
Expression of Terpenoid Biosynthetic Genes and Accumulation of Chemical Constituents in Valeriana fauriei

 ,
,
Abstract
:1. Introduction
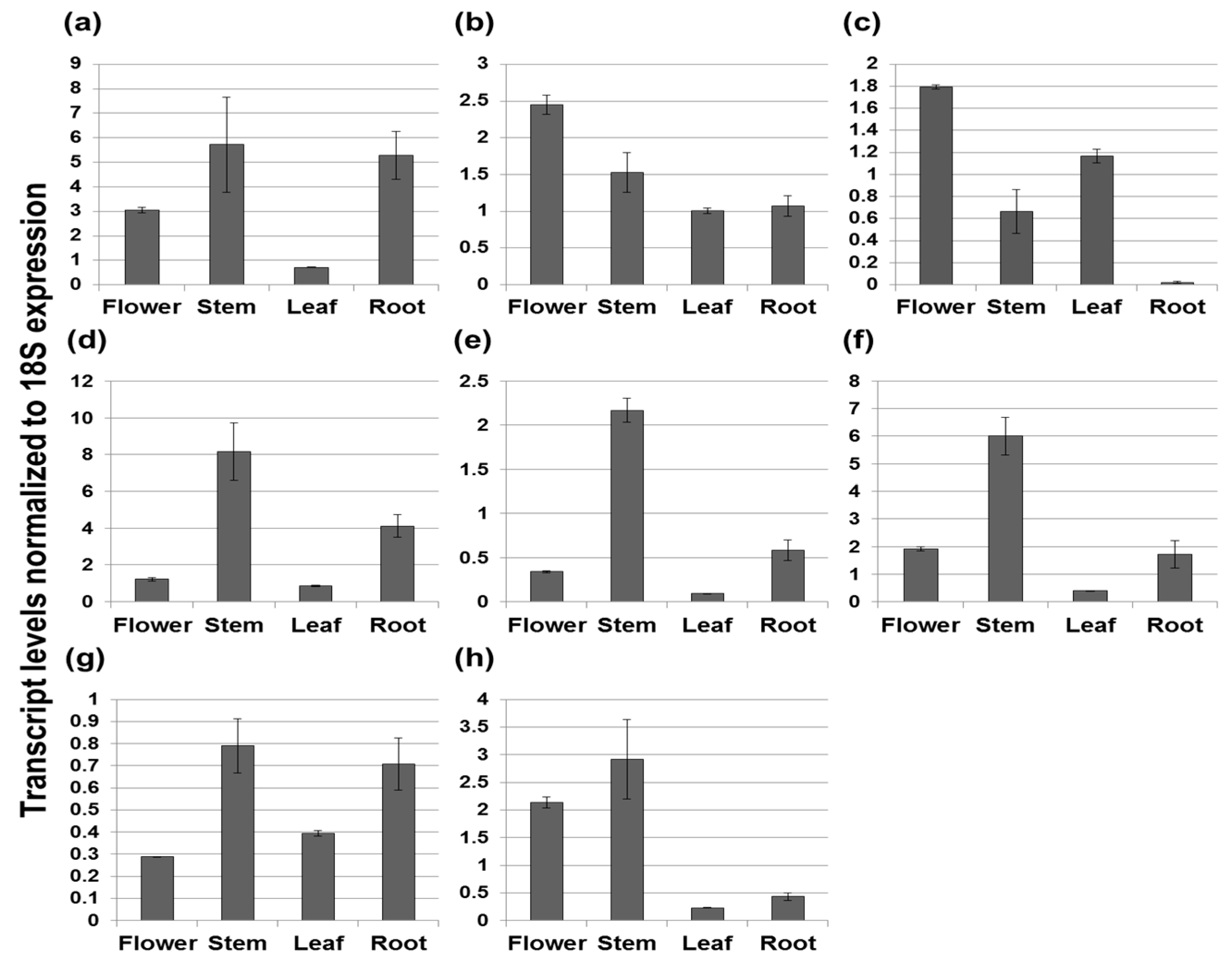
2. Results
2.1. Transcript Levels of Terpenoid Biosynthetic Genes
2.2. Analysis of Volatile Constituents
2.3. Amounts of Valerenic Acid and Its Derivatives
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Total RNA Extraction and cDNA Preparation
4.3. qRT-PCR Analysis
4.4. GC and GC-MS
4.5. Measurement of Valerenic Acid and Derivatives
4.6. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AACT | Acetocetyl-CoA thiolase |
| CAR/PDMS | carboxen/polydimethylsiloxane |
| CMK | 4-(cytidine 5′-diphosphate)-2-C-methyl-d-erythritol kinase |
| DMAPP | dimethylallyl diphosphate |
| DXP | 1-deoxy-d-xylulose 5-phosphate |
| DXR | DXP reductoisomerase |
| DXS | DXP synthase |
| FDS | farndsyl diphosphate synthase |
| FPP | farnesyl diphosphate |
| GA-3P | glyceraldehyde-3-phosphae |
| GC | gas chromatography |
| GC/MS | gas chromatography-mass spectrometry |
| GDS | geranyl diphosphate synthase |
| GGPP | geranyl geranyl diphosphate |
| GPP | geranyl diphosphate |
| HDR | (E)-4-hydroxy-3-methylbut 2-enyl diphosphate reductase |
| HDS | (E)-4-hydroxy-3-methylbut-2-enyl diphosphate synthase |
| HMG-CoA | 3-hydroxy-3-methylglutaryl-CoA |
| HMGR | HMG-CoA reductase |
| HMGS | HMG-CoA synthase |
| IDI | isopentenyl diphosphate isomerase |
| IPP | isopentenyl diphosphate |
| MCT | 2-C-methyl-d-erythritol 4-phosphate cytidylyltransferase |
| MDS | 2-C-methyl-d-erythritol 2,4-cyclodiphosphate synthase |
| MEP | 2-C-methyl-d-erythritol 4-phosphate |
| MK | mevalonate kinase |
| MVA | mevalonae |
| MVD | mevalonate diphosphate decarboxylase |
| qRT-PCR | quantitative Real-time PCR |
| PMK | phosphomevalonate kinase |
| TPS | terpene synthase |
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′ to 3′) | Amplication (bp) | Primer Efficiency (%) |
|---|---|---|---|
| VfAACT-F | ATCGGGCATGAAAGCAACCA | 129 | 97.9 |
| VfAACT-R | GATCCCTTCCTTGCTTCCGCTA | ||
| VfHMGS-F | TGGTGGAACTGCAGCATTGTTC | 131 | 98.8 |
| VfHMGS-R | CCGCACTGTCTGTGCACACGA | ||
| VfHMGR-F | TCAGATGCCCTGCCTCTTCC | 102 | 100.2 |
| VfHMGR-R | GATCTTCTCGCGCCACCTTG | ||
| VfMK-F | CGGCGGCTGCGTATTGACT | 133 | 107.1 |
| VfMK-R | GCAAATCTCGAGCCCATTGC | ||
| VfPMK-F | GAGCCGGAATCACAGACGGA | 146 | 93.3 |
| VfPMK-R | CTCCACGTCTTTGCGAGGCT | ||
| VfMVD-F | AATGGAATCGCGCTGAAGGA | 105 | 98 |
| VfMVD-R | CACAGCATTCGGCCCAGCAT | ||
| VfIDI-F | AGCAGATGCAGGCGAAGAGG | 111 | 93.3 |
| VfIDI-R | GCCTCGCTTAAGTTCCCGTTCT | ||
| VfFDS-F | TGATGACGACGGCAAGGAGA | 131 | 94.9 |
| VfFDS-R | CACCAACCAAGTGAGCATGCAAGA | ||
| VfDXS-F | GCCCAATACCACCTGTCGGA | 121 | 95.8 |
| VfDXS-R | TGCATCGGTCCACCAATCTG | ||
| VfDXR-F | AGAACTCCGGTCATTGTGCCA | 120 | 100.1 |
| VfDXR-R | CCGCCTCGATCTTTGCAAGTTA | ||
| VfMCT-F | TCAGTTGCTCTGCAAATGGGAGT | 104 | 102.2 |
| VfMCT-R | TCCCATTCTTGTGCCCTTTCC | ||
| VfCMK-F | GCACCATTGTTGGGATCGGT | 113 | 82.6 |
| VfCMK-R | CTCGTTCTCGGCTCGTGTGA | ||
| VfMDS-F | TGCAGCTACTGCTGCTGTGGA | 144 | 70.6 |
| VfMDS-R | GGTATGTTGATGCCGCCGAT | ||
| VfHDS-F | CTGACAGGCGGGCACAGTTT | 134 | 97.1 |
| VfHDS-R | GCCGATTCGCATAGCTCTTCC | ||
| VfHDR-F | CCGAAGCAATCGGGAAGTTG | 112 | 95 |
| VfHDR-R | CGCTCTTGAGTAGCGTCGCA | ||
| VfGDS-F | TAGCAGTGCTGGCGGGAGAT | 145 | 102.8 |
| VfGDS-R | TCGCGCGTCGTACTCATTTG |
References
- Pichersky, E.; Noel, J.P.; Dudareva, N. Biosynthesis of plant volatiles: Nature’s diversity and ingenuity. Science 2006, 311, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Ståhl, B. Diversity and distribution of floral scent. Bot. Rev. 2006, 72, 1–120. [Google Scholar] [CrossRef]
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E. Metabolic engineering of plant volatiles. Curr. Opin. Biotechnol. 2008, 19, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Llusià, J. Plant VOC emissions: Making use of the unavoidable. Trends Ecol. Evol. 2004, 19, 402–404. [Google Scholar] [CrossRef] [PubMed]
- Goff, S.A.; Klee, H.J. Plant volatile compounds: Sensory cues for health and nutritional value? Science 2006, 311, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E.; Gertsch, J.; Appendino, G. Plant volatiles: Production, function and pharmacology. Nat. Prod. Rep. 2011, 28, 1359–1380. [Google Scholar] [CrossRef] [PubMed]
- Dicke, M.; Loreto, F. Induced plant volatiles: From genes to climate change. Trends Plant Sci. 2010, 15, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Pichersky, E.; Gershenzon, J. Biochemistry of plant volatiles. Plant Physiol. 2004, 135, 1893–1902. [Google Scholar] [CrossRef] [PubMed]
- Koyama, T.; Ogura, K. Isopentenyl diphosphate isomerase and prenyltransferases. Compr. Nat. Prod. Chem. 1999, 2, 69–96. [Google Scholar]
- Liang, P.H.; Ko, T.P.; Wang, A.H.J. Structure, mechanism and function of prenyltransferases. Eur. J. Biochem. 2002, 269, 3339–3354. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Besser, K.; Harper, A.; Welsby, N.; Schauvinhold, I.; Slocombe, S.; Li, Y.; Dixon, R.A.; Broun, P. Divergent regulation of terpenoid metabolism in the trichomes of wild and cultivated tomato species. Plant Physiol. 2009, 149, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Mathela, C.S.; Chanotiya, C.S.; Sammal, S.S.; Pant, A.K.; Pandey, S. Compositional diversity of terpenoids in the himalayan valeriana genera. Chem. Biodivers. 2005, 2, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jin, L.; Yu, S.; Shi, Q.; Gu, Y.; Kiyota, H. Chemical constituents of plants from the genus valeriana. Mini Rev. Org. Chem. 2010, 7, 161–172. [Google Scholar] [CrossRef]
- Houghton, P.J. The scientific basis for the reputed activity of valerian. J. Pharm. Pharmacol. 1999, 51, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-W.; Wei, B.-J.; He, X.-H.; Liu, Y.; Wang, J. Chemical components and cardiovascular activities of Valeriana spp. Evid. Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef]
- Bos, R.; Woerdenbag, H.J.; Van Putten, F.; Hendriks, H.; Scheffer, J. Seasonal variation of the essential oil, valerenic acid and derivatives, and velopotriates in valeriana officinalis roots and rhizomes, and the selection of plants suitable for phytomedicines. Planta Med. 1998, 64, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Oprean, R.; Oprean, L.; Tamas, M.; Sandulescu, R.; Roman, L. Essential oils analysis. II. Mass spectra identification of terpene and phenylpropane derivatives. J. Pharm. Biomed. Anal. 2001, 24, 1163–1168. [Google Scholar] [CrossRef]
- Dhaubhadel, S.; McGarvey, B.D.; Williams, R.; Gijzen, M. Isoflavonoid biosynthesis and accumulation in developing soybean seeds. Plant Mol. Biol. 2003, 53, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.B.; Thwe, A.A.; Li, X.; Tuan, P.A.; Lee, S.; Lee, J.W.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. Accumulation of astragalosides and related gene expression in different organs of astragalus membranaceus BGE. Var mongholicus (BGE.). Molecules 2014, 19, 10922–10935. [Google Scholar] [CrossRef] [PubMed]
- Lykkesfeldt, J.; Moller, B. Synthesis of benzylglucosinolate in Tropaeolum majus L. (isothiocyanates as potent enzyme inhibitors). Plant Physiol. 1993, 102, 609–613. [Google Scholar] [PubMed]
- Gillissen, B.; Bürkle, L.; André, B.; Kühn, C.; Rentsch, D.; Brandl, B.; Frommer, W.B. A new family of high-affinity transporters for adenine, cytosine, and purine derivatives in arabidopsis. Plant Cell 2000, 12, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, K. Transporters of secondary metabolites. Curr. Opin. Plant Biol. 2005, 8, 301–307. [Google Scholar] [PubMed]
- Nishiya, K.; Kimura, T.; Takeya, K.; Itokawa, H. Sesquiterpenoids and iridoid glycosides from valeriana fauriei. Phytochemistry 1994, 36, 1547–1548. [Google Scholar] [CrossRef]
- Zhang, Z.-X.; Dou, D.-Q.; Liu, K.; Yao, X.-S. Studies on the chemical constituents of valeriana fauriei briq. J. Asian Nat. Prod. Res. 2006, 8, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Nagegowda, D.A. Plant volatile terpenoid metabolism: Biosynthetic genes, transcriptional regulation and subcellular compartmentation. Fed. Eur. Biochem. Soc. 2010, 584, 2965–2973. [Google Scholar] [CrossRef] [PubMed]
- Croteau, R. Biosynthesis and catabolism of monoterpenoids. Chem. Rev. 1987, 87, 929–954. [Google Scholar] [CrossRef]
- Davis, E.M.; Croteau, R. Cyclization enzymes in the biosynthesis of monoterpenes, sesquiterpenes, and diterpenes. In Biosynthesis; Springer: Berlin, Germany, 2000; pp. 53–95. [Google Scholar]
- Adam, K.-P.; Croteau, R. Monoterpene biosynthesis in the liverwort conocephalum conicum: Demonstration of sabinene synthase and bornyl diphosphate synthase in honour of Professor G.H. Neil towers 75th birthday. Phytochemistry 1998, 49, 475–480. [Google Scholar] [CrossRef]
- Navarrete, A.; Avula, B.; Choi, Y.-W.; Khan, I.A. Chemical fingerprinting of valeriana species: Simultaneous determination of valerenic acids, flavonoids, and phenylpropanoids using liquid chromatography with ultraviolet detection. J. AOAC Int. 2006, 89, 8–15. [Google Scholar] [PubMed]
- Park, Y.J.; Li, X.; Noh, S.J.; Kim, J.K.; Lim, S.S.; Park, N.I.; Kim, S.; Kim, Y.B.; Kim, Y.O.; Lee, S.W. Transcriptome and metabolome analysis in shoot and root of valeriana fauriei. BMC Genom. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not Available.




| No. | Volatile Compounds | RI * | Relative Peak Area (%) | |||
|---|---|---|---|---|---|---|
| Flower | Stem | Leaf | Root | |||
| 1 | n-Hexanal | 1.43 | 0 | 0 | 0 | 0.162 ± 0.004 |
| 2 | Isovaleric acid | 11.96 | 0 | 0 | 0 | 3.338 ± 0.082 |
| 3 | Camphene | 12.91 | 0 | 0 | 0 | 3.338 ± 0.082 |
| 4 | β-Pinene | 13.64 | 0 | 1.244 ± 0.103 | 0 | 0.173 ± 0.0041 |
| 5 | β-Terpinene | 14.51 | 0 | 0 | 0 | 0.558 ± 0.014 |
| 6 | Myrcene | 15.44 | 0 | 0 | 0 | 0.465 ± 0.011 |
| 7 | p-Cymene | 17.22 | 0 | 0 | 0 | 0.085 ± 0.002 |
| 8 | β-Phellandrene | 17.79 | 1.277 ± 0.111 | 0 | 0 | 0.803 ± 0.020 |
| 9 | γ-Terpinene | 19.09 | 0.132 ± 0.011 | 0 | 3.231 ± 0.262 | 0.052 ± 0.001 |
| 10 | Terpinolene | 21.35 | 0 | 0.901 ± 0.074 | 0 | 0.041 ± 0.001 |
| 11 | Ethyl 3-Hydroxymandelate | 21.54 | 0.197 ± 0.017 | 0.699 ± 0.058 | 5.331 ± 0.433 | 0 |
| 12 | d-Limonene | 22.90 | 0.282 ± 0.024 | 0 | 0 | 0.082 ± 0.002 |
| 13 | 1,3-Bis-(p-carbamoylmethylphenoxy)-2-propanol | 23.18 | 0.512 ± 0.044 | 0 | 0 | 0 |
| 14 | endo-Borneol | 23.60 | 0 | 0.125 ± 0.010 | 11.482 ± 0.932 | 2.841 ± 0.069 |
| 15 | 1-(4-methoxyphenyl)imidazzoline-2-thione | 24.22 | 0.564 ± 0.049 | 0.211 ± 0.017 | 0 | 0 |
| 16 | 2,2-Dimethyl-3-heptanone | 24.41 | 0 | 0 | 0.561 ± 0.046 | 0 |
| 17 | 1,2,4-Trizol-3-amene | 24.87 | 0.179 ± 0.016 | 0 | 0 | 0 |
| 18 | Borneol | 25.11 | 0 | 0 | 0 | 22.96 ± 0.561 |
| 19 | Benzoic acid | 25.35 | 0 | 0.966 ± 0.080 | 0 | 1.522 ± 0.037 |
| 20 | Benzothialzole | 25.87 | 2.725 ± 0.237 | 0 | 0 | 0 |
| 21 | Eudesmol | 25.95 | 0 | 0.354 ± 0.029 | 0 | 0.077 ± 0.002 |
| 22 | 2-Isopropyl-5-methyl anisole | 26.47 | 0.987 ± 0.086 | 0.497 ± 0.041 | 0 | 0 |
| 23 | Myrtenyl acetate | 26.73 | 0 | 0 | 2.226 ± 0.181 | 1.729 ± 0.042 |
| 24 | 2-Hexanoylfuran | 27.27 | 0.610 ± 0.053 | 0 | 0.556 ± 0.045 | 0.132 ± 0.003 |
| 25 | Perillaldehyde | 27.57 | 0.834 ± 0.072 | 0 | 0 | 1.071 ± 0.026 |
| 26 | 9-oxo-(2,6-dimethylpehenyl)amide 9-H-Fluorene-4-carboxylic acid | 27.94 | 1.067 ± 0.093 | 0.241 ± 0.020 | 21.436 ± 1.741 | 0.581 ± 0.014 |
| 27 | Acetic acid | 28.31 | 0.612 ± 0.053 | 0 | 0.353 ± 0.029 | 0.86 ± 0.021 |
| 28 | α-Gurjunene | 29.74 | 0.640 ± 0.056 | 0 | 0 | 0.133 ± 0.003 |
| 29 | Bornyl acetate | 30.51 | 0.132 ± 0.011 | 0 | 0 | 19.923 ± 0.487 |
| 30 | 9H-Fluorene-4-carboxylic acid | 30.77 | 0.266 ± 0.023 | 0.306 ± 0.025 | 0.253 ± 0.021 | 0.087 ± 0.002 |
| 31 | α-Elemene | 30.88 | 0 | 0 | 0 | 0.038 ± 0.001 |
| 32 | 1-Trifluoromethyl-4-(2-emthoxylbenzyloxy)-3-nitro-benzene | 31.06 | 0.298 ± 0.026 | 0.078 ± 0.006 | 0 | 0.029 ± 0.001 |
| 33 | N-(Cyclohexanecarbonyl)-l-proline isobutyl ester | 31.35 | 0.133 ± 0.012 | 0 | 0 | 0.022 ± 0.001 |
| 34 | Bromopropylate | 31.97 | 0.461 ± 0.040 | 0.367 ± 0.030 | 0 | 0.318 ± 0.008 |
| 35 | Methadone N-oxide | 32.50 | 0.286 ± 0.025 | 0 | 2.524 ± 0.205 | 0.509 ± 0.012 |
| 36 | Quinoline | 33.01 | 0.191 ± 0.017 | 0.174 ± 0.014 | 1.510 ± 0.123 | 1.35 ± 0.033 |
| 37 | 2-Chloro-6-methyl-pyridine | 33.40 | 0.190 ± 0.016 | 0.226 ± 0.019 | 0.456 ± 0.037 | 0.065 ± 0.002 |
| 38 | Pentadecane | 33.51 | 0.170 ± 0.015 | 0 | 0 | 0.095 ± 0.002 |
| 39 | 4,6-bis(1-Methylethyl)-trans-1,3-dioxane | 34.00 | 0 | 0.074 ± 0.006 | 0.751 ± 0.061 | 0.065 ± 0.002 |
| 40 | o-Choloroaniline | 34.12 | 0 | 0.955 ± 0.079 | 1.886 ± 0.153 | 0 |
| 41 | α-Caryophyllene alcohol | 34.72 | 0 | 0 | 0 | 1.272 ± 0.031 |
| 42 | Isolonhifolan-8-ol | 34.81 | 0.8410.073 | 0 | 0.424 ± 0.034 | 0.209 ± 0.005 |
| 43 | γ-Elemene | 34.93 | 0 | 0 | 0 | 0.357 ± 0.009 |
| 44 | Caryophyllene | 35.22 | 0.680 ± 0.059 | 0 | 0.205 ± 0.017 | 0.134 ± 0.003 |
| 45 | 5-(Phenylmethyl)-2-thioxo-4-imidazolidinone | 35.37 | 0 | 2.235 ± 0.0185 | 0.434 ± 0.035 | 0 |
| 46 | α-Acorenol | 36.04 | 0.284 ± 0.025 | 0 | 0 | 4.025 ± 0.098 |
| 47 | cis-β-Farnesene | 36.18 | 0 | 0 | 0 | 0.041 ± 0.001 |
| 48 | Humulene | 36.28 | 0 | 0 | 0 | 0.085 ± 0.002 |
| 49 | Aromadendrene | 36.56 | 0 | 0 | 0 | 0.741 ± 0.018 |
| 50 | Alloaromandendrene | 36.71 | 1.162 ± 0.101 | 0 | 0 | 0.066 ± 0.002 |
| 51 | Ylangene | 36.92 | 0 | 0.299 ± 0.025 | 0.779 ± 0.063 | 2.878 ± 0.070 |
| 52 | 1-(1,5-Dimethyl-4-hexenyl)-4-methyl-benzene | 37.13 | 1.181 ± 0.102 | 0.297 ± 0.025 | 0 | 0.065 ± 0.002 |
| 53 | β-Ionone | 37.39 | 0.554 ± 0.048 | 0.179 ± 0.015 | 0.922 ± 0.075 | 2.333 ± 0.057 |
| 54 | Pentadecane | 37.62 | 0 | 0.523 ± 0.043 | 0.439 ± 0.036 | 1.786 ± 0.044 |
| 55 | N-(1H-1,3-Benzimidazol-2-ylmethyl)-4-methoxy-benzamide | 38.31 | 1.646 ± 0.143 | 0.047 ± 0.004 | 0.629 ± 0.051 | 0.475 ± 0.012 |
| 56 | Malonic acid | 38.50 | 0 | 0 | 1.177 ± 0.096 | 3.118 ± 0.076 |
| 57 | β-Bisabolene | 38.81 | 0 | 0.158 ± 0.013 | 0.119 ± 0.010 | 0.54 ± 0.013 |
| 58 | cis-Sesquisabinene | 38.87 | 2.302 ± 0.200 | 0.640 ± 0.053 | 2.530 ± 0.0205 | 0.527 ± 0.013 |
| 59 | N-Phenyl-(3-methyl-2-oxiranyl) methyl carbamate | 39.03 | 0 | 0.362 ± 0.030 | 0 | 0.287 ± 0.007 |
| 60 | Bornyl isovalerate | 39.20 | 1.110 ± 0.096 | 0.143 ± 0.012 | 0.674 ± 0.055 | 0.216 ± 0.005 |
| 61 | exo-3-Methyl-1,7,7-trimethylbicyclo[2,2,1]hept-2-yl butanoate | 39.50 | 0 | 0.143 ± 0.012 | 0 | 2.464 ± 0.060 |
| 62 | 2.6-Dimethylnon-1-en-3-yn-5-yl valeric acid | 39.81 | 0.708 ± 0.061 | 0 | 0 | 1.249 ± 0.031 |
| 63 | cis-α-Bisabolene | 40.33 | 0.621 ± 0.054 | 0 | 0.533 ± 0.043 | 0.187 ± 0.005 |
| 64 | trans-Sesquisabinene | 40.50 | 0 | 0 | 0.467 ± 0.038 | 0.213 ± 0.005 |
| 65 | 1-Phenylthio-3-(1-piperidyl)-propan-2ol | 40.61 | 0.894 ± 0.078 | 0 | 0 | 0.134 ± 0.003 |
| 66 | 6-epi-Shyobunol | 40.79 | 0 | 0.139 ± 0.011 | 0 | 2.376 ± 0.058 |
| 67 | Photocitral B | 40.97 | 0.779 ± 0.068 | 0.356 ± 0.029 | 2.119 ± 0.0172 | 0.282 ± 0.007 |
| 68 | N-(4-Methoxyphenyl)-propanamide | 41.42 | 1.247 ± 0.019 | 0.339 ± 0.028 | 1.212 ± 0.098 | 2.537 ± 0.062 |
| 69 | Menthyl acetate | 41.68 | 0 | 0.254 ± 0.021 | 0.901 ± 0.073 | 0.08 ± 0.002 |
| 70 | (−)-Isolongifolol methyl ether | 42.09 | 0 | 0 | 0 | 0.207 ± 0.005 |
| 71 | 5,5-Dimethyl-4-(3-methyl-1,3-butadienyl)-1-oxaspiro[2,5]octane | 42.20 | 1.121 ± 0.097 | 0 | 0 | 0.127 ± 0.003 |
| 72 | 3,3-dichlorodihydro-2(3H)-furanone | 42.38 | 0 | 0 | 0 | 0.313 ± 0.008 |
| 73 | Z-9-Pentadecenol | 42.74 | 2.454 ± 0.213 | 1.530 ± 0.126 | 3.563 ± 0.289 | 0.544 ± 0.013 |
| 74 | Acetonylacetone dioxime | 42.97 | 0 | 0.122 ± 0.010 | 0 | 0 |
| 75 | 1-Adamantylmethyl 3-methyl-2-butenoate | 43.27 | 1.593 ± 0.138 | 1.392 ± 0.115 | 0.223 ± 0.018 | 0.221 ± 0.005 |
| 76 | 3-(Methylthio)phenyl isothiocyanate | 43.89 | 0 | 0.527 ± 0.044 | 0.921 ± 0.075 | 0.041 ± 0.001 |
| 77 | 1-Adamantylmethyl octanoic acid | 44.20 | 0.780 ± 0.068 | 11.040 ± 0.912 | 0 | 0.123 ± 0.003 |
| 78 | 2-(Methylthio)-benzothialzole | 44.43 | 1.013 ± 0.088 | 0.300 ± 0.025 | 0.280 ± 0.023 | 0.06 ± 0.001 |
| 79 | 3-Methyl-2(3H)-benzothiazolethione | 44.74 | 0.596 ± 0.052 | 0.439 ± 0.036 | 0.654 ± 0.053 | 0.046 ± 0.001 |
| 80 | Pentanoic acid | 45.25 | 1.072 ± 0.09 | 0.531 ± 0.044 | 0.522 ± 0.042 | 0.165 ± 0.004 |
| 81 | Terpinyl acetate | 45.42 | 1.550 ± 0.135 | 3.000 ± 0.248 | 0 | 0 |
| 82 | p-Methoxybenzylazidoformate | 46.29 | 0.975 ± 0.085 | 2.492 ± 0.206 | 1.051 ± 0.085 | 0.439 ± 0.011 |
| 83 | 5-Amino-ethyl ester [1,2,4]triazolo[4.3-a]pyrimidine-6-carboxylate | 46.47 | 0.611 ± 0.053 | 0.288 ± 0.024 | 0 | 0 |
| 84 | 1,3,-Trimethyl-2-hydroxymethyl-3,3-dimethyl-4(3-methylbut-2-eny)-cyclohexene | 46.71 | 1.044 ± 0.091 | 0.766 ± 0.063 | 0 | 0 |
| 85 | 5-Butyl-6-hexyloctahydro-1H-indene | 47.14 | 0.992 ± 0.086 | 0.229 ± 0.019 | 0 | 0 |
| 86 | 4-Hydroxy-2-hydroxymethyl-6-methylpyrimidine | 47.47 | 0.456 ± 0.040 | 0 | 0 | 0 |
| 87 | Murolan-3,9(11)-diene-10-peroxy | 47.76 | 0.888 ± 0.077 | 0.355 ± 0.029 | 0.482 ± 0.039 | 0.131 ± 0.003 |
| 88 | Methanone | 48.04 | 0.414 ± 0.036 | 0.2750.023 | 0 | 0 |
| 89 | 8-Chlorooctyl isobutyl carbonate | 48.29 | 0.522 ± 0.045 | 0.134 ± 0.011 | 0 | 0 |
| 90 | 1,2-Pentanediol | 48.72 | 0.719 ± 0.062 | 0 | 0 | 0.031 ± 0.001 |
| 91 | Bicyclogermacrene | 49.21 | 1.642 ± 0.143 | 0 | 0 | 0 |
| 92 | Larixone | 49.39 | 0 | 0.159 ± 0.013 | 0 | 0 |
| 93 | Ursane-3,16-diol | 49.70 | 0.856 ± 0.074 | 0.921 ± 0.076 | 0.640 ± 0.052 | 0.04 ± 0.001 |
| 94 | Dodecahydro-3,8,8,11a-tetramethyl-5H-3,5a-epoxynaphth[2,1-c]oxepin | 49.94 | 1.248 ± 0.108 | 0.354 ± 0.029 | 0 | 0 |
| 95 | Hexahydro-5-methyl-1-phenyl-1,3,5-triazine-2-thione | 50.27 | 0 | 1.515 ± 0.125 | 0.240 ± 0.019 | 0 |
| 96 | (2R,4R)-p-Mentha-6,8-diene-2-hydroperoxide | 50.71 | 0.793 ± 0.069 | 0.271 ± 0.022 | 0 | 0 |
| 97 | 4-(Diethoxyphosphiniyl)butanoic acid | 51.18 | 0.259 ± 0.022 | 0.246 ± 0.020 | 0 | 0 |
| 98 | 2,2-Dimethylpropanoic acid | 51.74 | 0.878 ± 0.076 | 0.076 ± 0.006 | 0 | 0 |
| 99 | Nerolidol isobutyrate | 52.17 | 0.587 ± 0.051 | 0.253 ± 0.021 | 0 | 0 |
| 100 | Longifolenaldehyde | 52.31 | 0.510 ± 0.044 | 0 | 0 | 0 |
| 101 | Caryophyllene oxide | 52.67 | 0.359 ± 0.031 | 17.454 ± 1.442 | 0 | 0 |
| 102 | Carbamic acid | 52.89 | 0.670 ± 0.058 | 0.939 ± 0.078 | 0.220 ± 0.018 | 0.061 ± 0.001 |
| 103 | Ptenin-6-carboxylic acid | 53.22 | 1.473 ± 0.127 | 2.224 ± 0.183 | 0 | 0.024 ± 0.001 |
| 104 | 2,4,4-Trimethyl-3-hydroxymethyl-5a-(3-methyl-but-2-enyl)-cyclohexene | 53.48 | 0.218 ± 0.019 | 0.324 ± 0.027 | 0 | 0 |
| 105 | 2-(2-Dodecen-1yl)succinic acid | 53.76 | 0.801 ± 0.070 | 0.145 ± 0.012 | 0 | 0 |
| 106 | Ginsenol | 54.37 | 0.294 ± 0.026 | 0.394 ± 0.033 | 0 | 0 |
| 107 | Costunolide | 54.55 | 0 | 0.114 ± 0.009 | 0 | 0 |
| 108 | 1-Formyl-2,2,6-trimethyl-3-(3-methyl-but-2-enyl)-6-cyclohexene | 54.95 | 3.140 ± 0.273 | 1.125 ± 0.093 | 0 | 0 |
| 109 | 4-epi-Cubedol | 55.61 | 0.819 ± 0.071 | 0.313 ± 0.026 | 0 | 0 |
| 110 | 5-(6-Bromodecahydro-2-hydroxy-2,5,5a,8a-tetramethyl-1-naphthalene)-1,2-pentanediol | 55.82 | 0.441 ± 0.038 | 0 | 0 | 0 |
| 111 | (8S,14)-Cedran-diol | 56.00 | 0.855 ± 0.074 | 0.914 ± 0.075 | 0 | 0 |
| 112 | Cedrol | 56.39 | 0.379 ± 0.033 | 0 | 0 | 0 |
| 113 | 8-Propoxy cedrane | 56.65 | 0.187 ± 0.016 | 0.078 ± 0.006 | 0 | 0 |
| 114 | 5,6,6-Trimethyl-undeca-3,4-diene-2,10-dione | 56.81 | 0.311 ± 0.027 | 0.130 ± 0.011 | 0 | 0 |
| 115 | Octahydro [1,2]azaborino[1,2-a][1,2]azaborine | 57.53 | 0.535 ± 0.046 | 0 | 0 | 0 |
| 116 | 2.6-Dimethylnon-1-en-3-yn-5-yl valeric acid | 58.56 | 0.292 ± 0.025 | 0 | 0 | 0 |
| 117 | Terephthalic acid | 58.80 | 0.174 ± 0.015 | 7.487 ± 0.618 | 0 | 0 |
| 118 | Adamantane | 58.98 | 0.457 ± 0.040 | 0.944 ± 0.078 | 0 | 0 |
| 119 | 4,7-Methano-3,6,8-methenocyclopent[a]indene | 59.80 | 0.105 ± 0.009 | 0 | 0 | 0 |
| 120 | 4-Dimethylamino-2-methyl-1-phenyl-butan-2-ol | 60.11 | 0.316 ± 0.027 | 0.376 ± 0.031 | 0 | 0 |
| 121 | (±)cis-3,4-Dimethyl-2-phenyltetrahydro-1,4-thiazine | 61.09 | 0.185 ± 0.016 | 0 | 0 | 0 |
| 122 | 2-Isopropyl-6-phenylnicotinonitrile | 62.82 | 0.251 ± 0.022 | 0 | 0 | 0 |
| 123 | 2-Propenoic acid | 64.22 | 0.242 ± 0.021 | 0.300 ± 0.025 | 0 | 0 |
| 124 | Hexanedioic acid | 64.57 | 0.476 ± 0.041 | 7.976 ± 0.659 | 0 | 0 |
| 125 | Methadone N-oxide | 65.01 | 0.115 ± 0.010 | 0 | 0 | 0 |
| 126 | Isophthalic acid | 67.21 | 0.591 ± 0.051 | 1.164 ± 0.096 | 0 | 0 |
| 127 | Diisooctyl phthalate | 68.01 | 0.276 ± 0.024 | 0.420 ± 0.035 | 0 | 0 |
| 128 | Di-(2-methyoxyethyl) Isophthalate | 69.18 | 0 | 0.113 ± 0.009 | 0 | 0 |
| Total | 65.33 ± 5.564 | 84.057 ± 6.932 | 74.916 ± 6.073 | 95.661 ± 2.342 | ||
| Compound | Dry Weight (μg/g) |
|---|---|
| valerenic acid | 69.450 ± 0.263 |
| acetoxyvalerenic acid | 32.234 ± 0.961 |
| hydroxyvalerenic acid | n.d. 1 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.J.; Arasu, M.V.; Al-Dhabi, N.A.; Lim, S.S.; Kim, Y.B.; Lee, S.W.; Park, S.U. Expression of Terpenoid Biosynthetic Genes and Accumulation of Chemical Constituents in Valeriana fauriei. Molecules 2016, 21, 691. https://doi.org/10.3390/molecules21060691
Park YJ, Arasu MV, Al-Dhabi NA, Lim SS, Kim YB, Lee SW, Park SU. Expression of Terpenoid Biosynthetic Genes and Accumulation of Chemical Constituents in Valeriana fauriei. Molecules. 2016; 21(6):691. https://doi.org/10.3390/molecules21060691
Chicago/Turabian StylePark, Yun Ji, Mariadhas Valan Arasu, Naif Abdullah Al-Dhabi, Soon Sung Lim, Yeon Bok Kim, Sang Won Lee, and Sang Un Park. 2016. "Expression of Terpenoid Biosynthetic Genes and Accumulation of Chemical Constituents in Valeriana fauriei" Molecules 21, no. 6: 691. https://doi.org/10.3390/molecules21060691