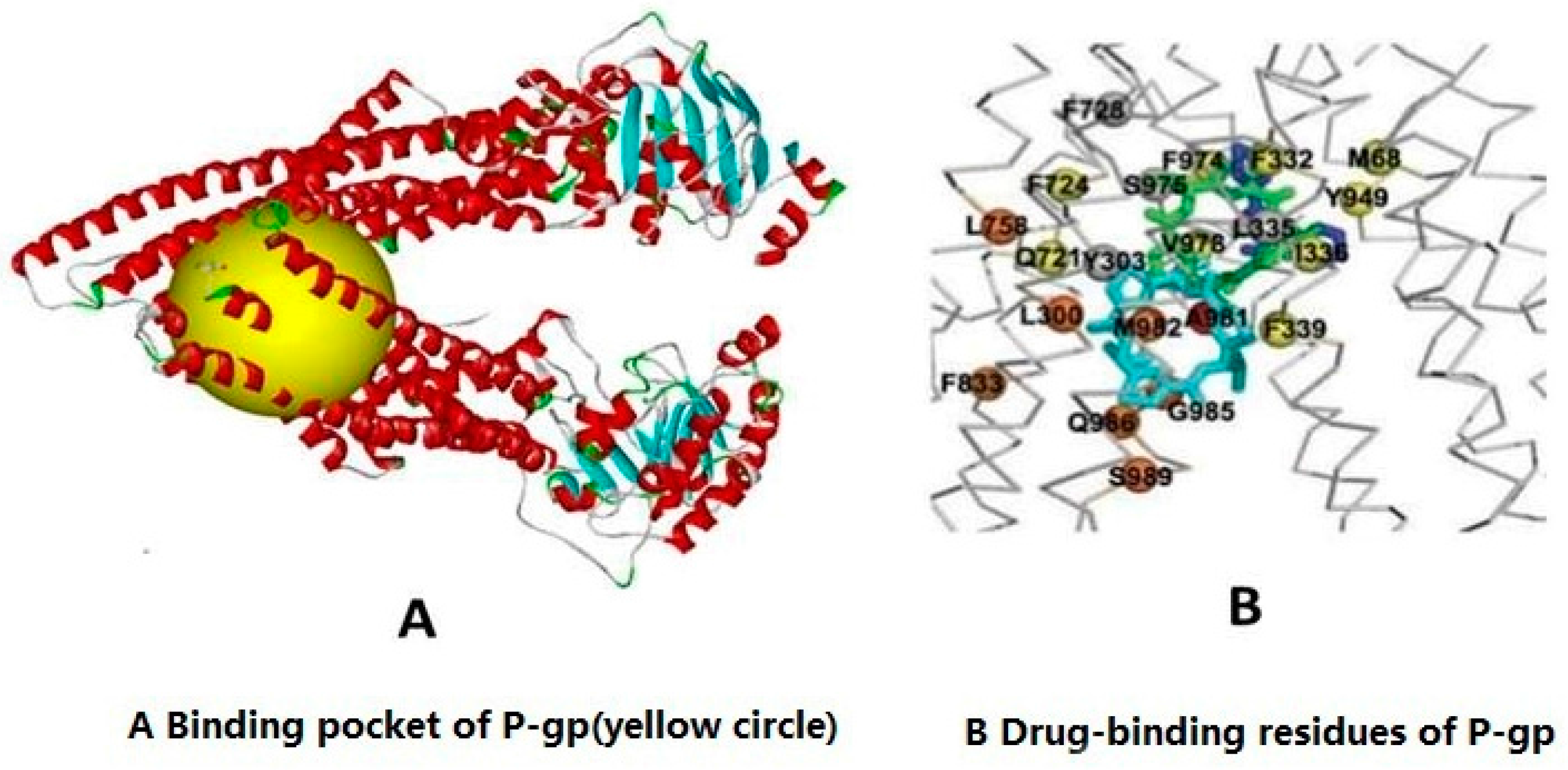
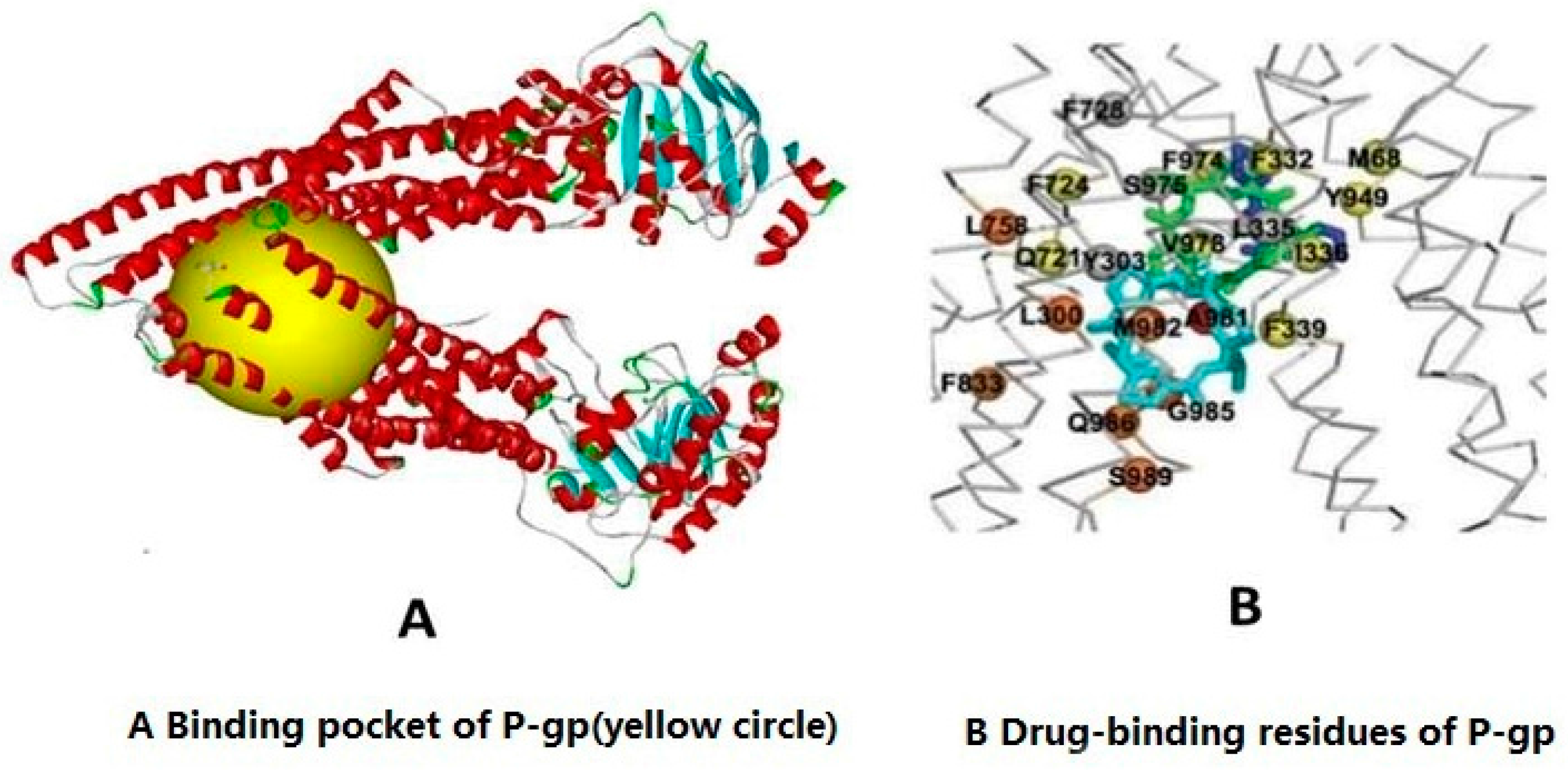
2.2. Molecular Docking Experiment to Detect the Association of Imperatorin and P-gp
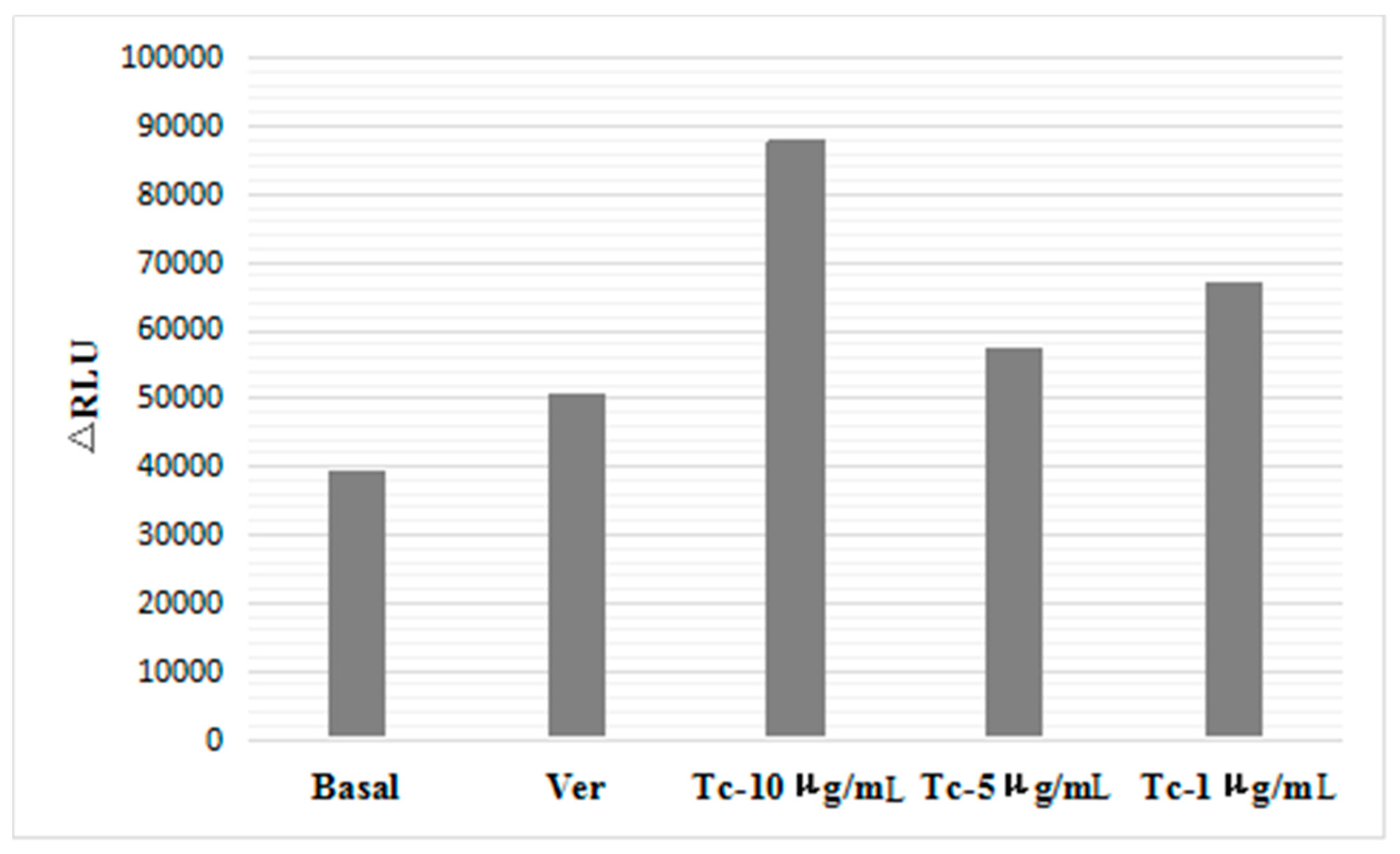
2.3. Effect of Imperatorin on P-gp Atpase Activity
2.4. Effect of Imperatorin on Membrane Fluidity
2.5. Inhibition of Rh-123 Efflux by Imperatorin
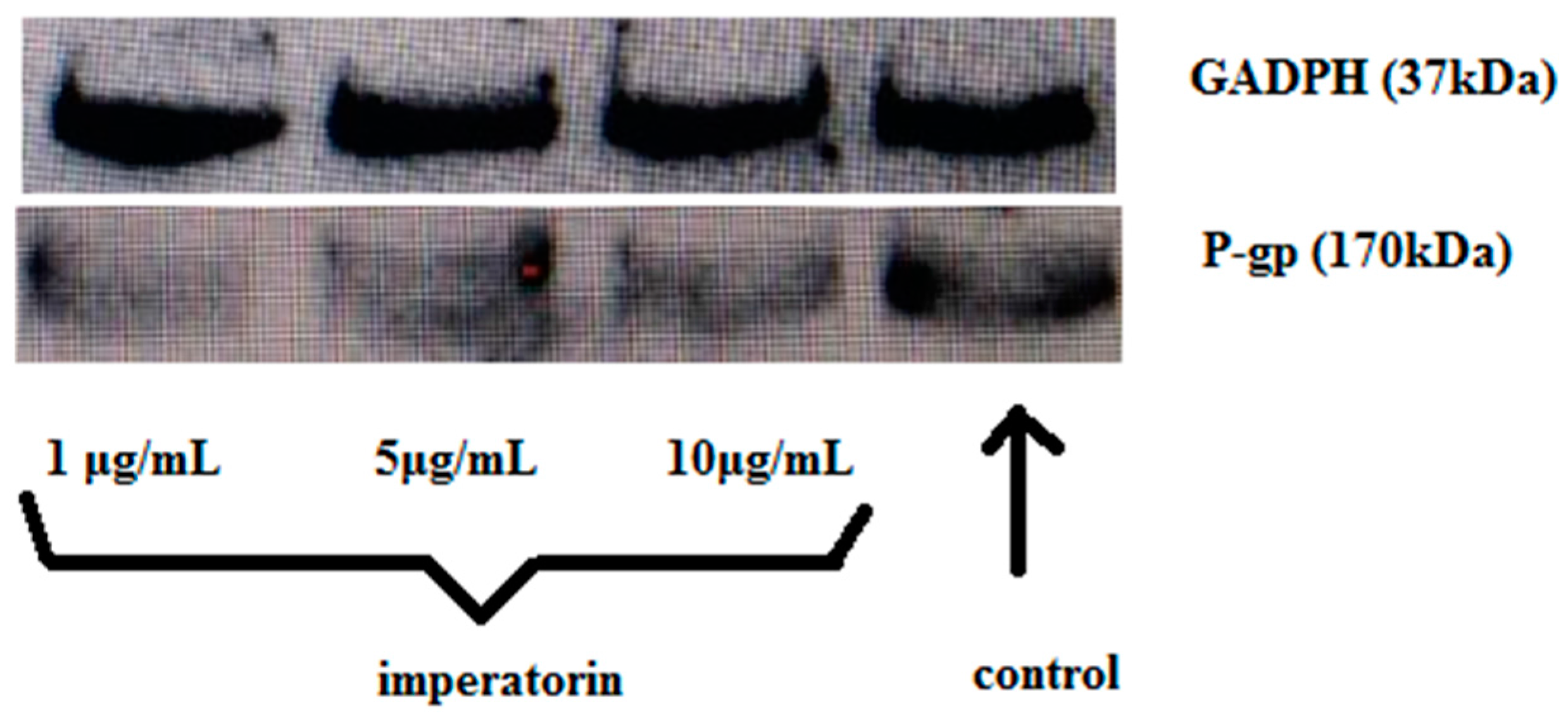
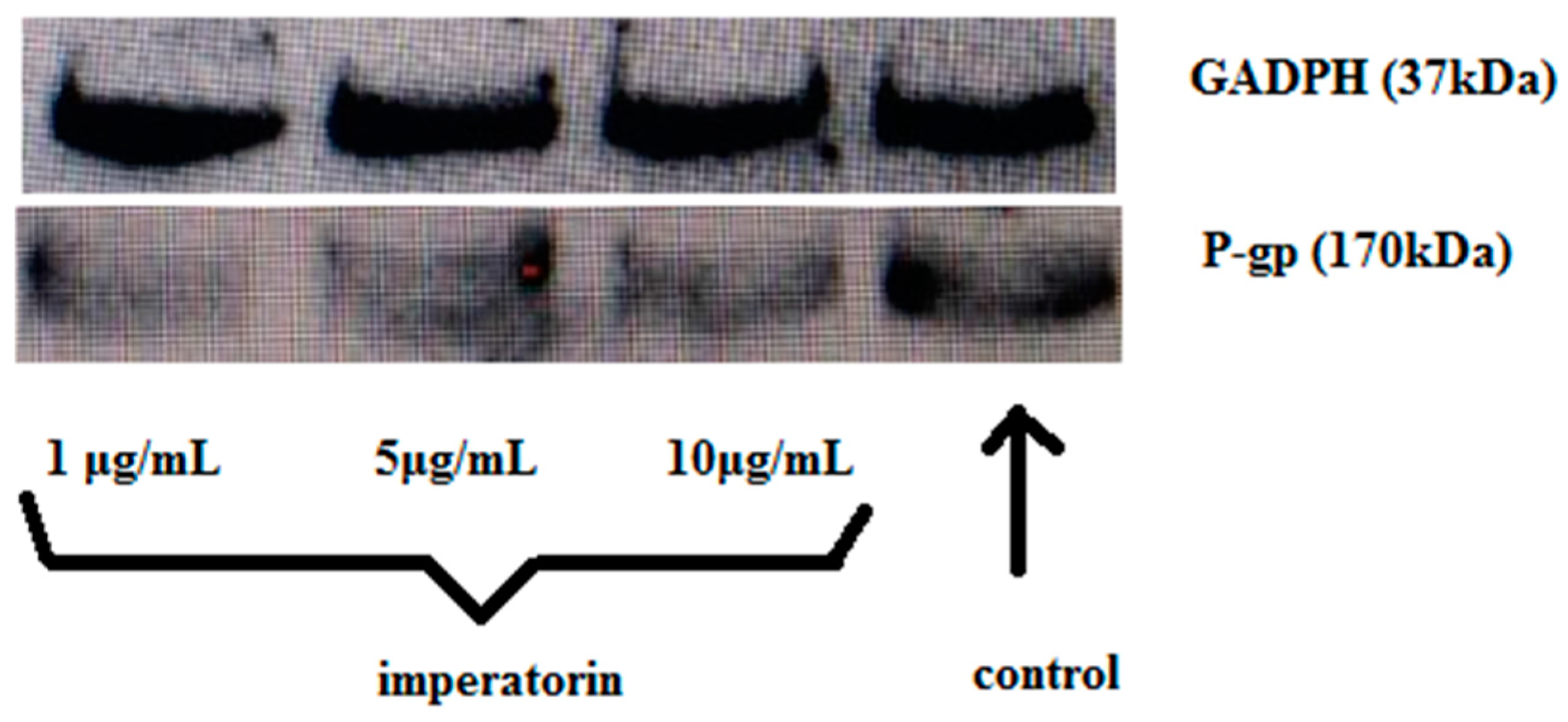
2.6. Effect of Imperatorin on P-gp Protein Expression
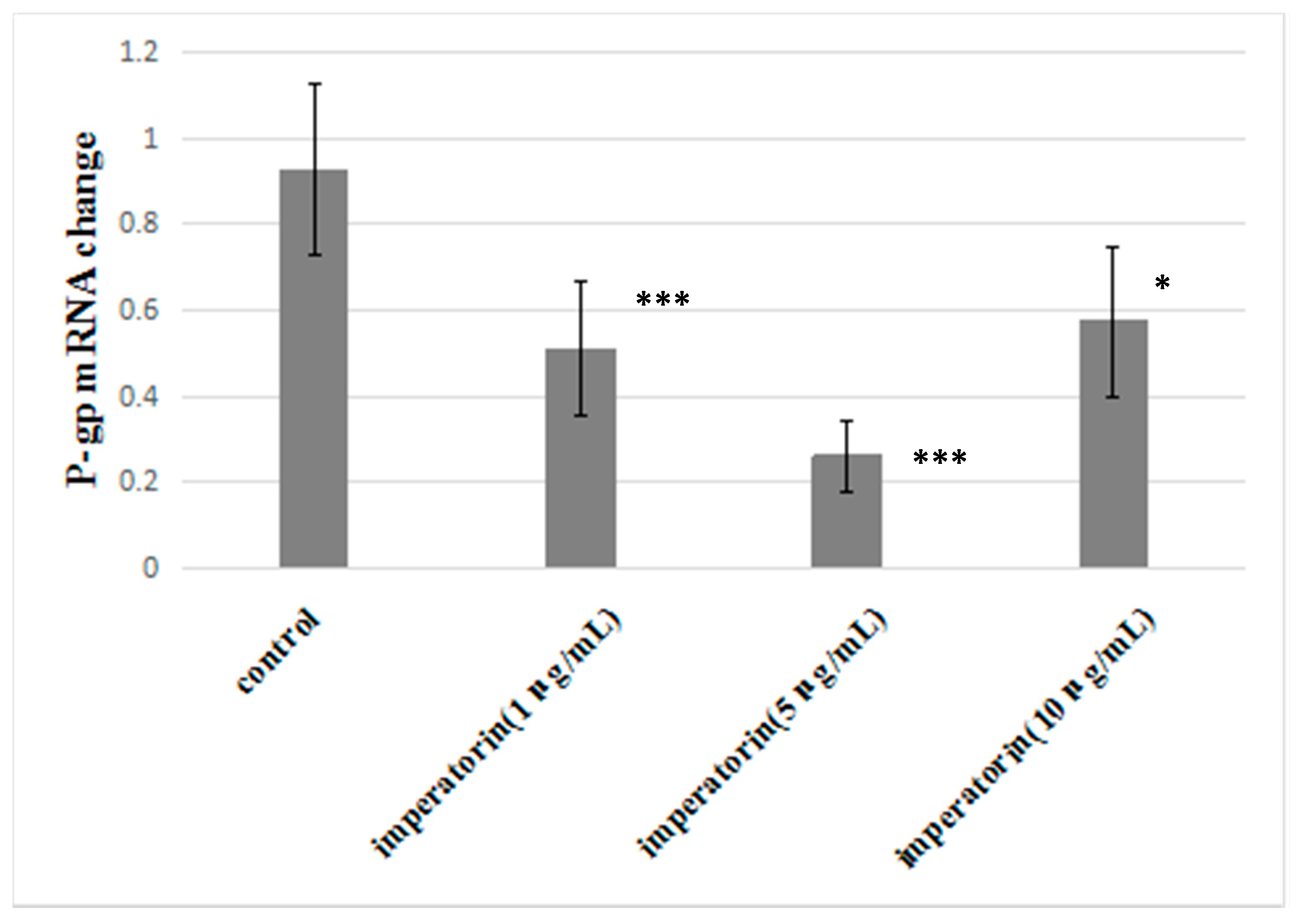
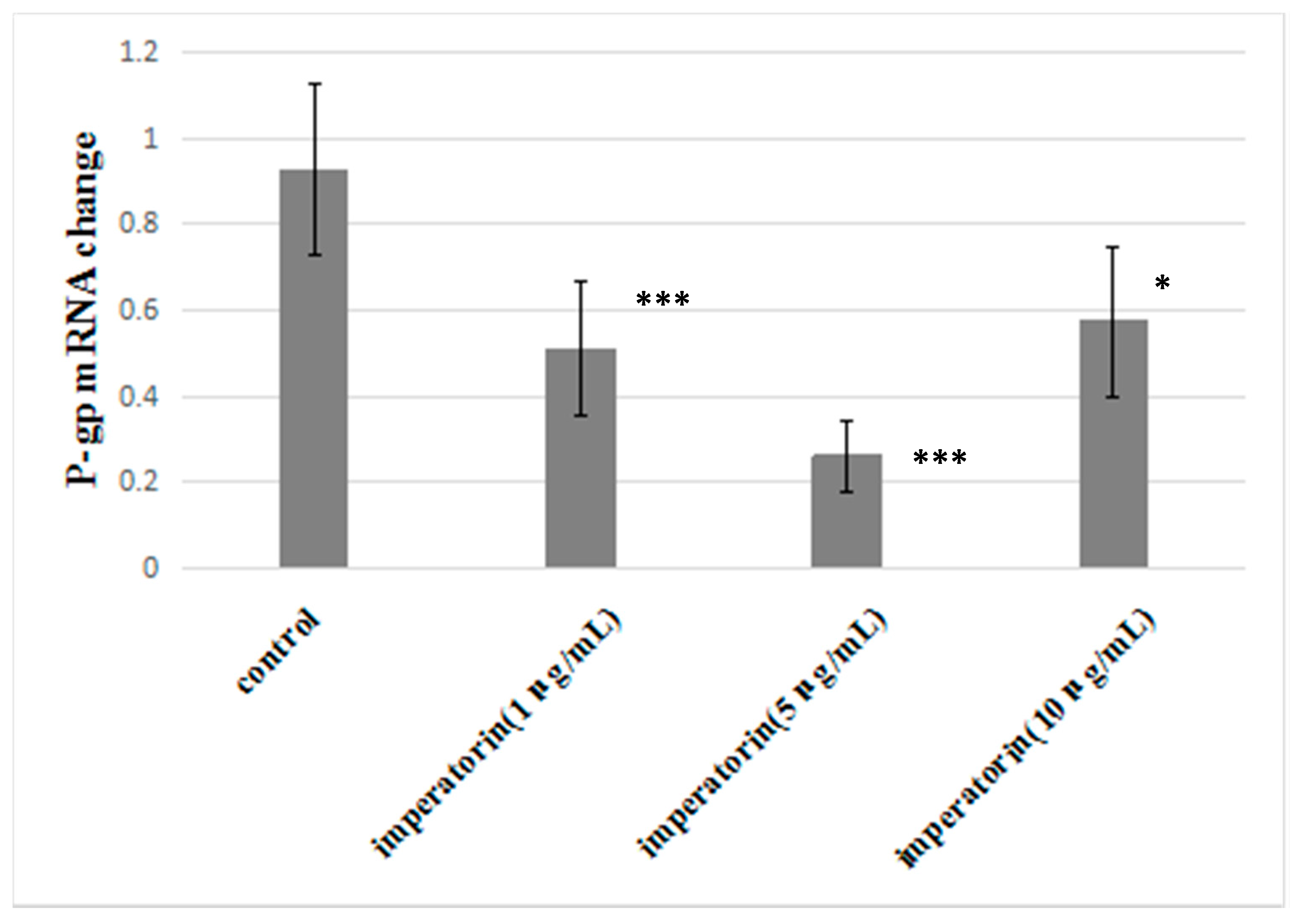
2.7. Effect of Imperatorin on P-gp mRNA Levels
3. Discussion
4. Materials and Methods
4.1. Chemicals and Drugs
4.2. Establishment and Validation of the MDCK-MDR1 Cell Model
4.2.1 MDCK Cell Culture
4.2.2 MDCK-MDR1 Cell Culture
4.2.3 Western Blotting to Detect P-gp Expression in MDCK and MDCK-MDR1 Cells
4.3. Molecular Docking
4.4. P-gp ATPase Activity Assay
4.5. Membrane Fluidity
4.6. Rhodamine 123 Accumulation
4.7. Effect of Imperatorin on P-gp Expression (Western Blotting)
4.8. qRT-PCR
4.9. Statistics
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DPH | 1,6-diphenyl-1,3,5-hexatriene |
| P-gp | P-glycoprotein |
| TCM | traditional Chinese medicine |
| FBS | fetal bovine serum |
| DMEM | Dulbecco’s modified Eagle’s medium |
| NEAA | non-essential amino acids |
| HBSS | Hank’s Balanced Salts Solution |
| Ver | Verapamil |
References
- Chan, L.M.; Lowes, S.; Hirst, B.H. The ABCs of drug transport in intestine and liver: Efflux proteins limiting drug absorption and bioavailability. Eur. J. Pharm. Sci. 2004, 21, 25–51. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.H.; Kang, G.; Min, Y.D. Reversal of P-glycoprotein-mediated multidrug resistance by protopanaxatriol ginsenosides from Korean red ginseng. Planta Med. 2003, 69, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhou, Y.; Zhang, J.; Liu, Y.; Guan, T.; Wang, Y.; Xing, L.; Rao, T.; Zhou, L.; Hao, K.; et al. In vitro to in vivo evidence of the inhibitor characteristics of Schisandra lignans toward P-glycoprotein. Phytomedicine 2013, 20, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Pleban, K.; Kopp, S.; Csaszar, E.; Peer, M.; Hrebicek, T.; Rizzi, A.; Ecker, G.F.; Chiba, P. P-glycoprotein substrate binding domains are located at the transmembrane domain/transmembrane domain interfaces: A combined photoaffinity labeling-protein homology modeling approach. Mol. Pharmacol. 2005, 67, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, M.F.; Callaghan, R.; Modok, S.; Higgins, C.F.; Ford, R.C. Three-dimensional structure of P-glycoprotein: The transmembrane regions adopt an asymmetric configuration in the nucleotide-bound state. J. Biol. Chem. 2005, 280, 2857–2862. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, M.M.; Pastan, I. Biochemistry of multidrug resistance mediated by the multidrug Transporter. Annu. Rev. Biochem. 1993, 62, 385–427. [Google Scholar] [CrossRef] [PubMed]
- Marzolini, C.; Paus, E.; Buclin, T.; Kim, R.B. Polymorphisms in Human MDR1 (P-glycoprotein): Recent Advances and Clinical Relevance. Clin. Pharmacol. Ther. 2004, 75, 13–33. [Google Scholar] [CrossRef] [PubMed]
- Teft, W.A.; Mansell, S.E.; Kim, R.B. Endoxifen, the active metabolite of tamoxifen, is a substrate of the efflux transporter P-glycoprotein (multidrug resistance 1). Drug Metab. Dispos. 2011, 39, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y. Research progress on Chinese material medica for reversing tumor multidrug resistance. Chin. Tradit. Herb. Drugs 2015, 46, 1096–1102. [Google Scholar]
- Gao, X.R.; Jiang, X.H.; Du, Q.Q.; Song, L. Effect of P-glyprotein inhibitors on the transport of dauricine through membrane. West Chin. J. Pharm. Sci. 2013, 28, 135–139. [Google Scholar]
- Wang, X.; Morris, M.E. Effects of the flavonoid chrysin on nitrofurantoin pharmacokinetics in rats: Potential involvement of ABCG2. Drug Metab. Dispos. 2007, 35, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Benet, L.Z. The gut as a barrier to drug absorption: Combined role of cytochrome P450 3A and P-glycoprotein. Clin. Pharm. 2001, 40, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Cornaire, G.; Woodley, J.F.; Saivin, S.; Legendre, J.Y.; Decourt, S.; Cloarec, A.; Houin, G. Effect of polyoxyl 35 castor oil and Polysorbate 80 on the intestinal absorption of digoxin in vitro. Arzneimittelforschung 2000, 50, 576–579. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, D.M.; Davis, R.B.; Ettner, S.L.; Appel, S.; Wilkey, S.; Van Rompay, M.; Kessler, R.C. Trends in alternative medicine use in the United States, 1990–1997: Results of a follow-up national survey. JAMA 1998, 280, 1569–1575. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y. Traditional Chinese medicine that improve transdermal absorption. J. External Ther. TCM 2001, 10, 48–49. [Google Scholar]
- Li, J.H.; Qin, F.Q.; Yang, P.M. Reversal of multidrug resistance in human K562/ADM cell line by dauricine. J. Dalian Med. Univ. 2002, 24, 94–96. [Google Scholar]
- Zhang, S.; Yang, X.; Morris, M.E. Combined effects of multiple flavonoids on breast cancer resistance protein (ABCG2)-mediated transport. Pharm. Res. 2004, 21, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Y.; Jiang, G.H.; Ma, Y.Y.; Gao, Y. Research advancement on pharmacological effect of Radix Angelicae Dahuricae. Lishizhen Med. Mater. Med. Res. 2009, 20, 625–627. [Google Scholar]
- Wang, M.Y.; Jia, M.R.; Ma, Y.Y.; Li, X.B. Pharmacological effect of four linear Furocoumarins in Radix Angelicae dahuricae. Nat. Prod. Res. Dev. 2010, 22, 485–489. [Google Scholar]
- Abad, M.J.; Heras, B.D.; Silvan, A.M.; Pascual, R.; Bermejo, P.; Rodriguez, B.; Villar, A.M. Effect of furo-coumarins from Cachrys trifida on some macrophage unctions. J. Pharm. Pharmacol. 2001, 53, 1163–1168. [Google Scholar] [CrossRef] [PubMed]
- Ban, H.S.; Lim, S.S.; Suzuki, K.; Jung, S.H.; Lee, S.; Lee, Y.S.; Shin, K.H.; Ohuchi, K. Inhibitory effects of furanocoumarins isolated from the roots of Angelica dahurica on prostaglandin E2 production. Planta Med. 2003, 69, 408–412. [Google Scholar] [PubMed]
- Kim, Y.K.; Kim, Y.S.; Ryu, S.Y. Antiproliferative effect of furanocoumarins from the root of Angelica dahurica on cultured human tumor cell lines. Phytother. Res. 2007, 21, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Meng, L.Z.; Zhou, J.Y.; Fan, X.F.; Luo-, Y.; Zhang, G.W. Imperatorin is responsible for the vasodilatation activity of Angelica Dahurica var. Formosana regulated by nitric oxide in an endothelium-dependent manner. Chin. J. Integr. Med. 2009, 15, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Mammen, J.S.; Kleiner, H.E.; Di Giovanni, J.; Sutter, T.R.; Strickland, P.T. Coumarins are competitive inhibitors of cytochrome P450 1B1, with equal potency for allelic variants. Pharm. Genom. 2005, 15, 183–188. [Google Scholar] [CrossRef]
- Guan, X.J.; Liao, Z.G.; Zhu, W.; Zhao, G.; Lun, Y.; Liang, X. Effects of 10 furanocoumarins in Angelica dahurica on intestinal transport of vincristine. Chin. Tradit. Herb. Drugs 2015, 46, 2117–2121. [Google Scholar]
- Liang, X.L.; Zhu, M.L.; Zhao, G.W.; Liao, Z.G. Effect of furan coumarins from Angelica Dahuricae Radix on intestinal transport absorption of puerarin, paeoniflorin, and vincristine. Chin. Tradit. Herb. Drugs 2015, 46, 1007–1011. [Google Scholar]
- Luo, S.H.; Wang, Z.; Kansara, V.; Pal, D.; Mitra, A.K. Activity of a sodium-dependent vitamin C transporter (SVCT) in MDCK-MDR1 cells and mechanism of ascorbate uptake. Int. J. Pharm. 2008, 358, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Sarvagalla, S.; Singh, V.K.; Ke, Y.Y.; Shiao, H.Y.; Lin, W.H.; Hsieh, H.P.; Hsu, J.T.; Coumar, M.S. Identification of ligand efficient, fragment-like hits from an HTS library: Structure-based virtual screening and docking investigations of 2H- and 3H-pyrazolo tautomers for Aurora kinase A selectivity. J. Comput. Aided Mol. Des. 2005, 29, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Usha, T.; Middha, S.K.; Goyal, A.K.; Karthik, M.; Manoj, D.; Faizan, S.; Goyal, P.; Prashanth, H.; Pande, V. Molecular docking studies of anti-cancerous candidates in Hippophae rhamnoides and Hippophae salicifolia. J. Biomed. Res. 2014, 28, 406–415. [Google Scholar] [PubMed]
- Shityakov, S.; Förster, C. Multidrug resistance protein P-gp interaction with nanoparticles (fullerenes and carbon nanotube) to assess their drug delivery potential: A theoretical molecular docking study. Int. J. Comput. Biol. Drug Des. 2013, 6, 343–357. [Google Scholar] [CrossRef] [PubMed]
- Hui, S.W.; Sen, A. Effect s of lipid packing on polymorphic phase behavior and membrane properties. Proc. Natl. Acad. Sci. USA 1989, 86, 5825–5829. [Google Scholar] [CrossRef] [PubMed]
- Pajeva, I.K.; Wiese, M.; Cordes, H.P.; Seydel, J.K. Membrane interactions of some catamphiphilic drugs and relation to their multidrug-resistance-reversing ability. J. Cancer Res. Clin. Oncol. 1996, 122, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.L.; Zhao, L.J.; Liao, Z.G.; Yin, R.L.; Zhao, G.W.; Cao, Y.C. Effect of Radix Angelicae Dahuricae extract on P-glycoprotein activity in Caco-2 cells. Chin. Hosp. Pharm. J. 2012, 32, 1781–1785. [Google Scholar]
- Hou, X.L.; Takahashi, K.; Tanaka, K.; Tougou, K.; Qiu, F.; Komatsu, K.; Takahashi, K.; Azuma, J. Curcuma drugs and curcumin regulate the expression and function of P-gp in Caco-2 cells in completely opposite ways. Int. J. Pharm. 2008, 358, 224–229. [Google Scholar] [CrossRef] [PubMed]
- The Molecular Structure of the Ligands Was Found from NCBI. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/11415344#section=Top (accessed on 31 July 2015).
- Mei, M.; Zhang, Y.; Ren, J.H.; Xie, D.; Jia, Y.F.; Hu, J.P.; Li, Y.; Dai, J.G.; Chen, X.G. Resistance reversal effect of a novel taxane compound NPB304 and its collaboration with verapamil. Yao Xue Xue Bao 2014, 49, 1279–1288. [Google Scholar] [PubMed]
- Aller, S.G.; Yu, J.; Ward, A.; Weng, Y.; Chittaboina, S.; Zhuo, R.; Harrell, P.M.; Trinh, Y.T.; Zhang, Q.; Urbatsch, I.L.; et al. Structure of p-glycoprotein reveals a molecular basis for poly-specific drug binding. Science 2009, 323, 1718–1722. [Google Scholar] [CrossRef] [PubMed]
- Shityakov, S.; Neuhaus, W.; Dandekar, T.; Förster, C. Analysing molecular polar surface descriptors to predict blood-brain barrier permeation. Int. J. Comput. Biol. Drug Des. 2013, 6, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Shityakov, S.; Förster, C. In silico predictive model to determine vector-mediated transport properties for the blood-brain barrier choline transporter. Adv. Appl. Bioinform. Chem. 2014, 7, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.B.; Fang, P.F.; Cai, H.L.; Yan, M.; Liu, Y.P.; Xu, P.; Zhu, R.H.; Li, H.D. In vitro study on the interactions of P-gp and central depressants. Chin. Hosp. Pharm. J. 2012, 32, 919–923. [Google Scholar]
- Garriga, C.; Vázquez, C.M.; Ruiz-Gutiérrez, V.; Planas, J.M. Regional differences in transport, lipid composition, and fluidity of apical membranes of small intestine of chicken. Poult. Sci. 2002, 81, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Type of Interaction | Key Amino Acids | LibDock Score |
|---|---|---|---|
| Imperatorin | Hydrogen Bond | leu971, phe332 and phe974 | 107 |
| Hydrophobic | ser975, phe71 and phe953 | ||
| Verapamil | r | gly222 | 132 |
| Hydrophobic | ala225 |
| Group | Ivv | Ivh | Anisotropy |
|---|---|---|---|
| Control | 566.923 ± 0.449 | 234.240 ± 0.081 | 0.103 ± 0.001 |
| Imperatorin (10 µg/mL) | 376.096 ± 6.845 | 151.398 ± 3.120 | 0.113 ± 0.002 * |
| Imperatorin (5 µg/mL) | 429.017 ± 2.370 | 174.359 ± 1.395 | 0.109 ± 0.001 * |
| Imperatorin (1 µg/mL) | 481.778 ± 8.710 | 197.003 ± 3.869 | 0.107 ± 0.002 |
| Cholesterol (25 µm) | 826.818 ± 6.157 | 288.311 ± 1.139 | 0.165 ± 0.001 ** |
| Benzyl alcohol (30 mm) | 540.317 ± 7.759 | 240.089 ± 3.142 | 0.077 ± 0.001 ** |
| Group | Dose (µg/mL) | Fluorescence Intensity |
|---|---|---|
| Control | - | 35830 ± 7718 |
| Rh-123 | - | 229466 ± 28022 |
| Verapamil + Rh-123 | 10 | 428521 ± 34018 ** |
| Imperatorin + Rh-123 | 1 | 258163 ± 16538 |
| Imperatorin + Rh-123 | 5 | 294909 ± 35083 * |
| Imperatorin + Rh-123 | 10 | 339324 ± 8778 * |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Z.-G.; Tang, T.; Guan, X.-J.; Dong, W.; Zhang, J.; Zhao, G.-W.; Yang, M.; Liang, X.-L. Improvement of Transmembrane Transport Mechanism Study of Imperatorin on P-Glycoprotein-Mediated Drug Transport. Molecules 2016, 21, 1606. https://doi.org/10.3390/molecules21121606
Liao Z-G, Tang T, Guan X-J, Dong W, Zhang J, Zhao G-W, Yang M, Liang X-L. Improvement of Transmembrane Transport Mechanism Study of Imperatorin on P-Glycoprotein-Mediated Drug Transport. Molecules. 2016; 21(12):1606. https://doi.org/10.3390/molecules21121606
Chicago/Turabian StyleLiao, Zheng-Gen, Tao Tang, Xue-Jing Guan, Wei Dong, Jing Zhang, Guo-Wei Zhao, Ming Yang, and Xin-Li Liang. 2016. "Improvement of Transmembrane Transport Mechanism Study of Imperatorin on P-Glycoprotein-Mediated Drug Transport" Molecules 21, no. 12: 1606. https://doi.org/10.3390/molecules21121606
APA StyleLiao, Z.-G., Tang, T., Guan, X.-J., Dong, W., Zhang, J., Zhao, G.-W., Yang, M., & Liang, X.-L. (2016). Improvement of Transmembrane Transport Mechanism Study of Imperatorin on P-Glycoprotein-Mediated Drug Transport. Molecules, 21(12), 1606. https://doi.org/10.3390/molecules21121606