
Flavonoids, Antioxidant Potential, and Acetylcholinesterase Inhibition Activity of the Extracts from the Gametophyte and Archegoniophore of Marchantia polymorpha L.



Abstract
:1. Introduction
2. Results and Discussion
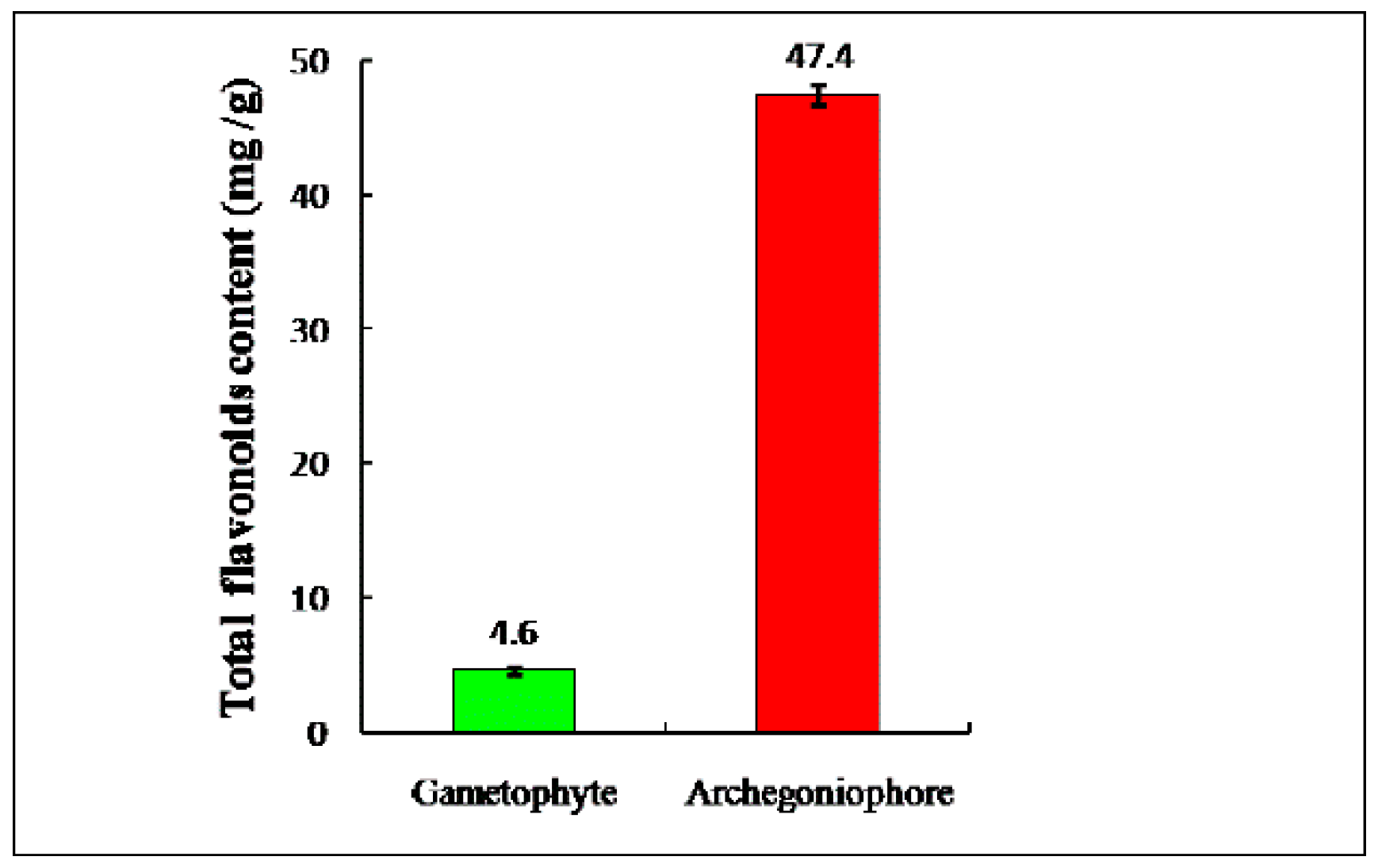
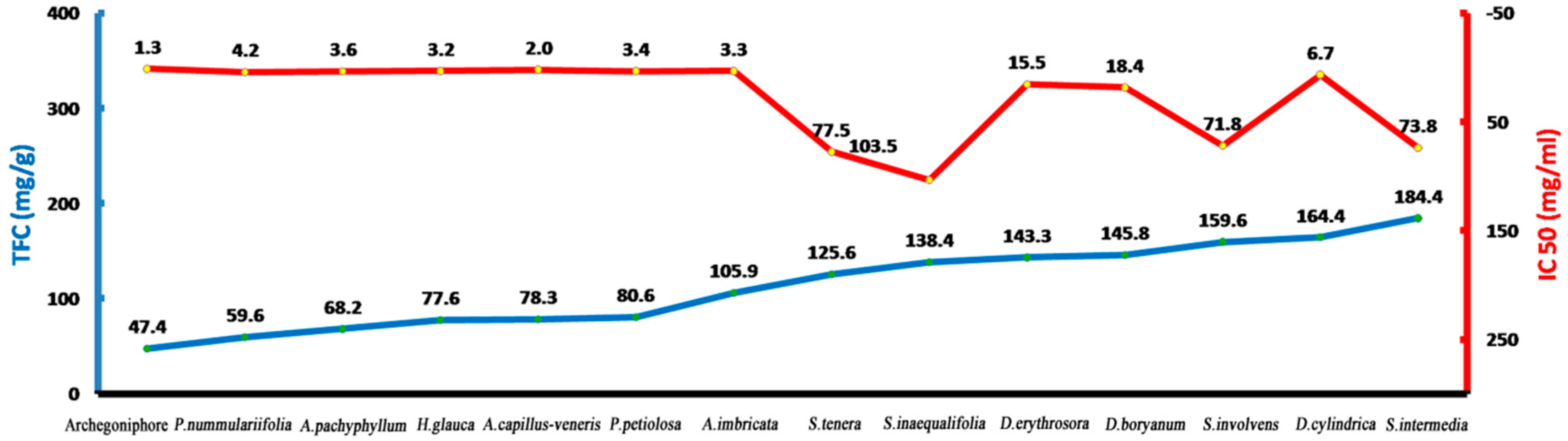
2.1. Total Flavonoid Contents in M. polymorpha L.
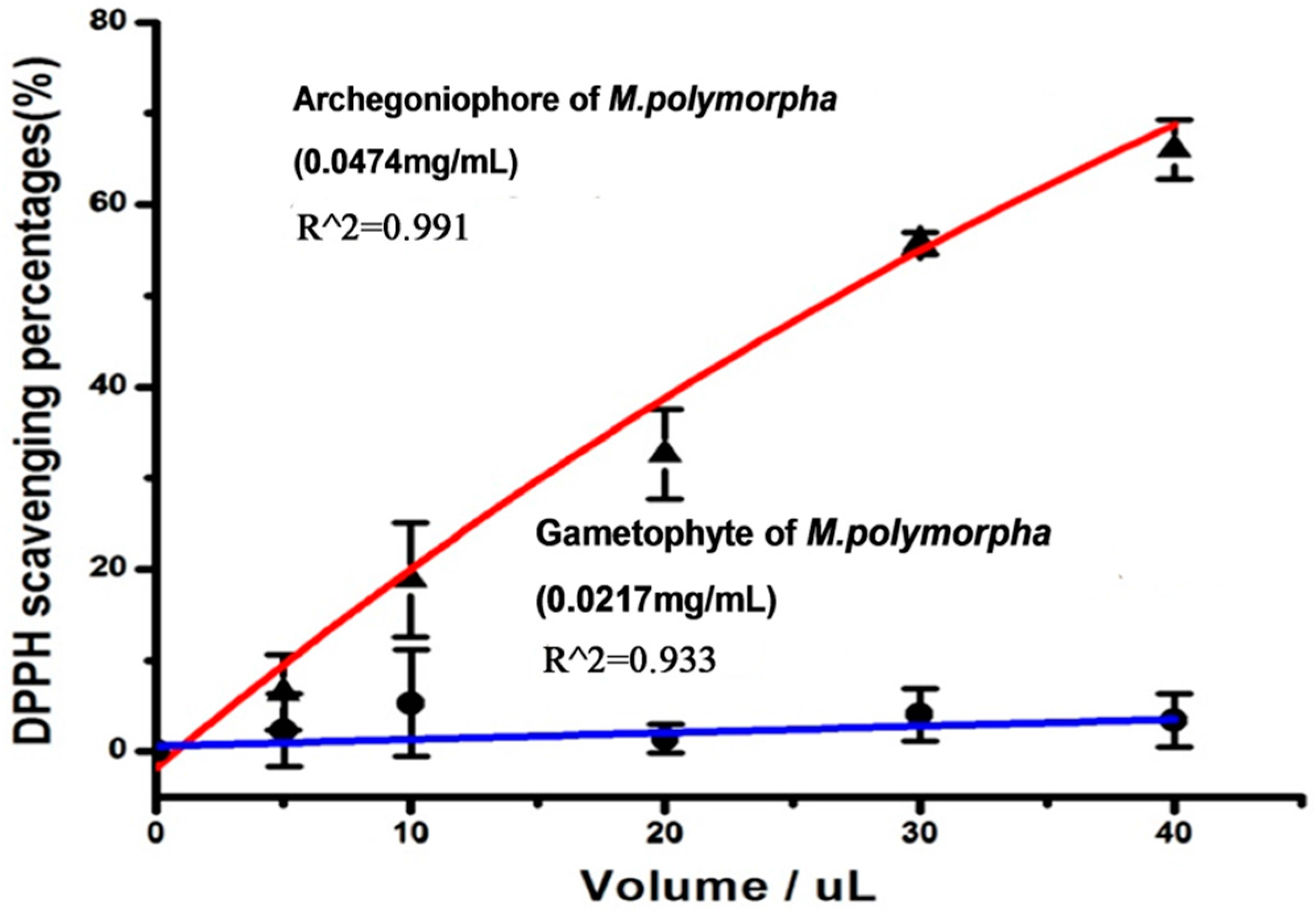
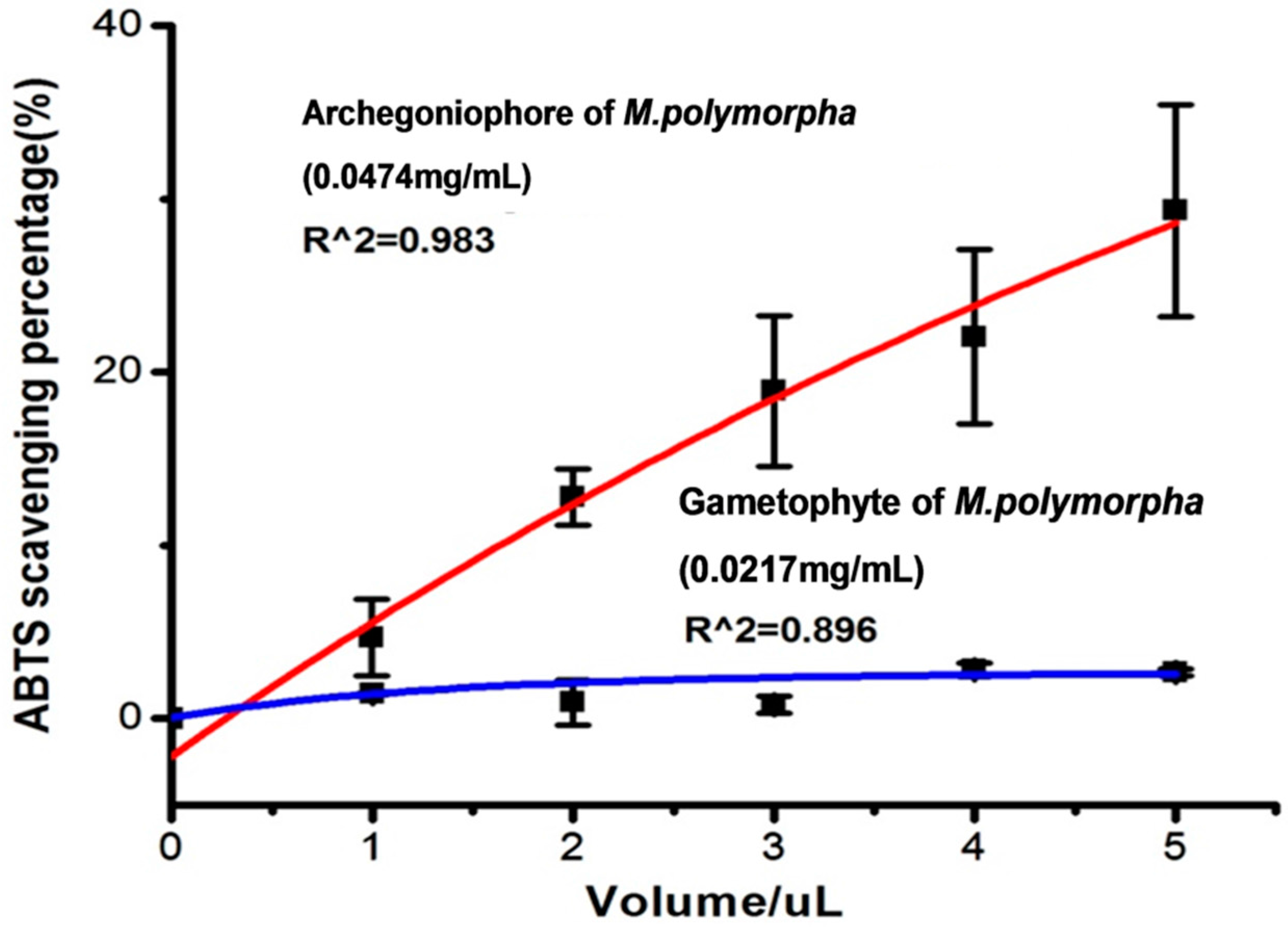
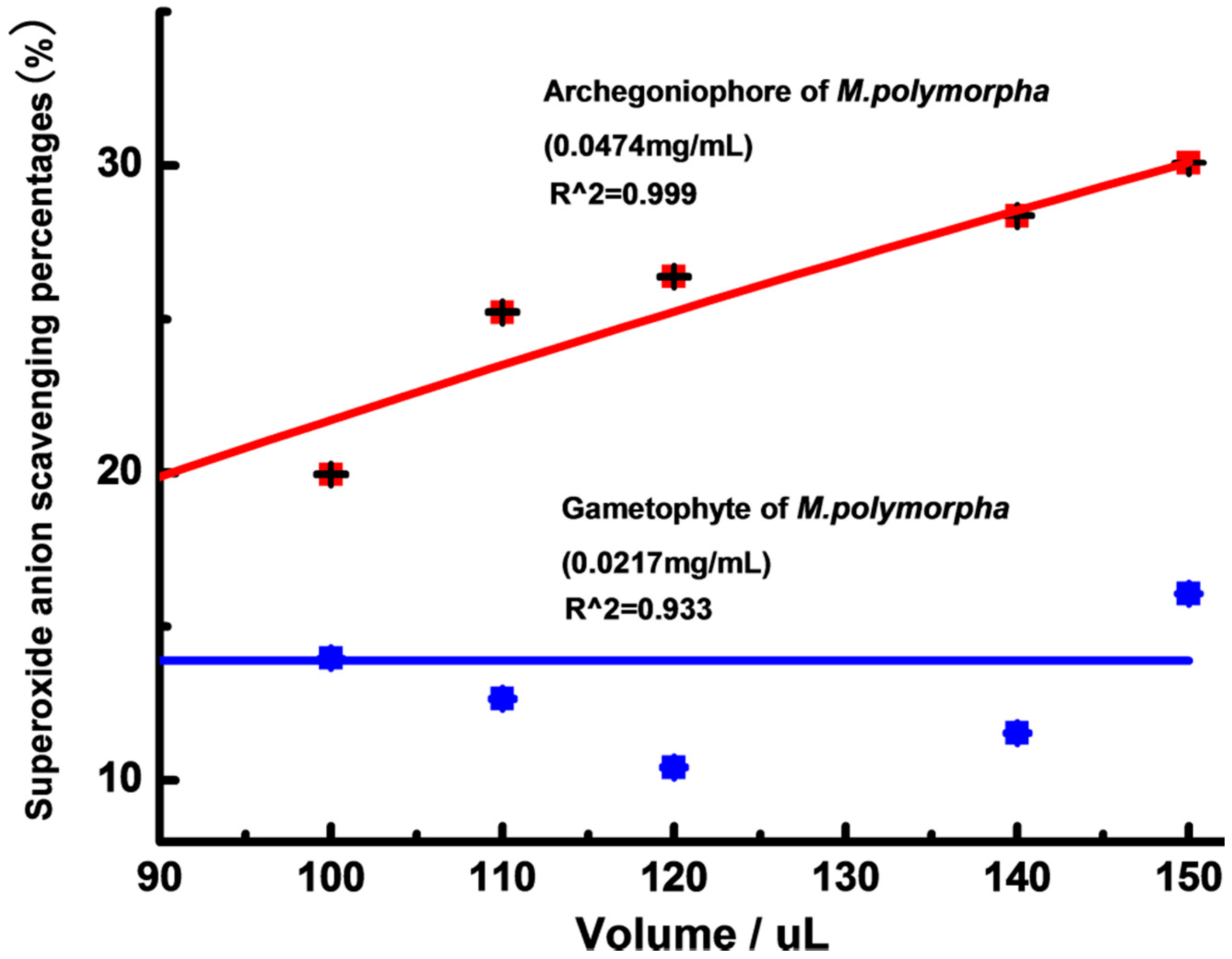
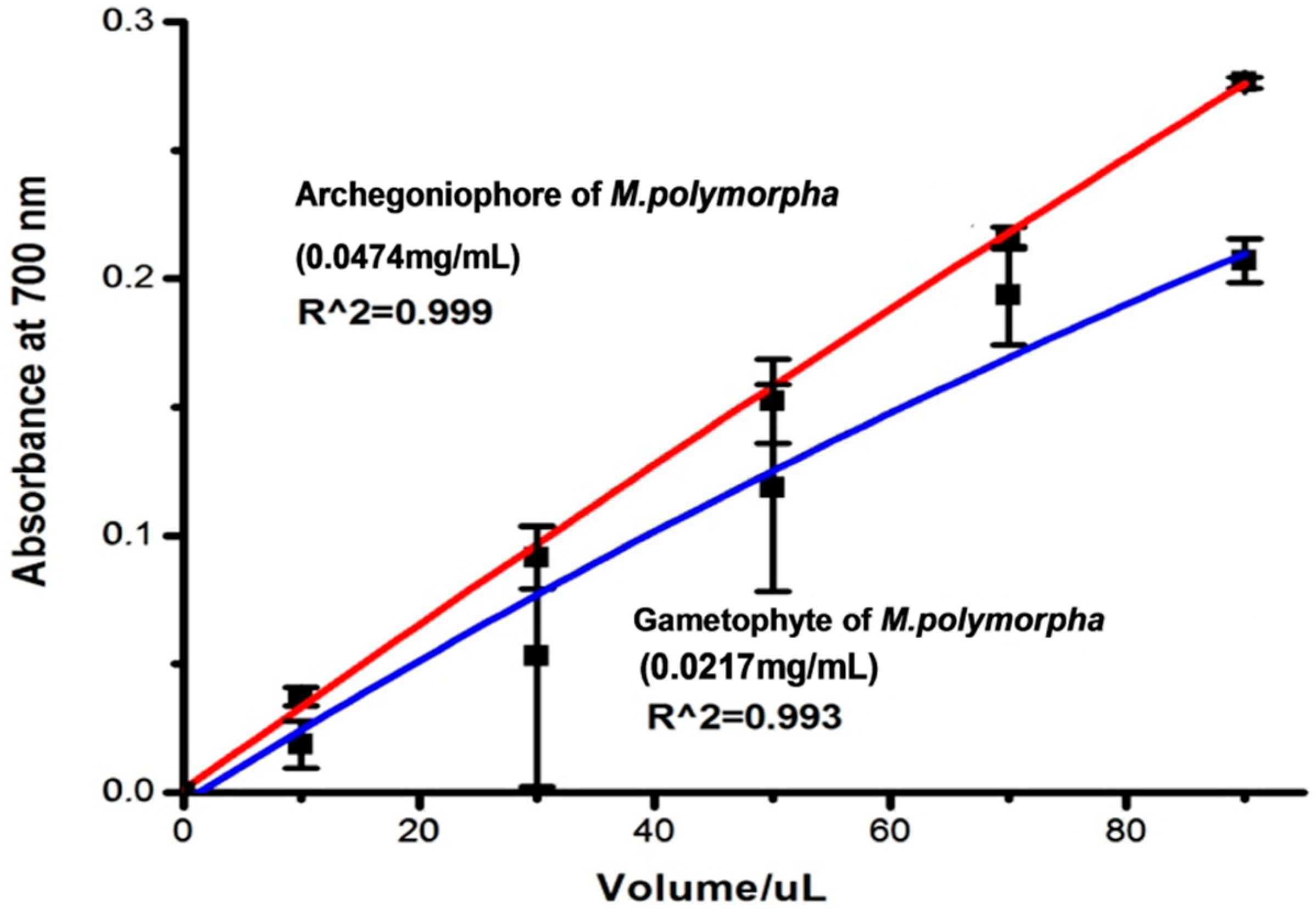
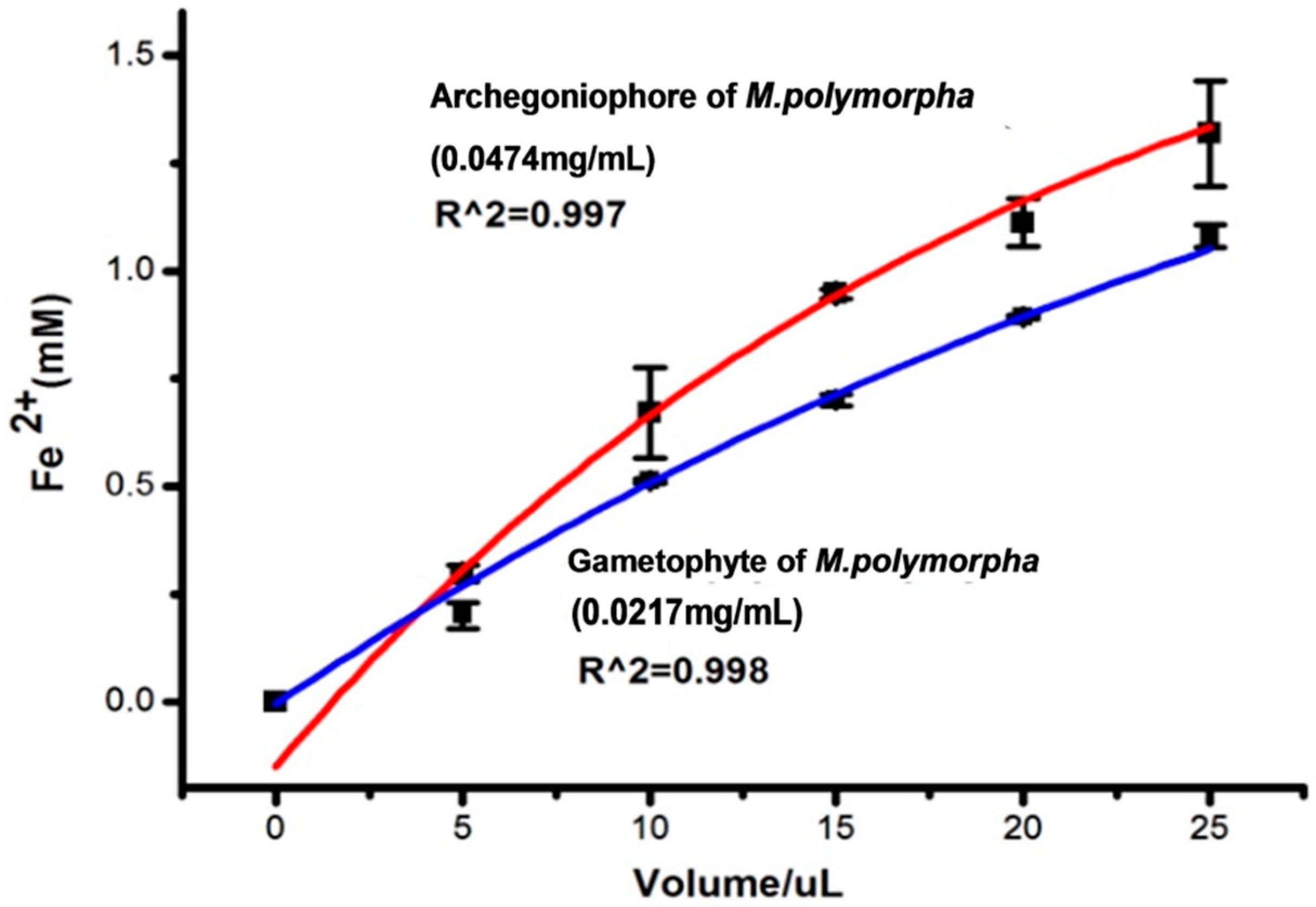
2.2. Antioxidant Activity
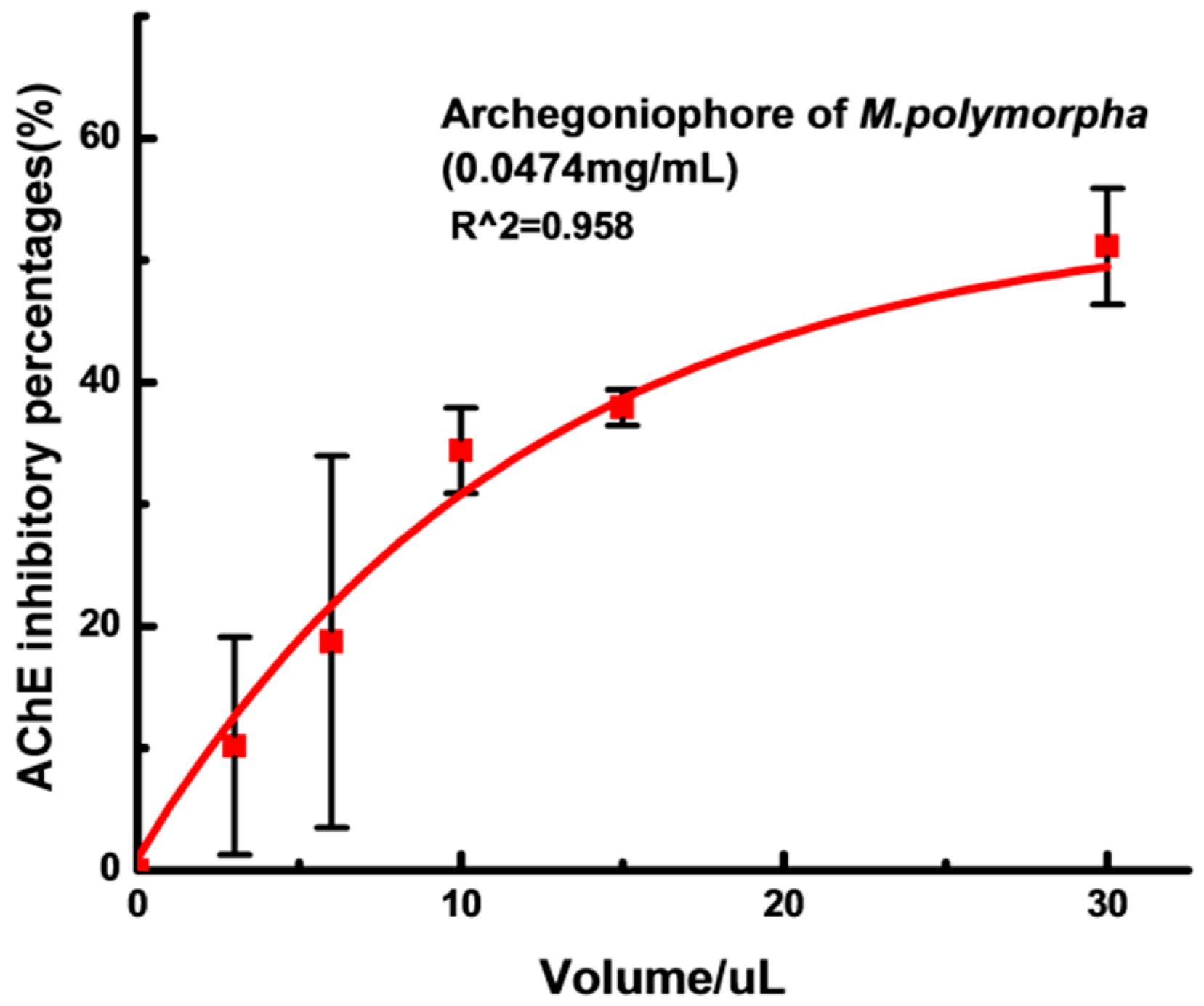
2.3. AChE Inhibition Activity
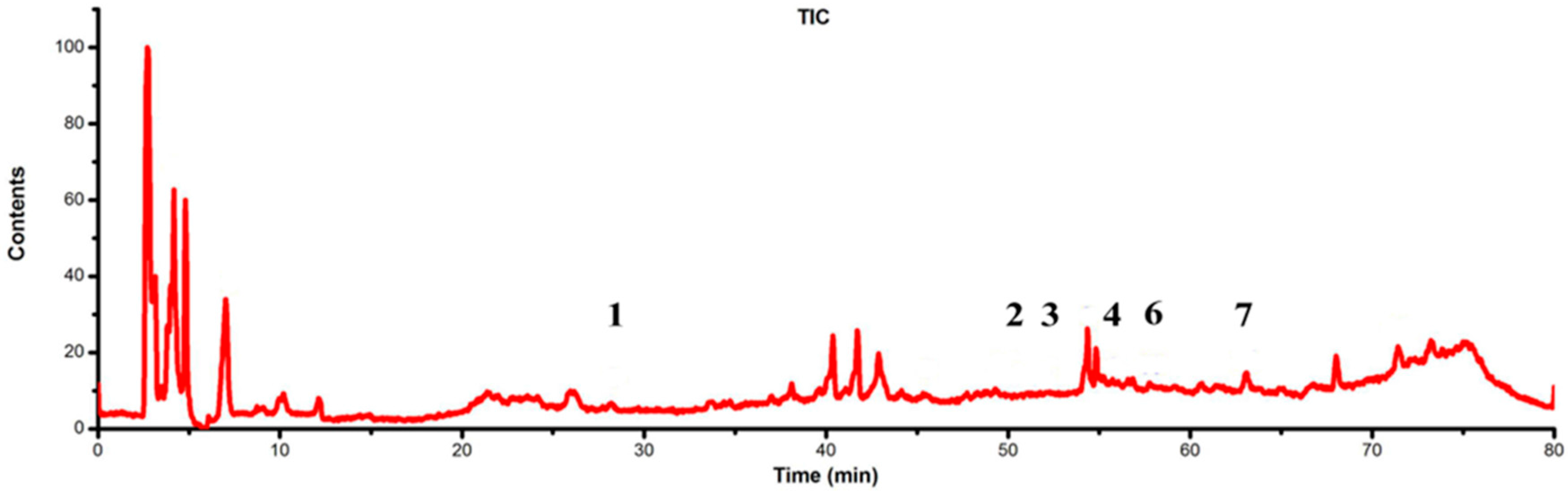
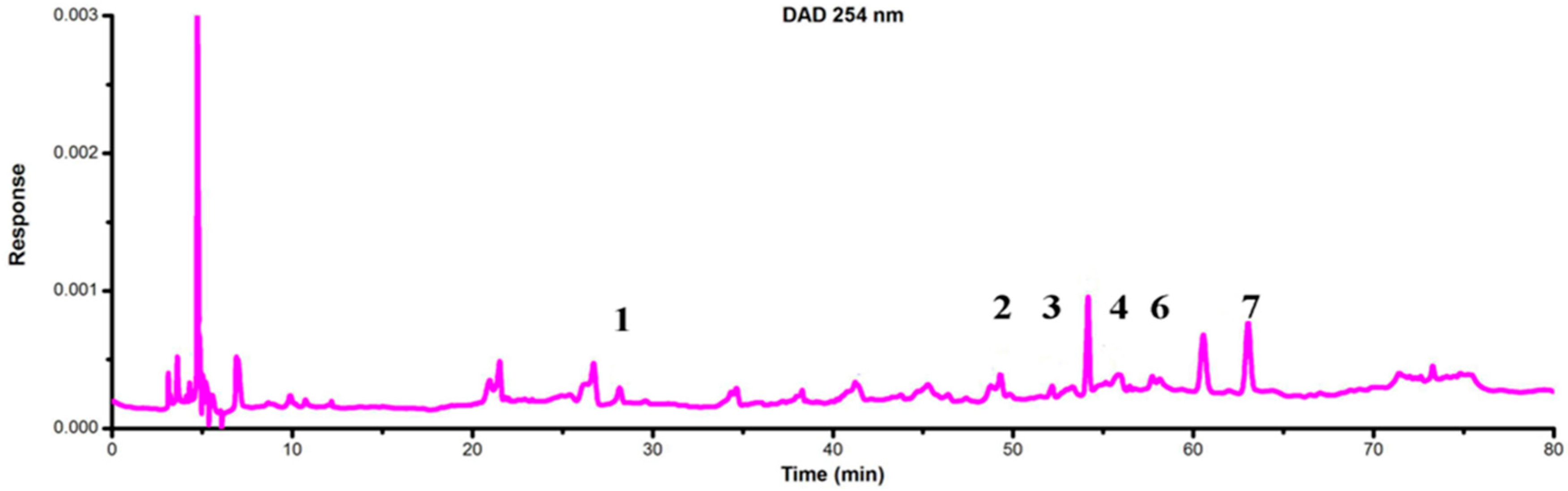
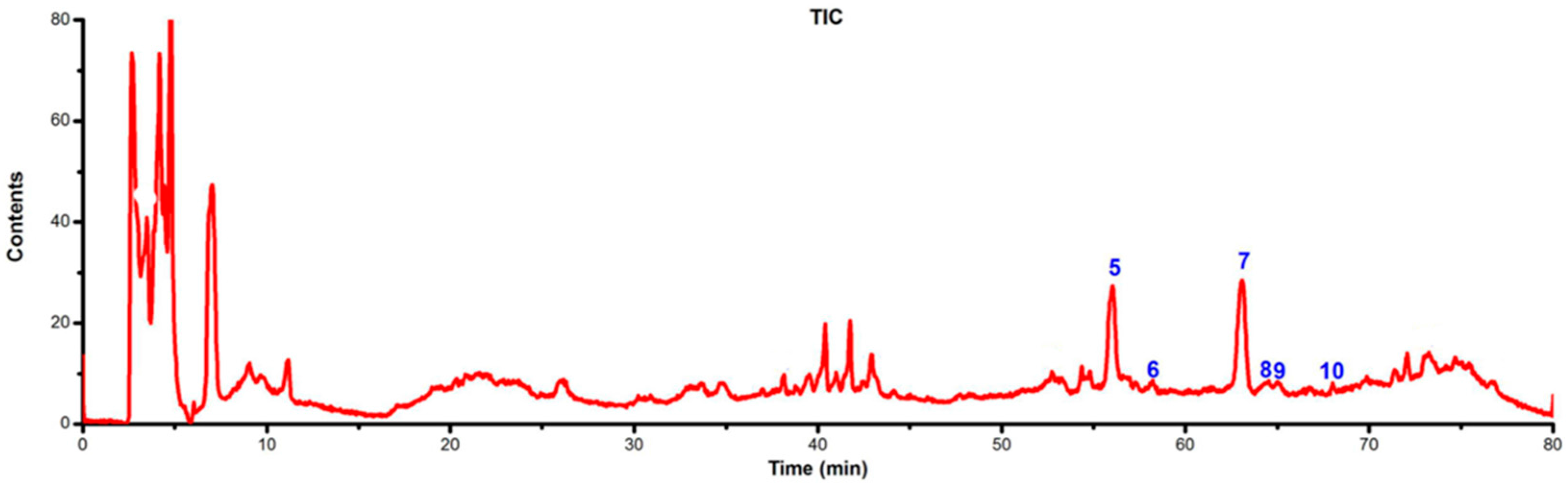
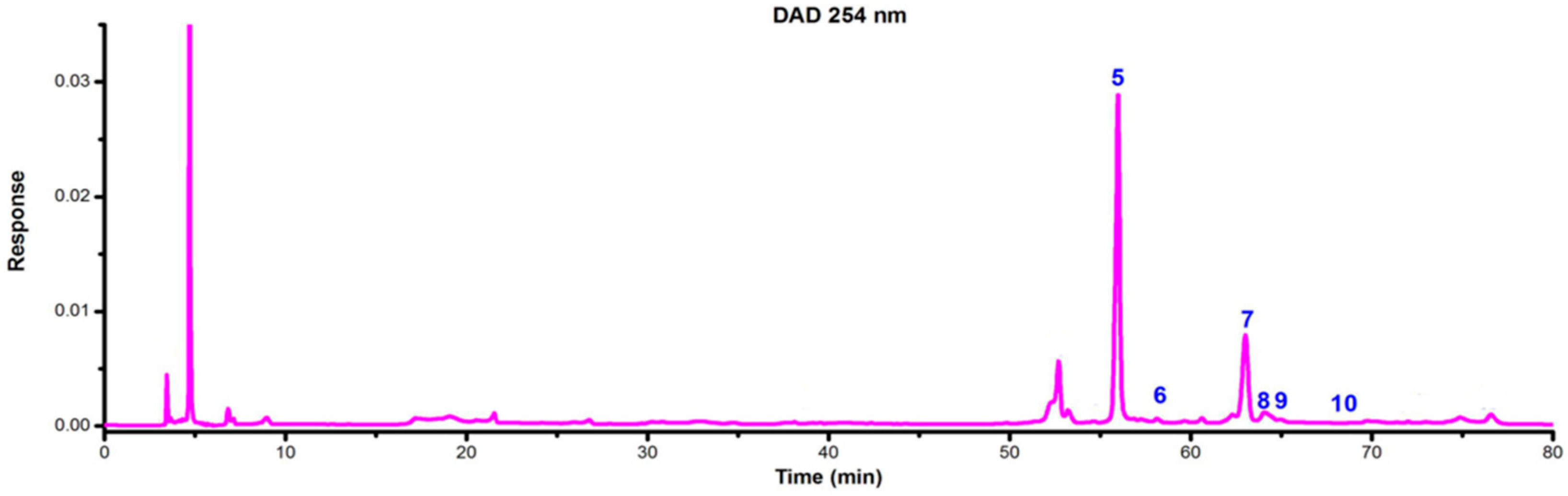
2.4. Identification of Flavonoids in Extracts from the Archegoniophore and Gametophyte of M. polymorpha L.
3. Materials and Methods
3.1. Plant Materials
3.2. Chemicals and Reagents
3.3. Preparation of Plant Extracts
3.4. Determination of Total Flavonoids Content
3.5. Antioxidant Activity
3.5.1. DPPH Assay
3.5.2. ABTS Assay
3.5.3. Reducing Power Assay
3.5.4. Superoxide Anion (O2ˉ) Scavenging Activity
3.5.5. FRAP Assay
3.6. AChE Inhibitory Activity
3.7. LC/DAD/ESI–MS Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Banerjee, R.D.; Sen, S.P. Antibiotic activity of bryophytes. Bryologist 1979, 82, 141–153. [Google Scholar] [CrossRef]
- Asakawa, Y.; Chopra, R.; Bhatla, S. Biologically Active Substances from Bryophytes. Bryophyte Development: Physiology and Biochemistry; Chopra, R.N., Bhatla, S.C., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 259–287. [Google Scholar]
- Harborne, J.B. Phytochemical Methods a Guide to Modern Techniques of Plant Analysis, 3rd ed.; Chapman & Hall: London, UK, 1998; pp. 40–96. [Google Scholar]
- Wu, Y.H.; Yang, H.Y.; Luo, H.; Gao, Q. Resources of medicinal bryophytes in north–eastern China and their exploitation. Chin. J. Ecol. 2004, 23, 218–223. [Google Scholar]
- Xiao, J.B. Natural polyphenols and diabetes: understanding their mechanism of action. Curr. Med. Chem. 2015, 22, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.B.; Muzashvili, T.S.; Georgiev, M.I. Advances in the biotechnological glycosylation of valuable flavonoids. Biotechnol. Adv. 2014, 32, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Manoj, G.S.; Murugan, K. Phenolic profiles, antimicrobial and antioxidant potentiality of methanolic extract of a liverwort, Plagiochila beddomei Steph. IJNPR 2012, 3, 173–183. [Google Scholar]
- Delmas, D.; Xiao, J.B. EDITORIAL (Hot Topic: Natural Polyphenols Properties: Chemopreventive and Chemosensitizing Activities). Anti-Cancer Agent. Med. 2012, 12, 835. [Google Scholar] [CrossRef]
- Xiao, J.B.; Högger, P. Dietary polyphenols and type 2 diabetes: Current insights and future perspectives. Curr. Med. Chem. 2015, 22, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Andrae-Marobela, K.; Ghislain, F.W.; Okatch, H.; Majinda, R.R.T. Polyphenols: A diverse class of multi-target anti-HIV-1 agents. Curr. Drug. Metab. 2013, 14, 392–413. [Google Scholar] [CrossRef] [PubMed]
- Panickar, K.S. Effects of dietary polyphenols on neuroregulatory factors and pathways that mediate food intake and energy regulation in obesity. Mol. Nutr. Food. Res. 2013, 57, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Gahtori, D.; Chaturvedi, P. Antifungal and antibacterial potential of methanol and chloroform extracts of Marchantia polymorpha L. Arch. Phytopathol. Plant. Protect. 2011, 44, 726–731. [Google Scholar] [CrossRef]
- Mewari, N.; Kumar, P. Antimicrobial activity of extracts of Marchantia polymorpha. Pharm. Biol. 2008, 46, 819–822. [Google Scholar] [CrossRef]
- Gokbulut, A.; Satilmis, B.; Batcioglu, K.; Cetin, B.; Sarer, E. Antioxidant activity and luteolin content of Marchantia polymorpha L. Turk. J. Biol. 2012, 36, 381–385. [Google Scholar]
- Asakawa, Y.; Tori, M.; Masuya, T.; Frahm, J.P. Ent-sesquiterpenoids and cyclic bis (bibenzyls) from the German liverwort Marchantia polymorpha. Phytochemistry 1990, 29, 1577–1584. [Google Scholar] [CrossRef]
- Niu, C.; Qu, J.B.; Lou, H.X. Antifungal bis [bibenzyls] from the Chinese liverwort Marchantia polymorpha L. Chem. Biodivers. 2006, 3, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Markham, K.R.; Porter, L.J. Production of an aurone by bryophytes in the reproductive phase. Phytochemistry 1978, 17, 159–160. [Google Scholar] [CrossRef]
- Miksicek, R.J. Commonly occurring plant flavonoids have estrogenic activity. Mol. Pharmacol. 1993, 44, 37–43. [Google Scholar] [PubMed]
- Qin, D.N.; She, B.R.; She, Y.C.; Wang, J.H. Effects of flavonoids from Semen Cuscutae on the reproductive system in male rats. Asian. J. Androl. 2000, 2, 99–102. [Google Scholar] [PubMed]
- Galluzzo, P.; Marino, M. Nutritional flavonoids impact on nuclear and extranuclear estrogen receptor activities. Genes. Nutr. 2006, 1, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Cao, J.G.; Zheng, Y.X.; Wang, Q.X.; Xiao, J.B. Flavonoid concentrations and bioactivity of flavonoid extracts from 19 species of ferns from China. Ind. Crop. Prod. 2014, 58, 91–98. [Google Scholar] [CrossRef]
- Sivaraman, A.; Johnson, M.; Parimelazhagan, T.; Irudayaraj, V. Evaluation of antioxidant potential of ethanolic extracts of selected species of Selaginella. IJNPR 2013, 4, 238–244. [Google Scholar]
- Cao, J.G.; Zheng, Y.X.; Xia, X.; Wang, Q.X.; Xiao, J.B. Total flavonoid contents, antioxidant potential and acetylcholinesterase inhibition activity of the extracts from 15 ferns in China. Ind. Crop. Prod. 2015, 75, 135–140. [Google Scholar] [CrossRef]
- Pourmorad, F.; Hosseinimehr, S.J.; Shahabimajd, N. Antioxidant activity, phenol and flavonoid contents of some selected Iranian medicinal plants. Afr. J. Biotechnol. 2006, 5, 1142–1145. [Google Scholar]
- Pohanka, M. Acetylcholinesterase inhibitors: A patent review (2008-present). Expert. Opin. Ther. Pat. 2012, 22, 871–886. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, A.; Svensson, A.L. Cholinesterase inhibitors in the treatment of Alzheimer’s disease. Drug Saf. 1998, 19, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Cimanga, K.; De, B.T.; Lasure, A.; Li, Q.; Pieters, L.; Claeys, M.; Berghe, D.V.; Kambu, K.; Tona, L.; Vlietinck, A. Flavonoid O-glycosides from the leaves of Morinda morindoides. Phytochemistry 1995, 38, 1301–1303. [Google Scholar] [CrossRef]
- Sánchez-Rabaneda, F.; Jáuregui, O.; Casals, I.; Andrés-Lacueva, C.; Izquierdo-Pulido, M.; Lamuela-Raventós, R.M. Liquid chromatographic/electrospray ionization tandem mass spectrometric study of the phenolic composition of cocoa (Theobroma cacao). J. Mass. Spectrom. 2003, 38, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Chen, X.Y.; Zhong, D.F. Stability of baicalin in biological fluids in vitro. J. Pharm. Biomed. 2005, 39, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Heitz, A.; Carnat, A.; Fraisse, D.; Carnat, A.P.; Lamaison, J.L. Luteolin 3′-glucuronide, the major flavonoid from Melissa officinalis subsp. officinalis. Fitoterapia 2000, 71, 201–202. [Google Scholar] [CrossRef]
- Cheng, H.L.; Zhang, L.J.; Liang, Y.H.; Hsu, Y.W.; Lee, I.J.; Liaw, C.C.; Hwang, S.Y.; Kuo, Y.H. Antiinflammatory and Antioxidant Flavonoids and Phenols from Cardiospermum halicacabum. JTCM 2013, 3, 33–40. [Google Scholar] [PubMed]
- Oliveira, D.M.D.; Siqueira, E.P.; Nunes, Y.R.; Cota, B.B. Flavonoids from leaves of Mauritia flexuosa. Rev. Bras. Farmacogn. 2013, 23, 614–620. [Google Scholar] [CrossRef]
- Markham, K.R.; Porter, L.J. Flavonoids of the liverwort Marchantia polymorpha. Phytochemistry 1974, 13, 1937–1942. [Google Scholar] [CrossRef]
- Xiao, J.B.; Huo, J.L.; Jiang, H.X.; Yang, F. Chemical compositions and bioactivities of crude polysaccharides from tea leaves beyond their useful date. Int. J. Boil. Macromol. 2011, 49, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.G.; Xia, X.; Dai, X.L.; Xiao, J.B.; Wang, Q.X.; Andrae-Marobela, K.; Okatch, H. Flavonoids profiles, antioxidant, acetylcholinesterase inhibition activities of extract from Dryoathyrium boryanum (Willd.) Ching. Food. Chem. Toxicol. 2013, 55, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.B.; Chen, X.; Zhang, L.; Talbot, S.G.; Li, G.C.; Xu, M. Investigation of the Mechanism of Enhanced Effect of EGCG on Huperzine A’s Inhibition of Acetylcholinesterase Activity in Rats by a Multispectroscopic Method. J. Agric. Food. Chem. 2008, 56, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.G.; Xia, X.; Chen, X.F.; Xiao, J.B.; Wang, Q.X. Characterization of flavonoids from Dryopteris erythrosora and evaluation of their antioxidant, anticancer and acetylcholinesterase inhibition activities. Food. Chem. Toxicol. 2013, 51, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | TR (min) | MW | m/z | UVλmax/nm | Identification | Archegoniophore | Gametophyte |
|---|---|---|---|---|---|---|---|
| 1 | 28.25 | 594 | 595.1 [M + H]+ 593.2 [M − H]− | 292 344 | Kaempferol-3-O-rutinoside | none | exist |
| 2 | 49.78 | 608 | 609.2 [M + H]+ 607.1 [M − H]− | 268 300 336 | Chrysoeriol 7-O-neohesperidoside | none | exist |
| 3 | 52.19 | 432 | 433.1 [M + H]+ 431 [M − H]− | 268 288 340 | Apigenin-7-O-β-d-glucoside | none | exist |
| 4 | 55.82 | 622 | 623.1 [M + H]+ 621.1 [M − H]− | 258 330 | Baicalein 6,7-di-O-β-d-glucopyranuronoside | none | exist |
| 5 | 55.87 | 286 | 287 [M + H]+ 285 [M − H]− 571 [2M − H] | 254 348 | Kaempferol | exist | none |
| 6 | 58.19 | 446 | 447.1 [M + H]+ 445 [M − H]− | 268 336 | Apigenin-7-O-β-d-glucuronide | none | exist |
| 7 | 63.03 | 270 | 271 [M + H]+ 269 [M − H]− | 268 338 | Apigenin | none | exist |
| 8 | 64.5 | 300 | 301.1 [M + H]+ 299 [M − H]− | 266 340 | Chrysoeriol | exist | none |
| 9 | 64.98 | 462 | 463 [M + H]+ 461 [M − H]− | 254 348 | Luteolin 3′-O-β-d-glucuronide | exist | none |
| 10 | 68.50 | 638 | 639.1 [M + H]+ 637.1 [M − H]− | 210 266 336 | Tricin-7-O-rutinoside | exist | none |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Cao, J.; Wu, Y.; Wang, Q.; Xiao, J. Flavonoids, Antioxidant Potential, and Acetylcholinesterase Inhibition Activity of the Extracts from the Gametophyte and Archegoniophore of Marchantia polymorpha L. Molecules 2016, 21, 360. https://doi.org/10.3390/molecules21030360
Wang X, Cao J, Wu Y, Wang Q, Xiao J. Flavonoids, Antioxidant Potential, and Acetylcholinesterase Inhibition Activity of the Extracts from the Gametophyte and Archegoniophore of Marchantia polymorpha L. Molecules. 2016; 21(3):360. https://doi.org/10.3390/molecules21030360
Chicago/Turabian StyleWang, Xin, Jianguo Cao, Yuhuan Wu, Quanxi Wang, and Jianbo Xiao. 2016. "Flavonoids, Antioxidant Potential, and Acetylcholinesterase Inhibition Activity of the Extracts from the Gametophyte and Archegoniophore of Marchantia polymorpha L." Molecules 21, no. 3: 360. https://doi.org/10.3390/molecules21030360
APA StyleWang, X., Cao, J., Wu, Y., Wang, Q., & Xiao, J. (2016). Flavonoids, Antioxidant Potential, and Acetylcholinesterase Inhibition Activity of the Extracts from the Gametophyte and Archegoniophore of Marchantia polymorpha L. Molecules, 21(3), 360. https://doi.org/10.3390/molecules21030360